











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introduction
Changes in the environment have spawned and emerging disease problems in forests worldwide (Ghelardini et al., 2016). Temperature and water stress spurred the activity of many endophytes, i.e., microorganisms that thrive asymptomatically on host plants (Terhonen et al., 2019a), that turn pathogenic as they struggle to adjust to new environments (Langer et al., 2021; Schulz et al., 2015). For example, the most damaging fungal species in Botryosphaeriaceae are found to be part of the endophytic community in woody plants that rapidly cause disease after an onset of host stress (Batista et al., 2021). Similarly, established fungal pathogens that are resistant to abiotic stressors become more active resulting in disease severity and spread. This was the case of warm temperature-adapted Heterobasidion sp. root pathogens that infected and caused mortality to new host trees under drought (Gomez‐Gallego et al., 2022; Terhonen al., 2019b). With climate change favoring many native and alien forest pathogens, disease forecasts generally point to a reduction of tree health as outbreaks become more severe and frequent (e.g., Contreras-Cornejo et al., 2023).
Philippine forests and managed landscapes have had a fair share of tree diseases and epiphytotics (Eusebio, 1998). The earliest yet sporadic published records of forest disease occurrences in the country could be traced back to 1890s during the onslaught of leaf rust (Hemileia vastatrix Berk. & Broome) on coffee that was introduced to the Philippines (Eusebio, 1998) followed by seedling damage due to damping-off in the 1920-30s (Kobayashi & de Guzman, 1988a b). Decades later, a survey from 1977 to 1985 by Kobayashi and de Guzman (1988a b) showed a diverse set of diseases occurring on natural forests, plantations, and forest nurseries in different regions of the country. Epiphytotics in Philippine forestry mostly occurred on monoculture plantations of non-native species which became increasingly extensive to meet increased demand for wood and fiber needed for construction and paper production (Philippine Council for Agriculture, Forestry and Natural Resources Research and Development [PCAARRD], 2015). For example, canker and gall rust (Uromycladium falcatariae Doungsa-ard, McTaggart & R. G. Shivas) diseases were two epidemic outbreaks caused by fungal pathogens that devastated large plantations and nurseries of Falcataria falcata (L.) Greuter & R. Rankin (Eusebio, 1998), a fast-growing non-native tree species commonly planted in Mindanao for pulp and paper production.
Fungal pathogens are equipped with a variety of evolutionary tools that enable them to transition from saprophytic to pathogenic and to overcome plant defenses (Masi et al., 2018), making these species highly efficient parasites. Fungal reproduction, both asexual and sexual, may further act to expand disease outbreaks by introducing and spreading novel hypervirulent strains (Ashu & Xu, 2015). Additionally, environmental modifications due to human activity only exacerbate pathogen spread and may accelerate evolution of virulence (Burgess et al., 2022). In the Philippines, Kobayashi and de Guzman (1988a b) noted 81 pathogenic fungi infecting about 65 hosts out of 85 taxonomically identified forest pathogens on 72 woody plants. Among these, 44 asexual morph species (i.e., species with unknown sexual state) of Ascomycota and Basidiomycota dominated the group. Although the list of tree pathogenic fungi in the country has expanded since then, many of these pathogens remain largely understudied.
As the Philippines experiences a much warmer climate with projections of seasonal heavy rainfall and drought (Philippine Atmospheric, Geophysical and Astronomical Services Administration [PAGASA], 2022), the impact on forest health under such conditions needs to be carefully examined. Furthermore, a better understanding of forest diseases in the Philippines will help assess the potential effects of environmental changes, particularly climate, on forest health in the country. This paper revisits the current state of knowledge on forest diseases in the Philippines with a specific focus on diseases caused by fungal pathogens to assess the vulnerability of Philippine forests to known foliar, stem and root diseases. Drawing from emerging forest disease trends worldwide, this paper also presents an assessment of forest health in the country and discusses its implications to forest management.
Materials and methods
This paper reviews the current knowledge of important forest diseases. Forest diseases in the Philippines were categorized based on the location of symptomatic tissue within the tree (foliage, stem, and roots. The study specifically focuses on diseases caused by fungal pathogens mostly occurring on economically important forest tree species. This review was initially conducted according to the Reporting Standards for Systematic Evidence Syntheses (ROSES; Haddaway et al., 2018). Data was obtained from two academic citation databases, Web of Science and Scopus. Search strings used were “tree AND disease AND Philippines”, “tree AND disease AND Philippines AND forest*” and “tree AND disease AND Philippines AND fung*.” No limit was set for the publication year. Only literature in the English language from all publication years recorded in the database was considered (n = 511). Search results not related to fungal diseases of forest plants in the Philippines were eliminated. Removing duplicates and title and abstract screening further narrowed down the search results to 14. Thus, literature search was also performed at local libraries and websites of the University of the Philippines and the Ecosystems Research and Development Bureau (2022). Other literature was further researched using Google Scholar. Fungal taxonomic names were verified using Mycobank, Index Fungorum, and the Global Biodiversity Information Facility databases.
Results and discussion
Foliar Diseases
Foliage diseases primarily affect the photosynthetic ability of plants by reducing chlorophyll production and damaging active leaf tissues (Debona et al., 2014). The majority of the foliar diseases are caused by fungi of the phyla Ascomycota and Basidiomycota that develop short multiple infections within a year or only a single year-long infection (Ata et al., 2024). Spores from either sexual or asexual structures are typically transmitted via wind and/or water splashes (Jain et al., 2019). Symptoms include leaf spots, anthracnose, blights, discoloration, and defoliation (Eusebio, 1998; Pacho & Quimio, 2015). Although foliar diseases tend to be of minor importance, mortality can occur on small, understoried and highly compromised stands. In a full-sib family trial of Pinus strobus L. with more than 4 000 trees, the prolonged severe Lophodermium needle cast was found to have strong negative impact on tree growth (r = −0.25 to−0.62; p < 0.05) and survival (r = −0.70, p < 0.001; Jansons et al., 2020). Foliar fungal diseases may be more damaging to conifers than hardwoods, which re-foliate after the shedding season (Nguyen et al., 2016). In a seasonally dry forest in Panama, synchronized leaf shedding and flushing were significantly negatively correlated (adjusted R2 = 0.6; p = 0.02) with the species richness of foliar pathogenic microfungi (Asterina, Phyllacora and Puccinia), suggesting a potential plant adaptation to reduce disease incidence (Piepenbring et al., 2015).
Not surprisingly, leaf and needle diseases are common in the Philippines due to the warm and humid climate. Approximately 72 fungal pathogens that cause foliar diseases have been recorded in the country (Figures 1 and 2, Table 1; Eusebio, 1998; Kobayashi & de Guzman, 1988a; Pacho & Quimio, 2015) which, except for occasional problems in forest nurseries and plantations, have only caused minor damage. Some causes may include leaf anatomy, age-associated resistance, and unfavorable environmental conditions (Lan et al., 2022; Smith et al., 2018; Wyka et al., 2017). Many of the most severe leaf diseases documented in the Philippines were also significant diseases in tropical and subtropical areas. The leaf rust outbreak in Philippine coffee plantations caused byH. vastatrix in the late 1800s led to a steep decline in coffee exports, from 16 million pounds to nearly zero (McCook, 2006). Today, coffee leaf rust continues to cause significant yield losses, amounting to up to US$2 billion annually, resulting in severe social and economic impacts in many coffee-producing regions (Talhinhas et al., 2017). Needle blight due to Mycosphaerella gibsonii H. C. Evans, a fungal pathogen that primarily affects pines in the tropics and subtropics, led to mortality in some nursery seedlings and natural regenerations including young stands of Pinus spp. in Luzon (i.e., Baguio, Mountain Province, Zambales, Laguna) and Mindanao (i.e., Agusan del Norte and Bukidnon) in the Philippines (Kobayashi et al., 1979). However, production losses have not been quantified. Rubber leaf fall disease caused by pestalotioid species is the most recent foliar disease outbreak on introduced Hevea sp. which plagued many growing regions in Southeast Asia, including more than 800-hectare plantations in Mindanao, the Philippines (Rodriguez, 2023). The disease can adversely affect latex production leading to subsequent economic losses (Alchemi & Jamin, 2022).
Table 1 Fungal pathogens known to occur in natural and planted forests and forest nurseries in the Philippines, excluding those associated with cultivated fruit trees and agroindustrial crops; including pathogens whose names have changed since they were originally reported in the country. Current species names cross-checked through Mycobank (M), Index Fungorum (I), and the Global Biodiversity Information Facility (G) databases.
| Species | Current name (Database/s) | Disease | Host | References |
|---|---|---|---|---|
|
Aecidium
mori (Barclay) Barclay |
Peridiospora mori (Barclay)
K.V. Prasad, B.R.D. Yadav & Sullia (M) |
Leaf rust | Morus alba | Kobayashi and De Guzman (1988a, 1988b) |
|
Botryodiplodia theobromae Pat. |
Lasiodiplodia
theobromae (Pat.) Griffon & Maubl. (M, I, G) |
Canker; Wildling blight on Dipterocarpus grandiflora, Swietenia macrophylla | Acacia mangium; Falcataria falcata | Kobayashi and De Guzman, (1988a, 1988b); Pacho and Quimio (2015) |
|
Camptomeris
albiziae (Petch) E.W. Mason |
Helminthosporium albiziae Petch (M, I, G) |
Yellow leaf | Falcataria falcata | Kobayashi and De Guzman (1988a, 1988b) |
|
Ceratocystis
ips (Rumbold) C. Moreau |
Ceratocystis
montium (Rumbold) J. Hunt (M) |
Wood stain | Pinus kesiya | Kobayashi and De Guzman (1988a, 1988b) |
|
Cercospora
artocarpi Syd. & P. Syd. |
Pseudocercospora artocarpi (Syd. & P. Syd.) Deighton (M, I, G) |
Leaf spot | Artocarpus altilis | Kobayashi and De Guzman (1988a, 1988b) |
|
Cercospora
bauhiniae Syd. & P. Syd. |
Pseudocercospora bauhiniae (Syd. & P. Syd.) Deighton (M, I, G) |
Leaf spot | Piliostigma malabaricum | Kobayashi and De Guzman (1988a, 1988b) |
|
Cercospora eucalypti Cooke & Massee |
Teratosphaeria eucalypti
(Cooke & Massee) Crous (M, I, G) |
Leaf spot | Eucalyptus deglupta | Kobayashi and De Guzman (1988a, 1988b) |
|
Cercospora
gardeniae Boedijn |
Pseudocercospora gardeniae (Boedijn) Deighton (M, I, G) |
Yellow leaf spot | Gardenia philastrei | Kobayashi and De Guzman (1988a, 1988b) |
|
Cercospora
gliricidiae Syd. & P. Syd. |
Passalora gliricidiae (Syd.
& P. Syd.) U. Braun & Crous (M, I, G) |
Leaf spot | Gliricidia sepium | Kobayashi and De Guzman (1988a, 1988b) |
|
Cercospora
gmelinae J. M. Yen & Gilles |
Pseudocercospora gmelinae
(J.M. Yen & Gilles) J.M. Yen (M); Pseudocercospora ranjita (S. Chowdhury) Deighton (I, G) |
Leaf spot | Gmelina arborea | Kobayashi and De Guzman (1988a, 1988b) |
| Cercospora henningsii Allesch. |
Clarohilum
henningsii (Allesch.) Videira & Crous (G) |
Leaf spot | Manihot carthaginensis | Kobayashi and De Guzman (1988a, 1988b) |
|
Cercospor a kurimensis Fukui |
Pseudocercospora
kurimensis (Fukui) U. Braun (M, I, G) |
Leaf spot | Nerium oleander | Kobayashi and De Guzman (1988a, 1988b) |
|
Cercospora
lythracearum Heald & F. A. Wolf |
Pseudocercospra lythracearum (Heald & F.A. Wolf) X.J. Liu & Y.L. Guo (M, I, G) |
Brown leaf spot | Lagerstroemia speciosa | Kobayashi and De Guzman (1988a, 1988b) |
| Cercospora paulowniae Hori |
Pseudocercospora paulowniae Goh & W.H. Hsieh (M); Pseudocercospora horii C. Nakash. & Tak. Kobay. (I, G) |
Leaf spot | Paulownia taiwaniana | Kobayashi and De Guzman (1988a, 1988b) |
|
Cercospora
philippinensis Tak. Kobay. & E. D. Guzman |
Pseudocercospora
philippinensis (Tak. Kobay. & E.D. Guzmán) U. Braun & Crous (M, I, G) |
Leaf spot | Mussaenda philippica | Kobayashi and De Guzman (1988a, 1988b) |
|
Cercospora
pini-densiflorae Hori & Nambu |
Pseudocercospora
pini-densiflorae (Hori & Nambu) Deighton (M); Mycosphaerella gibsonii H.C. Evans (I, G) |
Needle blight | Pinus caribaea, P. kesiya, P. merkusii, P. oocarpa | Kobayashi and De Guzman (1988a, 1988b) |
|
Cercospora
plumeriae Chupp |
Pseudocercospora plumeriae (Chupp) Tak. Kobay., Nishij. & C. Nakash. (M, I, G) |
Brown leaf spot | Plumeria alba | Kobayashi and De Guzman (1988a, 1988b) |
|
Cercospora
pterocarpicola J. M. Yen |
Pseudocercospra pterocarpicola (J. M. Yen) J. M. Yen (M, I, G) |
Brown leaf spot | Pterocarpus indicus | Kobayashi and De Guzman (1988a, 1988b) |
|
Cercospora ranjita S. Chowdhury |
Pseudocercospora ranjita (S. Chowdhury) Deighton (M, I, G) |
Leaf spot | G. arborea | Pacho and Quimio (2015) |
|
Cercospora
sequoiae Ellis & Everh. |
Passalora sequoiae (Ellis
& Everh.) Y.L. Guo & W. H. Hsieh (M, I, G) |
Needle blight | Taxodium mucronatum | Kobayashi and De Guzman (1988a, 1988b) |
|
Cercospora viticis Ellis & Everh. |
Pseudocercospora
viticicola (J. M. Yen & Lim) J. M. Yen (M); Pseudocercospora vitis (J. M. Yen & Lim) J. M. Yen (G) |
Leaf spot | Vitex parviflora | Kobayashi and De Guzman (1988a, 1988b) |
| Cercospora zizyphi Petch |
Pseudocercospora
ziziphi (Petch) Crous & U. Braun (M, I, G) |
Leaf spot | Ziziphus mauritiana | Kobayashi and De Guzman (1988a, 1988b) |
|
Coniothyrium
zuluense M. J. Wingf., Crous & T. A. Cout |
Teratosphaeria
zuluensis (M. J. Wingf., Crous & T. A. Cout.) M. J. Wingf. & Crous (M, I, G) |
Canker | Eucalyptus camaldulensis | PCAARRD (2003) |
|
Corticium
rolfsii (Sacc.) Curzi |
Agroathelia rolfsii (Sacc.)
Redhead & Mullineux (M, I); Athelia rolfsii (Sacc.) C.C. Tu & Kimbr. (G) |
Southern blight; Root rot | Swietenia macrophylla, Sindora supa | Kobayashi and De Guzman (1988a, 1988b); Pacho and Quimio (2015) |
|
Corticium
salmonicolor Berk. & Broome |
Necator
salmonicolor (Berk. & Broome) K. H. Larss., Redhead & T. W. May (M, I, G) |
Canker | Falcataria falcata | Kobayashi and De Guzman (1988a, 1988b) |
|
Crossopsora
antidesmae- dioicae (Racib.) Arthur & Cummins |
Crossopsora
antidesmatis- dioicae (Racib.) Arthur & Cummins (M); Cronartium antidesmatis- dioicae Syd. & P. Syd. (I) |
Leaf rust | Antidesma ghaesembilla | Kobayashi and De Guzman (1988a, 1988b) |
|
Cylindrocladium scoparium Morgan |
Calonectria
cylindrospora (Ellis & Everhart) Rossman, L. Lombard & Crous (M); Calonectria morganii Crous, Alfenas & M. J. Wingf. (I, G) |
Leaf spot | E. camaldulensis | Pacho and Quimio (2015) |
|
Ellisiopsis
gallesiae Bat. & Nascim. |
Beltraniella
portoricensis (F. Stevens) Piroz. & S. D. Patil (M, I, G) |
Leaf blotch | P. indicus | Kobayashi and De Guzman (1988a, 1988b) |
| Exosporium leucaena |
Camptomeris
leucaenae (F. Stevens & Dalbey) Syd. (M, I, G) |
Leucaena leucocephala | Kobayashi and De Guzman (1988a, 1988b) | |
|
Fomes rimosus (Berk.) Cooke |
Phellinus rimosus (Berk.)
Pilát (M, I, G) |
Wood decay | Dipterocarpaceae | Pacho and Quimio (2015) |
| Fomes pinicola (Sw.) Fr. |
Fomitopsis pinicola (Sw.)
P. Karst. (M, I, G) |
Butt rot and heart rot | Agathis sp. | Eusebio (1998); Pacho and Quimio (2015) |
|
Fusarium
solani (Mart.) Sacc. |
Neocosmospora solani (Mart.) L. Lombard & Crous (M) |
Damping off and root rot; top-killing (on L. leucocephala) | F. falcata, E. deglupta, L. leucocephala, P. caribaea, P. elliottii, P. kesiya, P. oocarpa, S. macrophylla | Kobayashi and De Guzman (1988a, 1988b) |
| Ganoderma sessile Murrill |
Ganoderma
resinaceum Boud. (I, G) |
Heart rot | Anisoptera thurifera | Pacho and Quimio (2015) |
|
Glomerella
cingulate (Stoneman) Spauld. & H. Schrenk |
Colletotrichum gloeosporiodes (Penz.) Penz. & Sacc. (M, I) |
Anthracnose (on leaves) | P. caribaea, Lansium domesticum, L. leucocephala, Mangifera indica, P. indicus | Kobayashi and De Guzman (1988a, 1988b) |
|
Macrophoma
micromegala (Berk. & M. A. Curtis) Berl. & Voglino |
Sphaeropsis micromegala
Berk. & M. A. Curtis (M) |
Needle blight | P. merkusii | Kobayashi and De Guzman (1988a, 1988b) |
|
Meliola
clerodendricola var. micromera (Syd. & P. Syd.) Hansf. |
Meliola micromera Syd.
& P. Syd. (M, I, G) |
Sooty mold | G. arborea | Kobayashi and De Guzman (1988a, 1988b) |
|
Olivea tectonae (T. S.
Ramakr. & K. Ramakr.) J. L. Mulder |
Neoolivea tectonae (Racib.) Aime & McTaggart (M, I, G) |
Teak rust (on leaves) | Tectona grandis | Kobayashi and De Guzman (1988a, 1988b) |
|
Phellinus
noxius (Corner) G. Cunn. |
Pyrrhoderma
noxium (Corner) L.W. Zhou & Y.C. Dai (M, I, G) |
Root rot | Fabaceae, Aleurites moluccanus, A. occidentale, Casuarina equisetifolia, Dalbergia sissoo, Dracontomelon dao, Azadirachta indica, M. alba, Maranthes corymbosa | Militante and Manalo (1999); PCARRD (2006) |
|
Phaeoisariopsis anthocephala Ts. Kobay. |
Pseudocercospora anthocephali (Ts. Kobay.) Deighton (M, I, G) |
Brown leaf spot | B. chinensis | Kobayashi and De Guzman (1988a, 1988b) |
|
Phaeoseptoria eucalypti Hansf. |
Teratosphaeria epicoccoides (Cooke & Massee) Rossman & W. C. Allen (M) |
Black powdery spot (on leaves) | Eucalyptus sp. | Kobayashi and De Guzman (1988a, 1988b) |
|
Phakopsora
gossypii (Arthur) Dale |
Kuehneola gossypii Arthur
(M); Phakopsora desmium (Berk. & Broome) Cummins (I, G) |
Rust (on leaves) | Gossypium sp. | Kobayashi and De Guzman (1988a, 1988b) |
| Phyllachora parkiae Henn. |
Phyllacora acaciae subsp.
parkiae (Henn.) P. F. Cannon (M, I, G) |
Tar spot (on leaves) | Parkia timoriana | Kobayashi and De Guzman (1988a, 1988b) |
|
Phyllachora
spinifera (P. Karst. & Har.) Höhn. ex Rehm |
Phyllacora ficuum var.
spinifera P. Karst. & Har. (M); Phyllacora aspidea (Berk.) Sacc. (I, G) |
Tar spot (on leaves) | Ficus odorata | Kobayashi and De Guzman (1988a, 1988b) |
| Puccinia psidii G. Winter |
Austropuccinia psidii (G.
Winter) Beenken (M, I, G) |
Leaf rust | Myrtaceae | PCARRD (2006) |
|
Ravenelia
berkeleyi Mundk. & Thirum. |
Ravenelia
cassiicola G. F. Atk. (I, G) |
Leaf rust | Senna multijuga | Kobayashi and De Guzman (1988a, 1988b) |
|
Uromycladium
falcatarium Doungsa-ard, McTaggart & R. G. Shivas |
Uromycladium
falcatariae Doungsa-ard, McTaggart & R. G. Shivas (M, I) |
Gall rust (on stem) | F. falcata | Doungsa-ard et al. (2015) |
Foliage diseases are projected to become more prevalent as sporulation and infection of many foliar fungal pathogens intensify with frequent warm rain events (Ata et al., 2024). As in many regions worldwide, more intermittent outbreaks of foliar diseases could likely occur in the Philippines during warm and wet seasons, although seasonal drought may slow down foliar disease impact (Hoheneder et al., 2021). Nevertheless, Philippine forests remain likely candidates to host foliar diseases that are currently emerging in tropical and subtropical regions. For example, the climate space of the country is predicted to be suitable for the pandemic biotype of myrtle rust Austropuccinia psidii (G. Winter) Beenken, an emerging invasive pathogen in the North America, South America, Asia and the Pacific, and Africa that affects plants in the family Myrtaceae (Stewart et al., 2018). Introduction of this biotype (e.g., international plant trade, poor implementation of phytosanitary regulations, etc.) poses a huge risk to the Philippines that is rich in highly susceptible Myrtaceae species.
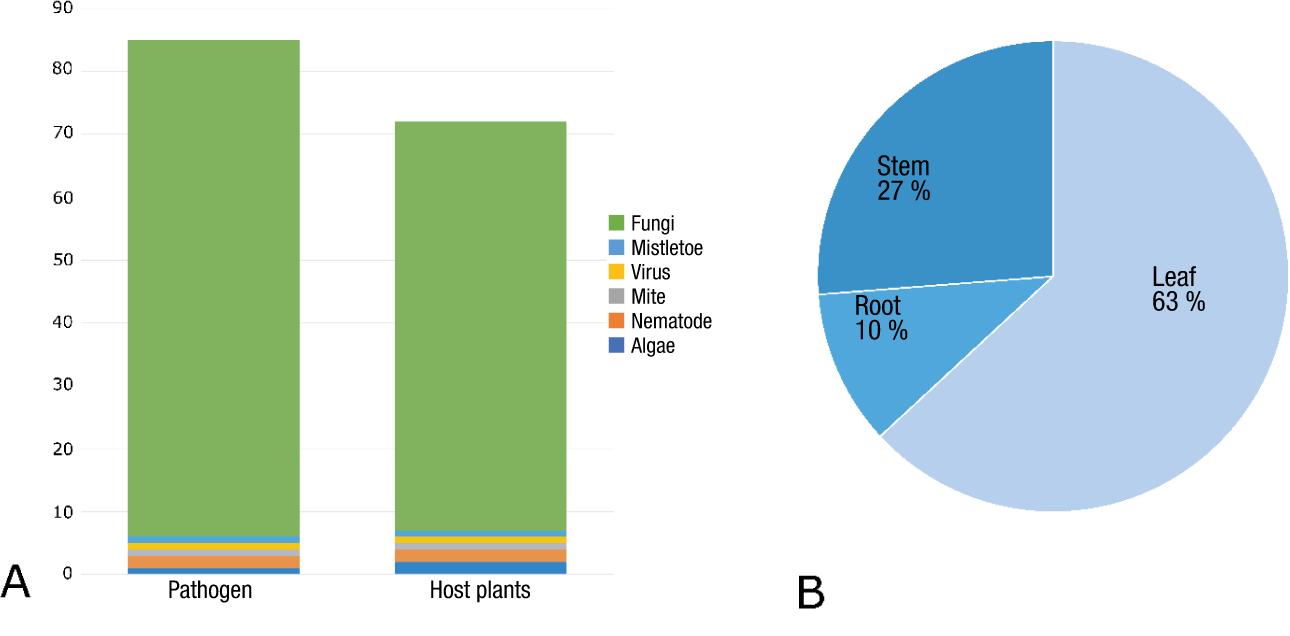
Figure 1 (A) Number of forest pathogens and woody host plants according to Kobayashi and de Guzman (1988a b), and (B) proportion of forest diseases on leaves, stems, and roots (Eusebio, 1998; Kobayashi & de Guzman, 1988a; Pacho & Quimio, 2015) in the Philippines.
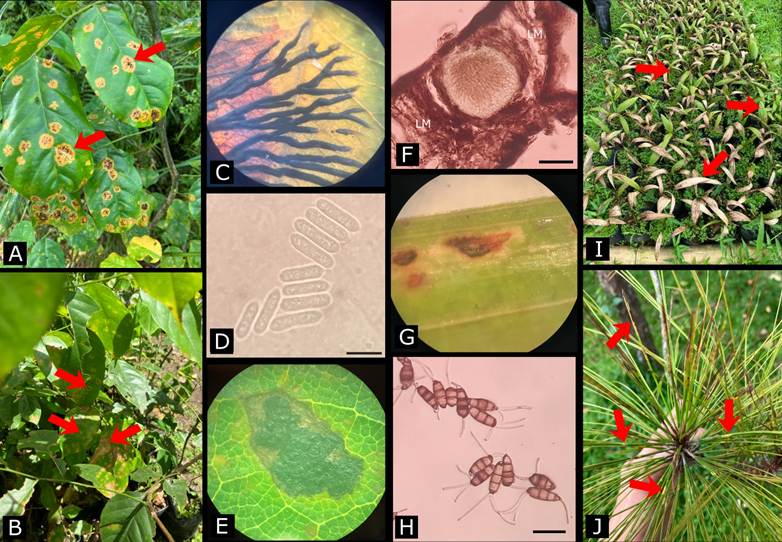
Figure 2 Example of foliar diseases of tree species observed in forests, plantations, urban environments and forest nurseries in the Philippines. Leaf spot on Pterocarpus indicus Willd. caused by Aldona stella-nigra Racib. (A) with linear, branching ascomata (C) on leaf surface. Leaf necrosis on P. indicus (B) associated with Colletotrichum sp. which produces spores (D) on agar. Tar spot on Intsia bijuga (Colebr.) Kuntze caused by Phyllacora sp. which produces dark shiny stromata (E) and ascomata (F) embedded on leaf mesophyll (LM). Leaf spot on Areca (G, I) caused by Pestalotiopsis sp. with conidia (H) produced on agar. Yellowish to reddish bands on needles of Pinus kesiya Royle ex Gordon (J) caused by unidentified agent. Red arrows show examples of symptoms on plant tissue. Scale bars for D, F and H measure 10 µm, 0.25 mm and 15 µm, respectively. Photo credits: Colletotrichum sp. spores (D) by C. A. Ramos, Phyllacora sp. on leaf tissue (F) by J. K. Saddoy, and Pestalotiopsis sp. conidia (H) by R. A. Mastrili.
Stem diseases
Many stem diseases are economically important due to their direct impact on wood and its derivatives (Yurkewich et al., 2017). Stem diseases typically result in canker or necrosis on the xylem tissue, wilting, heart rot or gall on the main stem and on branches (Harsh, 2025). These occur when spores land on wounds of branches or stems and initiate infections on stressed hosts (Ma et al., 2015). However, recent studies have shown that some fungal taxa from the family Botryosphaeriaceae can infect directly through natural openings of healthy plants (Slippers & Wingfield, 2007). Shi et al. (2024) found that the spatial pattern of host trees, in addition to host abundance and species composition, can influence the infection of fungal stem diseases. This finding has implications in the establishment of monoculture plantations and may also apply to tree diseases beyond those affecting stems. Economic losses resulting from reduced timber volume and wood quality due to stem diseases largely threaten the sustainability of pulp and solid-wood industries. For example, damage from canker disease (Caliciopsis pineaPeck) on P. strobus stands led to an average loss of 7.27 USD·m-3 in lumber value, and with even lower revenue from severely infected trees (Costanza et al., 2019). Tree mortality and forest decline are also warranted on severely cankered stands especially in anthropogenically disturbed sites (Paap et al., 2017).
The Philippine timber industry incurred production losses from stem diseases in tree plantations and natural forests (Pacho & Quimio, 2015). Recently, heart rot affected 20-38 % of planted overgrown exotic Acacia mangium Willd. (Rojo & Paquit, 2018), a tree species introduced to the Philippines. Causal agents are possibly fungal members of the phylum Basidiomycota including Fomes, Phellinus and Ganoderma (Figure 1, Table 1; Eusebio, 1998; Pacho & Quimio, 2015). Meanwhile, important canker and dieback diseases in the tropics caused by Necator salmonicolor (Berk. & Broome) K. H. Larss. and Diaporthe eres Nitschke, respectively, had also severely affected Philippine plantations of introduced F. falcata and Acacia auriculiformis A. Cunn ex Benth. in Surigao del Sur, Mindanao and in Nueva Ecija, Luzon, respectively. These plantations were established in likely compromised sites during the 1980s (Kobayashi & de Guzman, 1988a).
While incidence of some stem diseases seems to have dropped, others have persisted, and a few have newly emerged. Gall rust disease caused by U. falcatariae (Figure 3), a pathogen specific to F. falcata (Doungsa-ard et al., 2015), remains to exist in the Philippines since its first report in 1989 (Eusebio, 1998). On the other hand, a new canker disease caused by Teratosphaeria zuluensis (M. J. Wingf., Crous & T. A. Cout.) M. J. Wingf. & Crous was first spotted in the country on Eucalyptus camaldulensis Dehnh. plantations in 2002 (PCAARRD, 2003). Known to occur on nine Eucalyptus species including their hybrids, this pathogen is distributed across many countries and has continuously spread due to international trade (Aylward et al., 2019). In most cases, however, information on the economic losses caused by stem diseases in the Philippines remains scant.

Figure 3 Example of stem and root diseases of tree species observed in forests, plantations, urban environments and forest nurseries in the Philippines. Falcataria falcata (L.) Greuter & R. Rankin showing hypertrophic symptoms such as galls on branches and distorted shoots (red arrows) on mature trees (A) and seedlings (B) due to de presence of Uromycladium falcatariae Doungsa-ard, McTaggart & R.G. Shivas which produces teliospores (C) on symptomatic tissue. Characteristic crown thinning and defoliation on Fabaceae species caused by Ganoderma spp. which typically produce sporocarps basal part of the stem (D, E). Photo of U. falcatariae (C) courtesy of J. K. Soriano.
Climate change exacerbates stress on host trees which, in turn, increases host susceptibility to stem pathogens (Teshome et al., 2020). As successful infection and disease expression require hosts stressed by environmental factors, diseases and insects, stem diseases such as canker and decay are predicted to cause severe impact especially during warmer and drier seasons (Wang et al., 2023). Dry and warm weather also tends to favor insect vectors (Panzavolta et al., 2017) and, thus, forecasts of stem disease outbreaks caused by insect-vectored pathogens come as no surprise. Interestingly, in addition to climate change-related disturbances (e.g.Choi et al., 2022; Davydenko et al., 2021), human activities have been implicated in the increasing trend of invasive tree diseases caused by ophiostomatoid fungi associated with tree-colonizing insects (Wingfield et al., 2017). In the Philippines, Pinus kesiya Royle ex Gordon wood (Kobayashi & de Guzman, 1988a) has been reported infested by the ophiostomatoid fungus Ceratocystis, associated with Ips calligraphus (Germar, 1824). However, its involvement in the recent increase in tree mortality remains unclear.
Although the focus of this review is on tree diseases caused by fungal pathogens, it is important to note that other insect-vectored stem diseases such as pine wilt have recently become a serious global threat (Back et al., 2024). Pine wilt disease (PWD) is caused by an endoparasitic nematode Bursaphelenchus xylophilus (Steiner & Buhrer, 1934) Nickle, 1970 vectored by a pine sawyer beetle Monochamus and has reportedly invaded forests in Europe and Asia (Espada et al., 2022). Nematode infestation in tree vascular systems at favorable climate results to blockage of sap ascent which induces tracheid cavitation leading to mortality (Yazaki et al., 2018). No reports of entry have been recorded in the country despite its occurrence in neighboring Asian countries (Back et al., 2024). However, the invasion of PWD in forests in northern Philippines is highly likely to occur due to suitable climate and presence of susceptible Pinus species (Hirata et al., 2017).
Root diseases
Despite ecological and economic importance, root diseases often remain undetected and consequently understudied due to their inconspicuous occurrence and slow symptom development (Allikmäe et al., 2017). In mature stands, root-infecting fungal pathogens mainly under Basidiomycota phylum affect tree structure by attacking root cambium and xylem cells (e. g., Kedves et al., 2021). Some root pathogens spread through spores or rhizomorphs while others take advantage of root-to-root connections or root grafts (Devkota & Hammerschmidt, 2020; Zaļuma et al., 2019). Root pathogens can also spread through site preparation and thinning practices, where machinery both disperses inoculum and causes root injuries (Laflamme, 2008). Infected trees may initially appear to be nutrient-deficient showing discoloration to wilted leaves and growth decline attributed to altered physiological responses like stomatal conductance and chlorophyll fluorescence (Yazaki et al., 2018). Highly vulnerable to native root pathogens, exotic forest plantations can expect major losses from root diseases in sites with poor land conditions or a history of root disease (Eyles et al., 2008). For example, without proper site management, Malaysian rubber tree (Hevea brasiliensis [Willd. Ex A. Juss.] Müll. Arg.) plantations could face a 500 000 ha loss from white root rot caused by Rigidoporus microporus (Sw.) Overeem and consequently affect domestic latex production and supply chain (Olaniyi & Szulczyk, 2022).
While many of the root diseases recorded in the Philippines occur among forest nurseries (Figure 1; Table 1), a few have caused major losses in forest plantations. First reported in 1991, brown root rot caused byPyrrhoderma noxium(Corner) L. W. Zhou & Y. C. Dai resulted in up to 25 % mortality in 4- to 10-year-oldA. mangiumplantations, particularly in Mindoro and Mindanao, where most trees in 1-4-hectare plantations were affected (Militante & Manalo, 1999). Interestingly, despite its unknown origin and complex migration histories (Kozhar et al., 2022), the pathogen remains categorized as invasive in the Philippines owing to its wide host range (11 native Philippine tree species; PCAARRD, 2006). However, the current incidence ofP. noxiumin the country has reportedly declined, despite the relatively large number ofA. mangiumhosts still present in Mindanao (Forest Management Bureau [FMB], 2022). On the other hand, Ganoderma spp. continues to exist, albeit at a slow rate, in pockets within forest stands and plantations in the Philippines (Figure 3). Interestingly, despite a broad host range, current incidence reports of P. noxium and Ganoderma lucidum (Fr.) P. Karst. in natural forests seem to be minor (Pinol et al., 2006).
Forests experiencing drier and warmer climate are predicted to be the most vulnerable to root diseases (Kim et al., 2021). This may be attributed to multiple factors such as higher production of pathogen rhizomorph, decline of symbiotic microbial communities (Kubiak et al., 2017), and reduced endogenous defense mechanisms of predisposed hosts (Aslam & Magel, 2018). As the country's temperature rises, with maximum levels expected to exceed 35 °C (PAGASA, 2022), the future climate conditions are likely to remain favorable for the growth ofP. noxium, which can tolerate temperatures above 36 °C (Stewart et al., 2020). The same climate space however may be less suitable for other root pathogens like Ganoderma boninense Pat. (Paterson, 2020), a basal stem rot pathogen that aggressively attacks many Southeast Asian oil palm (Elaeis guineensis Jacq.) plantations but is currently of low incidence in the Philippines.
Implications for forest management
Global forest disease outbreaks and decline syndromes unravel the devastating effects of a changing climate (Ramsfield et al., 2016). As the climate worsens, predicted scenarios describe the likely winners and losers in forest disease interactions and a rather uncertain state of the forest health landscape (Desprez-Loustau et al., 2018). This uncertainty can be attributed to the complex nature and cryptic geographic distribution of fungal tree pathogens (Desprez-Loustau et al., 2018), many of which remain understudied if not unknown. Also, in many forest pathosystems, host-pathogen co-existence under varying climatic and environmental changes induced by human activities is poorly understood and thus further confounds disease projections (Heinzelmann et al., 2019). All these challenges are further amplified in countries like the Philippines where resources for forest health protection-related activities have been limited.
Despite projections of increased vulnerability, there is little current evidence linking declining forest health in the Philippines to climate-driven forest diseases, due to a lack of recent reports on forest pathogens in the literature. However, as Eusebio (1998) noted, marginal to no disease records in Philippine forests do not necessarily imply absence of infection nor perpetual immunity. The environmental situation in the Philippines, including the worsening climate (PAGASA, 2022) and still diverse yet declining ecosystem (Coritico et al., 2020; Parlucha et al., 2021), may have helped limit the spread of forest diseases. Alternatively, gaps in monitoring and research may have hindered an up-to-date record of these diseases. These significant information gaps are common in developing tropical countries, where the focus is often on more pressing issues. Greater attention is typically given to forest pressures such as deforestation from resettlement, large-scale agriculture, and shifting cultivation, which are seen as more critical than climate change (Zhao et al., 2005).
Uncertainties in forest disease projections present opportunities to establish and implement a more proactive forest management system in the country equipped with diverse and innovative practices (Gullino et al., 2022). Specifically, as the sensitivity of many forest fungal pathosystems to changing climate are high yet variable, forest managers must employ a more flexible forest management approach “that promotes reversible and incremental steps, and that favors ongoing learning and capacity to modify direction as situations change” (Millar et al., 2007). This entails a forest health management system that is fully integrated into four key aspects of forest management: monitoring, forecasting, planning and mitigation (Sturrock et al., 2011).
Monitoring
Regular and systematic tree health monitoring in permanent forest plots that incorporate other long-term ecological data can effectively track extensive, sudden and at times subtle changes in stand health (Weldon & Grandin, 2019). This can be integrated into the national forest resources assessment program which currently focuses on tree biophysical characteristics and stocks and flow of wood and non-wood forest products and services. Monitoring data would be stored in a national online database, providing updated records of existing and emerging tree diseases, along with distribution maps (e.g., Zhang et al., 2024). This allows users to scale data from provincial to regional levels, supporting forest health research and policy needs. Furthermore, this information could be integrated into species suitability assessments that consider regional stress conditions to better inform forest planning, including reforestation and plantation establishment programs.
However, in the Philippines, this may represent a challenge, as the identification of many fungal pathogens, especially those affecting trees, largely relies on morphological descriptions and outdated taxonomic keys (Crous et al., 2016). The diversity of phenotypic traits within a single species can be confusing, as the plasticity of the few distinguishing characteristics under varying environmental conditions may result in multiple names being assigned to the same species (Ata et al., 2021; Boddy, 2016). Conversely, unrelated fungal species may develop similar morphologies through evolutionary convergence, further contributing to inaccurate characterization (Boddy, 2016). This challenge is compounded by the lack of trained personnel for the assessment and monitoring of forest diseases, as well as by the absence of specialized facilities, such as forest diagnostic clinics, to support the identification of tree diseases.
The Philippines may also capitalize on the surge of new diagnostic tools for efficient disease diagnosis and phytosanitary measures. DNA-based tools, which may be inaccessible for many scientists in developing nations like the Philippines, are already available for fast, reliable and accurate identification and detection of several cultivable and non-cultivable fungal phytopathogens on either symptomatic or asymptomatic plant tissues (Ata et al., 2022; Hariharan & Prasannath, 2021). While not without its challenges, the advancements in molecular diagnostics for identifying fungal tree pathogens are promising for early disease surveillance and plant biosecurity (Luchi et al., 2020). These tools, including polymerase chain reaction assays, isothermal amplification methods, and next-generation sequencing, use DNA and/or RNA to detect target pathogens without the need for culturing (Hariharan & Prasannath, 2021). They offer high sensitivity, enabling the detection of a single target molecule in complex mixtures, while being rapid and versatile (Hariharan & Prasannath, 2021). In the Philippines, some of these techniques have been gradually integrated in agriculture (De la Cueva et al., 2021; Hussien et al., 2017) but have yet to be adopted in forestry.
A complementary, low-cost strategy to the more expensive molecular diagnostics is citizen science, which involves communities in data collection, project design, analysis, and dissemination. This approach has demonstrated potential for monitoring forest pests and engaging the public in forest health efforts across various regions, including Europe (Baker et al., 2019; de Groot et al., 2023), North America (Lanning et al., 2023), and in ecological research in Africa (Hulbert, 2016). While data from citizen science projects may be less accurate and biased (Baker et al., 2019), proper management can improve overall sampling efficiency. In regions like the Philippines, where support for forest health research and monitoring may be insufficient, the continuous emergence of pests and diseases highlights the value of centrally coordinated citizen science projects. These initiatives can significantly expand the network of observers and enhance monitoring efforts (Hurley et al., 2017).
In monitoring efforts within natural forests, particular attention should also be given to signs and symptoms of diseases caused by emerging and/or known invasive pathogens not yet reported locally but predicted to establish under Philippine climatic conditions, particularly those threatening hosts with small or fragmented populations. This is especially relevant in the Philippines, where resources for forest health management are limited. Focusing on high-risk species allows for early detection using field-based visual assessments, which require minimal equipment and training compared to molecular diagnostics or broad-scale surveys. For example, myrtle rust (A. psidii) poses a potential risk to Eucalyptus deglupta Blume in Mindanao, an endemic species classified as vulnerable by the International Union for Conservation of Nature (IUCN, 2024) due to its declining and fragmented populations. Similarly, symptoms of PWD, although caused by a nematode rather than a fungal pathogen, warrant surveillance in native Pinus merkusii Jung. & de Vriese, which is also listed as vulnerable by the IUCN (2024).
Forecasting
Bioclimatic envelope models, although not without its limitations, are important tools that inform forestry professionals on the potential magnitude and future shifts in the forest disease landscape (Sturrock et al., 2011; Woods et al., 2010). These dynamic simulation models integrate various ecological and climatic data to predict suitable spaces for hosts and pathogens across future conditions. Modeling the influence of climate to forest pathosystems, particularly those involving high-risk invasive pathogens, can help identify priority areas in monitoring programs especially in countries with limited financial resources like the Philippines. An initial application of such modeling in the Philippines includes the bioclimatic analysis of myxomycetes (Limbo-Dizon et al., 2022), demonstrating the feasibility of these tools for forest management. But to apply this in emerging forest diseases, there is a need for research on ecological niches of fungal pathogens in the country, which is currently lacking and/or fragmented. Notably, the assessment of the potential susceptibility of E. deglupta to A. psidii identified critical areas in Mindanao that warrant close monitoring for possible myrtle rust outbreaks (Villasan, 2025). This study, which leveraged environmental data from external sources, underscores the feasibility of bioclimatic modeling despite data limitations, thus highlighting the importance of collaborative research and data-sharing initiatives.
Furthermore, the diverse and often competing interests of stakeholders (forest communities, national and local government, non-government organizations, timber industries, etc.) in Philippine forestry, most affected by current and future forest disease outbreaks, can guide model development, simulations, and counterfactual analysis (Gaydos et al., 2019). Therefore, integrating participatory modelling in developing epidemiological models for plant disease control may be explored (Gaydos et al., 2019). This approach not only leverages local knowledge but also helps bridge knowledge-practice gap through collaborative learning and stakeholder empowerment.
Planning
Forest management plans can benefit from risk assessments and plant health legislations with the aim of a strategic immediate response to reduce possible undesirable impacts of climate change on forest pathogens (Sturrock et al., 2011). While relatively unexplored for forest pathogens in the Philippines, pest risk analyses and risk-rating systems accounting for climate change can help assess potential impacts of forest pathogens and guide regulatory decisions related to forest health. Along with monitoring data, research data on fungal pathogens as well as environmental drivers of host tree-pathogen interactions in forest ecosystems can further strengthen risk factor models to best understand climate-disease relationships (Hennon et al., 2021). Apart from ecological and biophysical aspects, factors related to pathogen movement through forest trade are equally important in identifying critical control points for effectively mitigating the entry of emerging forest pathogens (Ristaino et al., 2021). This requires a strong collaboration between academic and national research laboratories and partnerships with forest industries, which should be fostered and maintained to develop a solid forest pathology research program in the Philippines. Scientists must pursue global research collaborations to enhance efforts and address topics more advanced in other regions, as demonstrated in South America's forest health initiatives (Villacide et al., 2023). Funding for such collaborations typically involves partnerships between government and the commercial sector (Wingfield et al., 2015). However, greater coordination is needed to promote investments in long-term, sustainable landscape-scale initiatives (Clark et al., 2018).
Mitigation
Tree diversity, particularly of non-host species, is widely recognized to greatly improve forest resilience against pathogens (Roberts et al., 2020; Saadani et al., 2021). For example, although forest communities consequently harbored diverse foliar fungal pathogens, those with diverse non-host trees significantly reduced foliar disease severity per tree species (Rutten et al., 2021). Thus, Philippine forest biodiversity conservation programs and silvicultural interventions (e.g., field trials for tree improvement programs; Figure 4) aimed at maintaining or enhancing natural tree diversity at both genetic and species levels are increasingly critical for promoting forest health under uncertain future conditions. These efforts contribute to the development of near-natural forests, which are more resilient to environmental changes and threats. In production forests, simply increasing diversity may not be sufficient to incentivize timber industries with private concessions to adopt this approach, as diversification strategies often yield lower economic returns (Hildebrandt & Knoke, 2011). Therefore, it is essential to use forest management practices to achieve resilient forests while maintaining stable long-term revenues. These practices may include targeted species harvesting, varied harvest systems, a combination of natural regeneration and planting, and the use of a broader range of planted species (Dymond et al., 2014).

Figure 4 Progeny trials of Falcataria falcata (L.) Greuter & R. Rankin (A) and Swietenia macrophylla King (B) in Mindanao established from candidate mother trees representing diverse provenances from the regions of the Philippines
For large-scale tree plantations, economic and ecological trade-offs associated with mixed cropping of non-host trees must be carefully analyzed considering the cost and long-term benefits of such practice to manage emerging diseases (Roberts et al., 2020). Planted forests of economically important trees can also greatly benefit from resistance breeding programs and use of genetically diverse and well-adapted seedlots. Due to the slow nature of breeding programs, investigating general resistances or tolerances aside from species-specific resistance may be needed to respond to a rapidly changing climate (Woods et al., 2010).
Conclusions
Philippine forests face significant threats from potential emerging fungal diseases largely driven by climate change, which may lead to more frequent and severe tree disease outbreaks. Despite past forest epidemics, the country’s forest management has shown limited capacity to anticipate and effectively respond to these emerging risks. Therefore, the implementation of a more proactive and integrated forest health strategy is urgently needed. Given the limited financial and technical resources for forest health monitoring in the country, low-cost strategies like citizen science and targeted monitoring offer practical solutions for early detection and community engagement. These strategies safeguard vulnerable forests cost-effectively. It is essential to close information gaps through solid research and development in key areas such as forest pathology, ecology, and silviculture. Broad and coordinated participation among stakeholders, including government, the private sector, local communities, and international partners, will be crucial to ensuring sustainable and effective forest health management in the country.

