











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introduction
Pinus pinceana Gordon, known as weeping pinyon, is an endemic species of Mexico that is endangered and distributed in the semi-arid regions of the Sierra Madre Oriental (Favela & Thomas, 2013; Secretaría de Medio Ambiente y Recursos Naturales [SEMARNAT], 2023). In this upland area, pinyon pine forests are key components of the ecosystem; however, weeping pinyon remains one of the least studied species (Favela et al., 2009; Sánchez-Castillo et al., 2022; Santillán-Hernández et al., 2010).
Conifers like P. pinceana are naturally distributed in geographic areas with pronounced climate seasonality, where trees halt their growth during unfavorable periods and resume it when climatic conditions become optimal. The evidence of this activity and dormancy is recorded in the wood structure as annual concentric layers that represent radial growth, which, in a cross-section, appear as a pattern of tree rings (Gutiérrez, 2009).
The tree rings in conifers consist of two bands of coloration: a light-colored band representing earlywood developed during spring, and a dark-colored band indicating the formation of latewood during summer. The difference in coloration between the last and the first cells of the year defines an annual tree ring (Speer, 2010).
In Mexico, precipitation is one of the most significant climatic factors limiting the radial growth of trees, especially in temperate and semi-arid climates. This information has enabled the reconstruction of precipitation data in areas where most weather stations have less than 50 years of records (Arroyo-Morales et al., 2023; Carlón et al., 2018; Cortés-Cortés et al., 2021; Villanueva-Díaz et al., 2020). Nevertheless, it is important to increase dendrochronological studies at sites where long-range climatic information is not available. However, it is not only sufficient for tree species to produce annual tree rings, but also to show that they have dendrochronological potential.
A tree species is considered to have dendrochronological potential when it has the capacity to develop annual tree rings with a strong climatic signal, reflected in the variations in tree rings width (Speer, 2010). To determine the dendrochronological potential, some basic statistical parameters are considered, such as intercorrelation between series, mean sensitivity, standard deviation, autocorrelation and signal-to-noise ratio. All of them are focused on determining whether there is fluctuation in the annual tree rings width and the extent of that fluctuation (Grissino-Mayer, 2001).
The dendrochronological potential of P. pinceana has only been addressed by Santillán-Hernández et al. (2010), who studied the species' response to precipitation in populations from Coahuila, Hidalgo, San Luis Potosí, Querétaro, and Zacatecas, which were its known distribution areas at the time (including Nuevo León). Subsequently, P. pinceana was recorded in the southwestern part of Tamaulipas in 2015 (Martínez-Ávalos et al., 2015); therefore, it is now possible to expand the understanding of its dendrochronological potential to include its far northeastern distribution.
The northeast part of Mexico is characterized by low levels of precipitation and frequent droughts (Stahle et al., 2016), so the study of long-term precipitation patterns is essential to develop strategies to prevent the impacts of droughts. A long-range source of climate information, due to its longevity and habitat in semi-arid areas, is P. pinceana. The evaluation of the dendrochronological potential of this species would allow us to know the climatic factor that limits its radial growth in Tamaulipas, and then explore the use of the tree-rings of P. pinceana in climatic reconstructions of the region.
The hypothesis of this study was that due to the arid conditions where P. pinceana establishes itself in Tamaulipas, it is expected to have good dendrochronological potential, with precipitation being the climatic factor that limits its radial growth. To test the assumption of this research, the objectives were: 1) to develop an annual tree-ring width chronology of P. pinceana; 2) to determine its dendrochronological potential to register, in their tree rings, the climatic variability in the study area and 3) to associate radial growth with precipitation and regional temperature to support the climatic sensitivity of the species.
Materials and methods
Description of the study area
Specimens of P. pinceana are located in the municipality of Jaumave, near the ejido of Magdaleno Aguilar, and are part of the submontane scrub vegetation in the Sierra Madre Oriental in the southwestern part of Tamaulipas (SEMARNAT, 2014). The area where P. pinceana is found is a natural landscape not designated for timber extraction. The reference coordinates for the ejido are 23° 27’ 08.9565’’ N and -99° 33’ 32.945’’ W, where P. pinceana trees grow at an average elevation of 1 200 m (Figure 1).
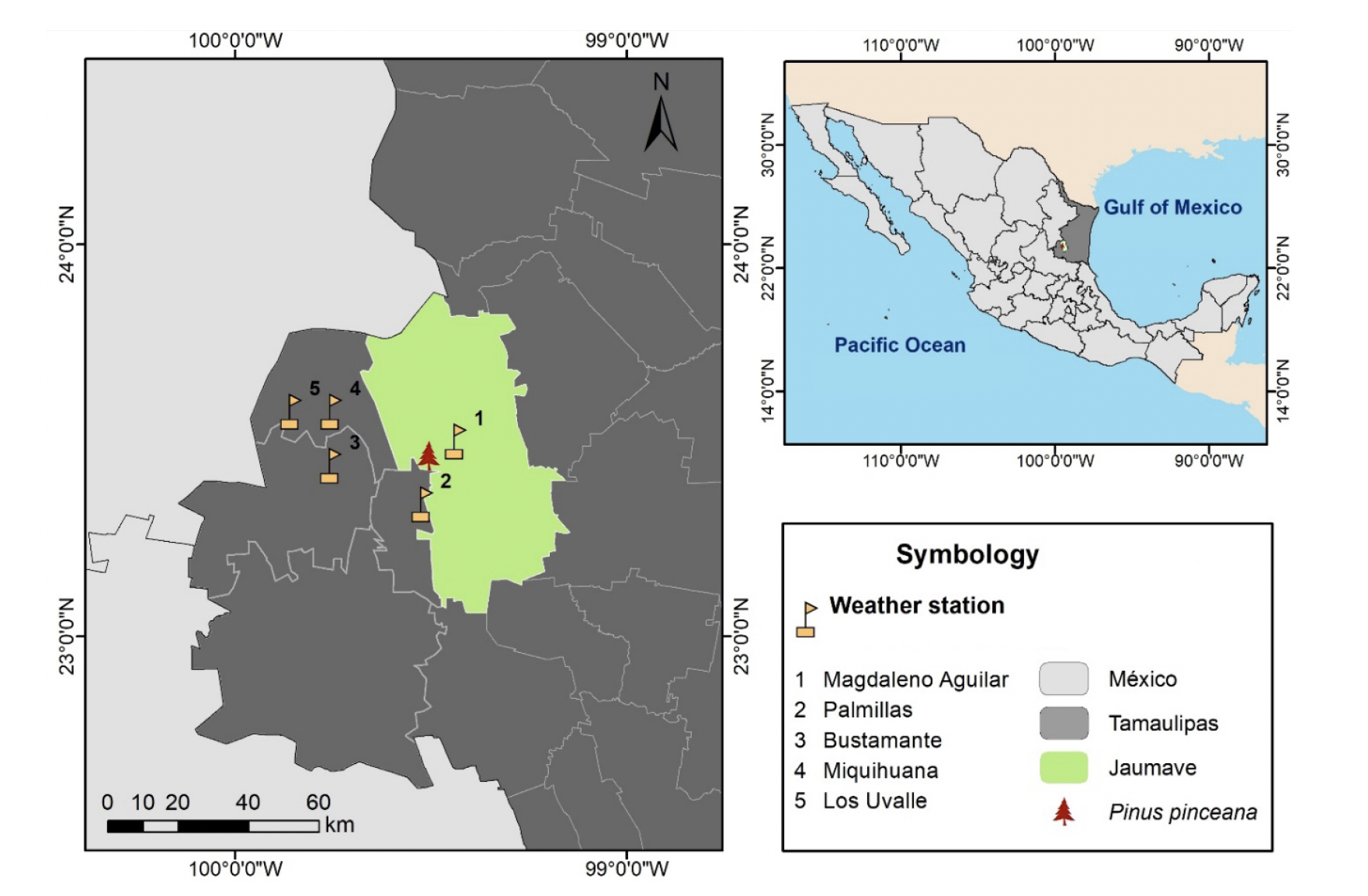
Figure 1 Geographic location of the Pinus pinceana population in Tamaulipas and the weather stations used in this study.
The climate of the study area is semi-arid with extreme temperatures (García, 2004). It has an average annual precipitation of 491.56 mm, with September being the wettest month at 80.11 mm and February the driest at 10.09 mm. The average annual precipitation for spring, summer, autumn, and winter is 24, 42, 26, and 8 %, respectively, with cold fronts playing an important role in low-intensity precipitation during winter (Magaña et al., 2003). The average annual temperature is 18 °C; May is the hottest month with a maximum temperature of 40 °C, while December and January are the coldest months with temperatures dropping to -4 °C (Comisión Nacional del Agua [CONAGUA], 2020).
The dominant soil type is Leptosol (Instituto Nacional de Estadística y Geografía [INEGI], 2023), where submontane scrub vegetation thrives, and P. pinceana stands are associated with Helietta parvifolia (A. Gray) Benth., Senegalia roemeriana (Scheele) Britton & Rose, Mimosa aculeaticarpa Ortega, Opuntia engelmannii Salm-Dyck ex Engelm., Bernandia myricifolia (Scheele) S. Wats., Jatropha dioica Sessé, Bursera fagaroides Engl., Dioon angustifolium Miq., Brahea dulcis (Kunth) Mart., Hechtia hernandez-sandovalii Ramírez, C. F. Jiménez and Treviño and Lantana camara L.
Field sampling
In June 2020, specimens of weeping pinyon were located in the ejido of Magdaleno Aguilar, and through selective sampling, 50 mature and healthy trees were chosen-meaning they showed no signs of pests, forest fire damage, or mechanical injury. Two to three cores were extracted from each tree using Pressler borers (Haglöf brand) that were 51 cm long with internal diameters of 5 and 12 mm. Samples were taken at 1.3 meters above the ground, following the slope and perpendicular to the trunk of the tree. Each sample was labeled with the site code, tree number, and subsample designation. Diameter at breast height of the corresponding tree (at 1.30 m), location coordinates, altitude, and site exposure were also recorded.
Sample processing, measurement and dating of tree rings
Cores were placed in grooved wooden molds and fixed with glue and tape. Subsequently, they were polished with sandpaper of different grit thickness (120 to 1500) to highlight the tree rings. Based on standard dendrochronological techniques (Stokes & Smiley, 1968) and with a high-resolution stereoscope, tree rings of each sample were counted, and a tentative date was assigned to the year of formation (pre-dated).
Earlywood and latewood widths were measured in each tree ring to determine the total (annual) ring width used in the rest of the analyses. These measurements were made at the Forest Ecology and Dendrochronology Laboratory of the Faculty of Engineering and Sciences of the Universidad Autónoma de Tamaulipas, where a Velmex System with 0.001 mm precision and the Measure J2X software (Speer, 2010; Velmex Inc., 2023) were used. The quality of the dating was verified with the COFECHA program. This program statistically compares the dating of each sample through the measurement files of each tree-ring width series by successively correlating segments of 50 years with an overlap of 25 years. COFECHA identifies segments of the tree-ring width series that are poorly correlated with the master chronology series, for which it shows dating errors, measurement errors, or growth problems; this process allows the assignment of the exact date to each of the tree rings (Grissino-Mayer, 2001; Holmes, 1983).
Development of chronology and determination of dendrochronological potential
To determine the dendrochronological potential, it is necessary to standardize the tree-ring width measurement values beforehand, i.e., to remove growth trends due to the age of the trees and underlying geometric effect, among other environmental factors unrelated to climate. This procedure was carried out using the ARSTAN program (Cook & Holmes, 1999).
A double standardization of the tree-rings width was used, which consisted of first applying a negative exponential curve or a linear regression line to each tree-ring width series according to its growth pattern. For better fitting, a second standardization was performed in which a cubic spline curve was applied that preserved 50 % of the variance contained in the series.
In each case, standardization consisted of dividing the annual tree-ring width by the value of the fitting curve, transforming these increments into ring width indices (RWI), dimensionless values that enable the comparison of growth series from different ages. Once RWI for all samples was collected, a chronology was developed; that is, a temporal sequence that shows the mean RWI for each year.
ARSTAN has three versions of the chronology: standard, residual and arstan. In this study we chose to work with the standard version, in which a biweight robust mean is applied. This statistic is used to remove endogenous effects originated by stand disturbance and maximize the common climatic signal contained in the tree rings. Finally, a 10-year spline curve was applied to the chronology to highlight low frequency events.
The basic statistics describing the standard chronology of P. pinceana and defining its dendrochronological potential were also derived from ARSTAN. These are intercorrelation between series (indicating the intensity of the climatic signal that is common at the tree population level), mean sensitivity (relative change in ring width from one year to the next), standard deviation (variation in ring width growth), first order autocorrelation (influence of previous ring growth on the ring width of the following year; low values reflect greater variation in growth from one year to the next) and the signal-to-noise ratio (ratio of the climatic signal to other non-climate related factors). Additionally, two statistics that depend on the depth of the sample along the chronology were obtained to accurately highlight the climatic sensitivity of the P. pinceana chronology. These are Rbar (running between average relationship) and EPS (expressed population signal), which were analyzed with the dplR package in R (Bunn et al., 2024). The Rbar is the average correlation between all series in 100-year intervals with 50 years of overlap. Being a continuous correlation between series, it is a good measure of the intensity of the common climatic signal between samples over the chronology (Speer, 2010). EPS indicates the intensity of the climatic signal expressed by the population as it approaches the longest part of the chronology. For EPS, a value of 0.85 is used as a significant point, from the year in which this value is exceeded and, in conjunction with the other dendrochronological parameters, the chronology has greater potential to be used for climate reconstruction purposes (Buras, 2017).
Relationship between radial growth of Pinus pinceana and regional climate
The influence of climate on radial growth of P. pinceana was analyzed through a correlation matrix between annual RWI series and series of regional average monthly values of precipitation and temperature (maximum and minimum). Prior to a Shapiro-Wilk normality test of the data series involved, which was satisfactory, the 'treeclim' package in R was used, with a Pearson bootstrap correlation, which was used to determine the months of precipitation and temperature that significantly influence tree-rings width, thereby supporting the climate sensitivity of weeping pinyon (Zang & Biondi, 2015).
Precipitation and temperature data were collected from weather stations near the study site that had reliable and complete climate records. Five stations met these requirements over a common period from 1981 to 2018 (38 years). The climatic data were averaged to estimate monthly precipitation and temperature (maximum and minimum) values at the regional level (Table 1).
Table 1 Weather stations near the P. pinceana sampling site in Tamaulipas.
| Estación | Latitude N | Longitude W | Altitude (m) | Period (years) | Distance (km) |
|---|---|---|---|---|---|
| Magdaleno Aguilar | 23º 27' 13'' | -99º 30' 33'' | 1 430 | 1981-2019 (39) | 1.9 |
| Palmillas | 23º 18' 00'' | -99º 33' 00'' | 1 550 | 1961-2018 (58) | 16.6 |
| Bustamante | 23º 26' 09'' | -99º 45' 15'' | 1 600 | 1971-2019 (49) | 26.5 |
| Miquihuana | 23º 34' 26'' | -99º 45' 11'' | 1 770 | 1963-2019 (57) | 30.1 |
| Los Uvalle | 23º 34' 28'' | -99º 51' 24'' | 1 560 | 1981-2019 (39) | 39.7 |
Climatic conditions prior to the tree growing season can influence the tree-ring width of the current year. Therefore, to analyze the months in which tree radial growth responds positively or negatively to climate, we used precipitation and temperature data (maximum and minimum) from 18 consecutive months: six months prior to the growing year and 12 months during the current growing year (Arroyo-Morales et al., 2023).
Results and Discussion
Dendrochronological potential of Pinus pinceana
From a total of 50 trees sampled, 149 cores were recorded, of which 124 were dated, representing 83 % of the samples collected. Twenty-five samples were discarded, corresponding to 17 %, because they showed growth problems and a high amount of resin. Even the samples that could be dated amounted to 72 missing rings representing 1.50 % of the total. The presence of missing rings is a common dendrochronological characteristic in P. pinceana and is consistent with the populations analyzed by Santillán-Hernández et al. (2010) in the latitudinal gradient of natural distribution in the Sierra Madre Oriental (north-south), whose chronologies recorded between 0.43 and 3.68 % of missing rings (Table 2).
Table 2 Missing rings detected in the dated series of Pinus pinceana population in Tamaulipas, compared to the Coahuila, Zacatecas, San Luis Potosí, Querétaro and Hidalgo populations (Santillán-Hernández et al., 2010). Populations are arranged north-south of the Sierra Madre Oriental.
| State | Population | Dated samples | Serie length (years) | Missing rings (%) | Mean anual precipitation (mm) |
|---|---|---|---|---|---|
| Tamaulipas | Magdaleno Aguilar | 124 | 153 | 1.50 | 400-600 |
| Coahuila | Las Norias | 43 | 106 | 0.43 | 350-400 |
| Palmas Altas | 25 | 205 | 2.26 | ||
| Zacatecas | Cañón de las Bocas | 46 | 125 | 1.27 | 350-400 |
| San José Carbonerillas | 43 | 157 | 2.93 | ||
| San Luis Potosí | Matehualilla | 33 | 125 | 3.53 | 400-500 |
| La Trinidad | 49 | 131 | 3.68 | ||
| Querétaro | Maguey Verde | 16 | 135 | 2.68 | 400-600 |
| El Tepozán | 6 | 57 | 0.00 | ||
| Hidalgo | San Cristóbal | 14 | 49 | 0.00 | 400-600 |
| *El Arenalito | 0 | 0 | 0.00 |
*Dating of El Arenalito population was not possible, because no growth pattern was found.
Table 2 shows that most populations of P. pinceana have missing rings in their growth series. Sites with lower precipitation such as Coahuila and Zacatecas (350-400 mm) and San Luis Potosí (400-500) have the highest percentages in this category. Although missing rings reflect the climatic sensitivity of the trees, this high variability represents a greater difficulty for the dating of the samples. Thus, problems in the radial growth of the trees can have a negative impact on the parameters that define their dendrochronological potential.
Despite the challenge of dating the radial growth of P. pinceana, the dendrochronological parameters presented in Table 3 showed that this species has the potential to record climatic variability in its tree-rings. This is supported by an intercorrelation between series of 0.54 (p < 0.001); that is, higher than the correlation value required by the COFECHA program (0.328, p < 0.01) for correct dating (Grissino-Mayer, 2001). Mean sensitivity, standard deviation, first-order autocorrelation, signal-to-noise ratio and Rbar values indicate high interannual variability in tree rings width (Grissino-Mayer, 2001; Speer, 2010). These dendrochronological parameters support the climatic sensitivity of P. pinceana from 1867 to 2019 (153 years), for which an EPS > 0.85 was determined from 1876, a year from which climatic reconstruction could be performed (Table 3; Figure 2).
Table 3 Dendrochronological parameters of the tree cores series of Pinus pinceana in Magdaleno Aguilar, Jaumave, Tamaulipas, Mexico.
| Statistical parameter | Value | *Reference value |
|---|---|---|
| Series intercorrelation | 0.54 | 0.40 a 0.90 |
| Mean sensitivity | 0.31 | 0.15 a 0.65 |
| Standard deviation | 0.33 | 0.30 a 0.80 |
| First-order autocorrelation | 0.28 | 0.60 a 0.80 |
| Signal-to-noise ratio | 43.18 | **28.90-34.63 |
| Rbar (total) | 0.34 | --- |
| EPS (Cores/Trees) | ***1876-2018 (4/4) | >0.85 |
| Chronology length (years) (Cores/Trees) | 1867-2019 (153 years) (124/46) |
*Reference value Grissino-Mayer, (2001), for the first four statistical parameters. **High values are recommended (Santillán-Hernández et al., 2010; Villanueva et al., 2018). ***Year from which the sample size is sufficiently robust for climate reconstructions. --- The reference value for Rbar (running between average relationship) is unique for each chronology, since it depends on the depth sample. EPS: Expressed Population Signal.
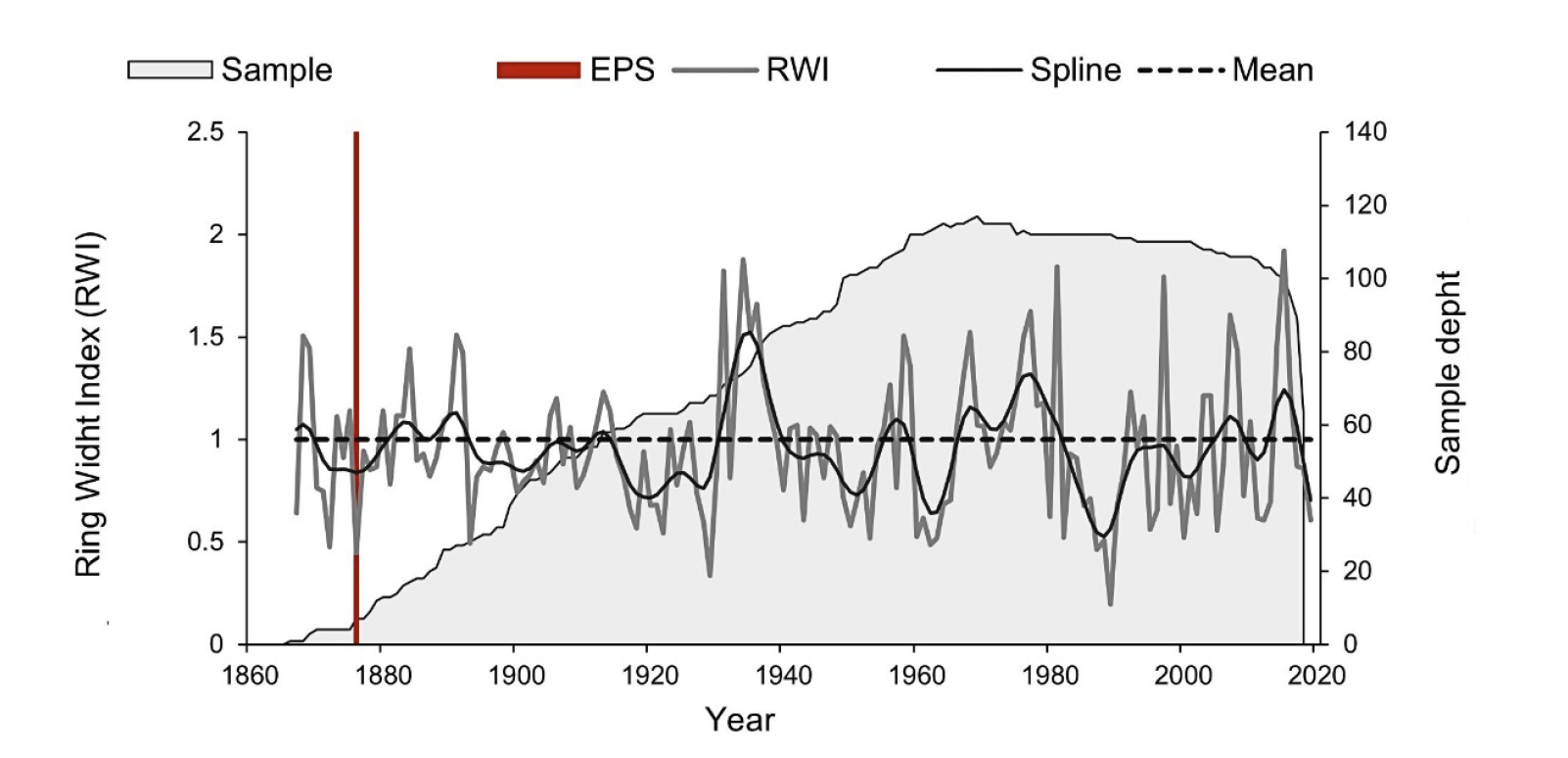
Figure 2 Annual ring width indices (RWI) chronology of Pinus pinceana period 1867-2019 (153 years). An EPS (Expressed Population Signal) > 0.85 was obtained from 1876. Spline stands the low frequency events at decadal level; The mean for chronology is approximately 1.0. Sample depth is the number of growth cores included in each annual value.
According to Table 4, the basic statistical criteria that define the dendrochronological potential of P. pinceana in Magdaleno Aguilar, Tamaulipas, show that this species presents lower values of intercorrelation, mean sensitivity and standard deviation compared to those recorded by Santillán-Hernández et al. (2010); however, a very important aspect to consider is the number of samples analyzed for the Magdaleno Aguilar site (124 growth cores), which doubles the sample size used by Santillán-Hernández et al. (2010) (Table 2). This feature largely supports the value of the dendrochronological parameters of this study and enhances the importance of the low autocorrelation value, which was similar to that recorded in Coahuila, Zacatecas and San Luis Potosí. As for the signal-to-noise ratio, this value was higher than that recorded in all the populations analyzed by Santillán-Hernández et al. (2010) (Table 4).
Table 4 Basic dendrochronological parameters of Pinus pinceana in Tamaulipas compared to those of Coahuila, Zacatecas, San Luis Potosí, Querétaro, and Hidalgo (Santillán-Hernández et al., 2010). Populations are arranged north-south of the Sierra Madre Oriental.
| State | Population | Intercorrelation | Mean sensitivity | Standard deviation | First-order autocorrelation | Signal noise ratio |
|---|---|---|---|---|---|---|
| Tamaulipas | Magdaleno Aguilar | 0.54 | 0.31 | 0.33 | 0.28 | 43.18 |
| Coahuila | Las Norias | 0.61 | 0.53 | 0.32 | 0.21 | 14.80 |
| Palmas Altas | 0.77 | 0.59 | 0.46 | 0.28 | 30.02 | |
| Zacatecas | Cañón de las Bocas | 0.74 | 0.49 | 0.38 | 0.27 | 33.64 |
| San José Carbonerillas | 0.72 | 0.61 | 0.42 | 0.29 | 34.63 | |
| San Luis Potosí | Matehualilla | 0.69 | 0.69 | 0.45 | 0.27 | 28.90 |
| La Trinidad | 0.69 | 0.64 | 0.45 | 0.20 | 29.41 | |
| Querétaro | Maguey Verde | 0.69 | 0.58 | 0.43 | 0.23 | 11.72 |
| El Tepozán | 0.63 | 0.51 | 0.38 | 0.09 | 1.35 | |
| Hidalgo | San Cristóbal | 0.75 | 0.58 | 0.47 | 0.39 | 15.83 |
| *El Arenalito | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
*Dating of El Arenalito population was not possible, because no growth pattern was found.
The dendrochronological potential of P. pinceana in Tamaulipas, compared to that reported for P. cembroides Zucc. and P. nelsonii Shaw in the study area, also shows a lower potential, particularly in the parameters of intercorrelation between series, mean sensitivity and standard deviation. Nevertheless, P. pinceana shows better performance in first-order autocorrelation and signal-to-noise ratio (Arroyo-Morales et al., 2023), even compared to P. cembroides populations from Coahuila (Constante et al., 2009). On the other hand, P. pinceana seems to have higher climatic sensitivity than some P. cembroides populations from the central part of Mexico (Carlón et al., 2018), being superior in values of intercorrelation between series, mean sensitivity and standard deviation.
The value of the statistical criteria that determine the dendrochronological potential of a species can vary depending on the geographic area, site characteristics, and the climatic variable that most limits its growth (Speer, 2010). For this reason, in addition to the geographic location of the P. pinceana populations, Table 5 shows other site characteristics such as slope exposure, slope, altitude, and amount of rainfall (climatic variable of the sites accessed) that provide additional information about the dendrochronological potential of the populations compared in this study.
Table 5 Site characteristics where the Pinus pinceana population is established in Tamaulipas and those studied by Santillán-Hernández et al. (2010), based on information from Villarreal et al. (2009). Populations are arranged north-south of the Sierra Madre Oriental.
| State | Population | Exposure | * Slope (%) | * Altitude (m) | Annual precipitation (mm) |
|---|---|---|---|---|---|
| Tamaulipas | Magdaleno Aguilar | N | 41 | 1 200 | 400-600 |
| Coahuila | Las Norias | N | 11 | 1 800 | 350-400 |
| Palmas Altas | S | 20 | 2 300 | ||
| Zacatecas | Cañón de las Bocas | N | 33 | 2 500 | 350-400 |
| San José Carbonerillas | N | 23 | 2 250 | ||
| San Luis Potosí | Matehualilla | N | 11 | 2 000 | 400-500 |
| La Trinidad | N | 39 | 1 950 | ||
| Querétaro | Maguey Verde | N | 18 | 2 200 | 400-600 |
| El Tepozán | S | 25 | 2 200 | ||
| Hidalgo | San Cristóbal | S | 35 | 1 750 | 400-600 |
| El Arenalito | S | 24 | 1 900 |
*Mean value
Table 5 indicates that most populations of P. pinceana are established on slopes with northern exposure and steep slopes, which is consistent with that reported by López et al. (2018), according to the potential distribution of this species in northeastern part of Mexico. The populations of Coahuila, Zacatecas and San Luis Potosí, located at higher altitudes and with lower precipitation, show greater dendrochronological potential, because these have higher values of intercorrelation between series, mean sensitivity and signal-to-noise ratio (Table 4). The dendrochronological potential of P. pinceana, in relation to elevation and precipitation of the sites where the species is established in Coahuila, Zacatecas and San Luis Potosí, is similar to that recorded for P. cembroides and P. nelsonii in sites with elevations between 1 900 and 2 000, and 1 950 and 2 250 m, respectively, with mean annual precipitation between 200 and 500 mm in Miquihuana, Tamaulipas (Arroyo-Morales et al., 2023).
Table 5 also shows that populations that grow at lower altitudes, with higher precipitation and, preferably, with southern exposure, have lower dendrochronological potential, as evidenced by the populations of Querétaro and Hidalgo, which show lower values of intercorrelation between series, mean sensitivity and signal-to-noise ratio. It is even possible not to find a defined growth pattern in the trees, as in the case of El Arenalito in Hidalgo (Table 4; Santillán-Hernández et al., 2010). In this regard, the low values of intercorrelation and mean sensitivity of the P. pinceana population in Tamaulipas are consistent with the characteristics of lower elevation and higher precipitation, highlighting this population for recording the lowest average elevation where the species grows (1 200 m) (Table 5).
The value of the signal-to-noise ratio of the P. pinceana population in Tamaulipas is higher than that recorded with the populations compared (Table 4), even for P. nelsonii that inhabits near the study area (Arroyo-Morales et al., 2023). This suggests that, in addition to the altitude and precipitation characteristics of the site, other factors such as the slope of the hillside and perhaps the type of soil influence the moisture retention capacity; therefore, these are factors to consider in the climatic sensitivity of trees. Meanwhile, the evidence so far indicates that P. pinceana trees in Magdaleno Aguilar, Tamaulipas, are found on slopes that are on average 41 % steep, the highest value recorded among the populations analyzed (Table 5).
Based on the dendrochronological values of P. pinceana in the study area, which exceed the average values suggested by Grissino-Mayer (2001), especially those of autocorrelation and signal-to-noise ratio, it is supported that the radial growth of weeping pinyon in Tamaulipas is sensitive to climate. Therefore, the species has adequate dendrochronological potential; that is, the tree rings width of P. pinceana in Magdaleno Aguilar, Tamaulipas, is controlled mainly by climatic conditions.
Climatic response of Pinus pinceana radial growth
The relationship between regional precipitation and radial growth of weeping pinyon for the common data period 1981-2018 (38 years) showed positive and significant response in the months of September of the previous year of growth and March, April and May of the current year. However, with the accumulated precipitation it is shown that the period of greatest influence on the radial growth of P. pinceana is from September of the previous year of growth to May of the current year (r = 0.71; p < 0.001); the coefficient of determination (r2 = 0.50) indicates that 50 % of the radial increase is explained by the autumn-spring precipitation. On the other hand, the most influential cumulative precipitation during the current growing year is March-May (r = 0.60; p < 0.001), where the coefficient of determination (r2 = 0.36) shows that 36 % of the radial increment is explained by spring precipitation (Figure 3a).
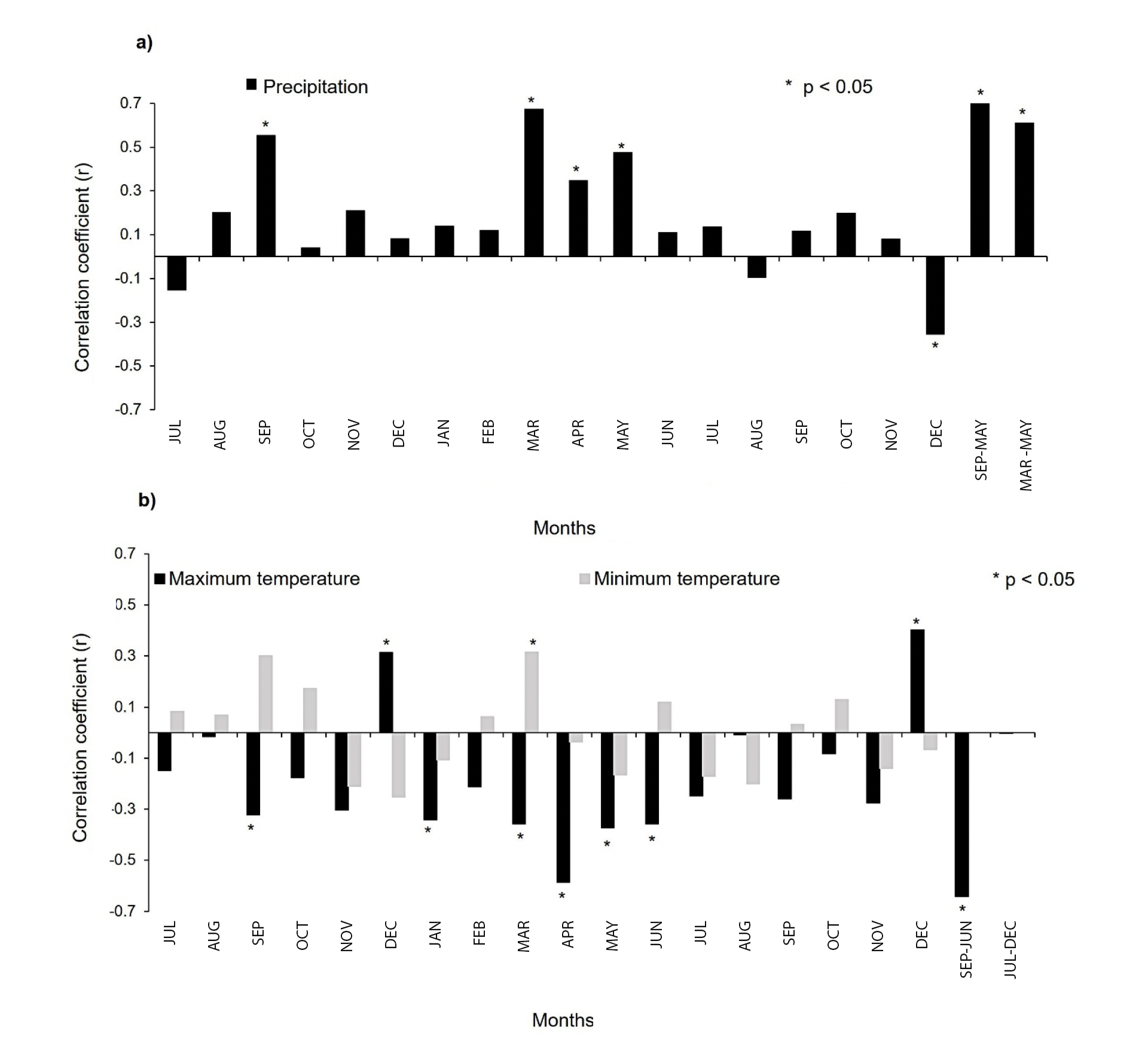
Figure 3 Influence of regional climate on the radial growth of Pinus pinceana. Correlation coefficient between the annual ring width indices and 18 consecutive months of a) precipitation and b) temperature (maximum and minimum). Lowercase letters indicate previous year months; capital letters, current year months.
The relationship between regional temperature and radial growth of P. pinceana shows a negative response. Compared to minimum temperature, maximum temperature has a greater influence on the radial growth of weeping pinyon (Figure 3). This climatic variable significantly limits radial growth in September of the previous growth year and in January, March, April, May, and June of the current year. However, a positive and significant influence was detected in December of both the previous and current years, when in both cases, the maximum value does not exceed 32 °C (Figure 3b).
The period of maximum temperature with the greatest negative and significant influence is from September of the previous year to June of the current year (r = -0.64; p < 0.001) and the coefficient of determination (r2 = 0.40) indicates that 40 % of the decrease in the width of the rings of P. pinceana is explained by the maximum temperature in autumn and early summer. On the other hand, the most influential period of maximum temperature during the current growing year is March-June (r = -0.53; p < 0.001), where the coefficient of determination (r2 = 0.28) shows that 28 % of the decrease in ring width is explained by the maximum temperature of spring and early summer (Figure 3b).
The relationship between minimum temperature and radial growth of P. pinceana showed an equal number of negative and positive responses; however, only the positive correlation in March of the current year of growth was significant (r = 0.31; p < 0.05), when the maximum minimum temperature value in this month is 6.6 °C. Since no significant seasonal period of minimum temperature was detected, the period of 18 consecutive months of minimum temperature was correlated with the radial growth of P. pinceana. Thus, a non-significant negative correlation (r = -0.00078; p > 0.05) of minimum temperature on radial growth of P. pinceana was recorded (Figure 3b).
In Tamaulipas, the response of P. pinceana to precipitation shows that this variable is the most limiting factor for the radial growth of the species, where precipitation in September, which accumulates with that from October to February, enhances its radial growth during March, April and May (Figure 3). This climatic response is consistent with the documented evidence for radial growth of P. cembroides and P. nelsonii in the study area (Arroyo-Morales et al., 2023).
The radial growth response of weeping pinyon in Tamaulipas is consistent with the general response of conifers in the northeastern part of Mexico, which do not react immediately to torrential rains such as those of September but need moisture to be available in the soil for later use. Thus, the moisture available in the soil profile as a result of the September rains, combine with the moisture that infiltrates during the winter after cold fronts, is used when trees emerge from dormancy and reactivate their metabolic functions during the spring growing season (Arroyo-Morales et al., 2023; Constante et al., 2009).
Although it has been demonstrated in this study that precipitation has a greater influence on the radial growth of P. pinceana, this does not mean that this variable acts independently of temperature. Thus, the gradual decrease of this climatic factor from September until winter also allows a lower degree of evapotranspiration in the region. Thus, at the beginning of the growing season in March, precipitation in this month stands out among all the months where P. pinceana has a positive response (Figure 3a). This could indicate an adequate combination between the available soil moisture and the environmental temperature that begins to increase in March but is not yet stressful for the physiology of the trees (the extreme value of maximum temperature in February is 34.6 and 37.8 °C in March, compared to 39.2 and 40.8 °C in April and May, respectively).
Since the radial growth of conifers in the northeastern part of Mexico generally responds better to precipitation, there is more information available on the interaction between these two variables. However, to date, the only dendroclimatic study on P. pinceana that addresses its response to precipitation is that of Santillán-Hernández et al. (2010). Therefore, the present study relies on this reference to compare the species' response to the common climate variable.
Santillán-Hernández et al. (2010) report that the radial growth of P. pinceana populations in northeastern Mexico (Coahuila) respond significantly to rainfall in December of the previous year, and February, May and July of the current year. Populations in central-northern Mexico, represented by the states of Zacatecas and San Luis Potosí, respond to rainfall in December of the previous year, and January, February, March, May and July of the current year. In Tamaulipas, the response of P. pinceana to rainfall (this study) only finds synchrony with the results of Santillán-Hernández et al. (2010), in the positive and significant response to the months of March and May of the current year of growth (Table 6). Regarding the response to a period of accumulated rainfall, the populations analyzed by Santillán-Hernández et al. (2010) show sensitivity to seasonal rainfall from January of the previous year of growth to July of the current year; that is, they are consistent with winter rainfall in early summer. On the other hand, the weeping pinyon in Tamaulipas shows to be more responsive to precipitation from September of the previous year of growth to May of the current year; that is, an autumn-spring period and in lower altitude zones (Figure 3a; Table 6).
Table 6 Influence of precipitation on the radial growth of P. pinceana in Tamaulipas and those studied by Santillán-Hernández et al. (2010) in Coahuila, Zacatecas and San Luis Potosí. Populations are arranged north-south of the Sierra Madre Oriental.
| State | Mean altitude (m) | Months with significant influence | Response period |
|---|---|---|---|
| Tamaulipas | 1 200 | September, March, April and May | Autumn-Spring (September-May) |
| Coahuila | 2 050 | December, February, May and July | Winter-Summer (January-July) |
| Zacatecas y San Luis Potosí | 2 175 | December, January, February, March, May and July | Winter-Summer (January-July) |
The radial growth response of P. pinceana, from a cumulative precipitation starting in September of the previous year, differs from that documented for other pinyon species such as P. cembroides and P. nelsonii, which usually respond starting in November or December of the previous year (Arroyo-Morales et al., 2023; Carlón et al., 2018; Constante et al., 2009). However, the response of P. pinceana to precipitation in the current year (March-May) is consistent with the behavior of P. cembroides and P. nelsonii in the study area (Arroyo-Morales et al., 2023). Thus, once the soil has available moisture (from the previous year's precipitation) during March, April and May (months of higher solar radiation and high temperatures), a minimum amount of rainfall (24 % of the annual precipitation in spring) is what enhances the growth of P. pinceana (CONAGUA, 2020).
The response of P. pinceana to precipitation in the southwestern part of Tamaulipas reflects precipitation conditions outside of the intra-summer drought or midsummer drought period that naturally occurs between July and August in the study area (Corrales-Suastegui et al., 2020; Sánchez-Santillán et al., 2018). Thus, the potential of the weeping pinyon to record precipitation variability during the seasonal period from September of the previous year to May of the current year is demonstrated. This information could be useful for further studies of precipitation reconstruction and droughts that have occurred in the region (Stahle et al., 2016); however, it is recommended to increase the number of dendroclimatic studies that address P. pinceana in Tamaulipas, to verify if the same response to climatic variables is maintained, once more populations are incorporated at different elevations.
Conclusions
This study represents the first dendroclimatic research that addresses Pinus pinceana in Tamaulipas and provides 153 years of information (1867-2019) on variability of radial growth, which is determined by both maximum temperature and precipitation with the latter being the more significant factor. Therefore, the dendrochronological potential of P. pinceana is demonstrated as an alternative source of long-term climatic information. This knowledge can be applied in future studies, not only to reconstruct the region's historical precipitation and analyze drought patterns but also to improve water resource management and support conservation efforts for this endangered species.

