











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Global trends indicate that human activities have significantly impacted biodiversity, particularly in the past five decades (Díaz et al. 2019). Currently, mammals are the second most endangered vertebrate group, comprising 22 % of threatened species (IUCN 2020). Among mammals, 15 % of bat species are threatened (Frick et al. 2020). Although bats have a global distribution and high diversity in tropical regions (Altringham 2011; Burgin et al. 2018), many species are sensitive to habitat disturbances (Voigt and Kingston 2016).
The Curaçaoan Long-nosed bat, Leptonycteris curasoae Miller, 1900, is one of the 19 threatened bat species in South America and the Caribbean islands. This species strongly depends on dry forests and arid ecosystems in Colombia, Venezuela, and the Caribbean islands (IUCN 2023). This fruit- and nectar-feeding bat species is the primary pollinator of several plant species, mainly of Cactaceae (e. g., Stenocereus spp., Cereus spp., Pilosocereus spp.) and Asparagaceae (Agave spp.) families (Griffiths and Gardner 2007). L. curasoae is a migratory bat, undertaking long-distance flights (28 to 89 km) between Caribbean islands, including Aruba, Bonaire, and Curaçao, and between those islands and the mainland of northern Venezuela (Simal et al. 2015).
The main threats for L. curasoae are habitat alteration due to urban expansion, mining, quarrying, wind farms development, and vandalism of roosts, mostly caves (Nassar 2015; Simal et al. 2021). Furthermore, dry forests, arid and semiarid zones are the most threatened and transformed ecosystems in the Neotropics (Escribano-Avila et al. 2017), with 66% of the dry forests having already disappeared (Portillo-Quintero and Sánchez-Azofeifa 2010). In Colombia, Venezuela, and the Caribbean islands, these ecosystems are in critical condition because they are underrepresented in protected areas. In Colombia and Venezuela, only 5 % of their coverage is under protection (Fajardo et al. 2005; Rodríguez et al. 2010; Pizano and García 2014), and in Caribbean islands, approximately 10 % of these ecosystems are within protected areas (Portillo-Quintero and Sánchez-Azofeifa 2010).
Most studies assessing habitat and feeding ecology, geographic range, roost location, and populational trends of L. curasoae have been carried out in the islands of Aruba, Bonaire, and Curaçao (Petit 1995, 1997; Simal et al. 2015, 2021) and Venezuela (Martino et al. 1998, 2002; Nassar et al. 2003; Newton et al. 2003). Much less information is available for Colombia (Cadena et al. 1998; Sánchez and Cadena 1999). The above topics are vital for updating the risk of extinction assessment provided by the IUCN Red List nearly a decade ago for L. curasoae (Nassar 2015). Information regarding long-distance routes, stopover foraging grounds, and diurnal and mating/maternity roosts along its potential seasonal migration paths remains scarce across L. curasoae´s entire geographical distribution, limiting our understanding of the annual migration dynamics attributed to this species.
Niche-based species distribution modeling (SDM) has proved to be valuable in conservation strategies (Rodríguez et al. 2007; Villero et al. 2017). Such modeling is particularly useful for locating suitable areas for field exploration and identifying key variables limiting the distribution of rare bat species of conservation concern (Razgour et al. 2011). In this study, we aimed 1) to generate a potential distribution map of L. curasoae, 2) to identify threats and under-protected areas within this bat's potential distribution, and 3) to fill in information gaps concerning the current distribution and roosts occurrence for the species, emphasizing understudied countries such as Colombia.
Materials and methods
Study area. We compiled data from all countries where L. curasoae is distributed: Colombia, Venezuela, Aruba, Bonaire, and Curaçao islands (ABC islands). Due to the limited information available on the current distribution of L. curasoae in Colombia, we conducted field expeditions in 2020, 2022, and 2023 according to the reproductive cycle and the possible annual migration of the species (Martino et al. 1998; Simal et al. 2021). Field expeditions were conducted in 27 locations across three geographic areas: 1) The Chicamocha River basin (ten sites) in January 2020. 2) the Caribbean coast (nine sites) in February 2020. 3) Tierra Bomba island in February 2020, April and December 2022 and June 2023. 4) La Guajira Peninsula (seven sites, Supplementary material Figure S1) in April 2022.
The Chicamocha River Basin is in the Colombian Eastern Mountain range, at 500 to 1,500 masl (Albesiano et al. 2003). This area features semi-arid, thorn scrub, and dry forest vegetation (Latorre et al. 2014). The mean annual temperature in this region ranges from 16 ºC to 36 ºC, and annual rainfall reaches 731 mm (Albesiano et al. 2003). Sampled localities in the Caribbean coast and La Guajira Peninsula belong to the Peri-Caribbean Arid Belt. The Caribbean coast is characterized by seasonally tropical dry forests, with an average annual temperature of 27 °C (16 to 36 °C, IDEAM 2010), while La Guajira Peninsula primarily comprises xerophytic and thorn scrub vegetation, with an average annual temperature of 29 °C (19 to 36 °C, IDEAM 2010). The Peri-Caribbean Arid Belt province generally has lower average yearly rainfall, strong winds, and higher evaporation rates than the Chicamocha River Basin (Hernández et al. 1992).
Current geographic distribution of Leptonycteris curasoae. We mapped the current geographical distribution of L. curasoae revisiting information on specimens deposited in mammal collections, including the Instituto de Ciencias Naturales of Universidad Nacional de Colombia (ICN), Instituto Alexander von Humboldt (IAvH), Universidad Industrial de Santander (UIS-MHN), and Vertebrate collection at Universidad de Los Andes, Venezuela (CVULA). Additional records were obtained from other institutions by revising databases such as GBIF (GBIF 2022) and VertNet (VertNet 2022). Furthermore, we examined information about geographical distribution in the scientific literature in Google Scholar (Google Scholar 2022) and Web of Science (Web of Science 2022) using the keywords “Leptonycteris* AND L. curasoae* AND Curaçaoan Long-nosed Bat*” without date restrictions. Also, we checked data compiled in the thesis and technical reports of studies carried out in zones where this bat species was historically found. This resulted in a total of 13 documents conducted in the five countries where L. curasoae is distributed. We assessed the Extent of Occurrence (EOO) and the Area of Occupancy (AOO) for the revisited current distribution that we obtained and for the previous distribution published in the last IUCN Red List assessment (Nassar 2015), using the sRedList platform (Cazalis et al. 2024).
As described earlier, we complemented the current distribution for Colombia conducting field expeditions across 27 locations. At each location, bat sampling was carried out using six ground-level mist nets (two 6 m, one 9 m, and three 12 m length, 20 mm mesh, ECOTONE, Poland). Mist nets were active from 18:00 to 00:00 and were checked every 25 minutes on average. Our total sampling effort was 16,609 m2 x mist net x hours (mnh). Also, we searched for caves and artificial roosts where bats were captured using mist nets at the entrances of the roosts or an entomological hand net inside the roosts. Capture and handling methods followed the guidelines of the American Society of Mammalogists (Sikes et al. 2011) and were authorized by licenses from the Environmental Licenses National Agency (ANLA 2014).
Ecological modeling and climate variables. We used 19 current bioclimatic variables sourced from the Chelsa database (Karger et al. 2021), which offers data at approximately 1 km² spatial resolution (0.0083 arc seconds). These variables depict annual and seasonal climate patterns over the baseline period from 1970 to 2000. To address collinearity issues among the 19 bioclimatic variables, we employed the corrSelect function from the fuzzySim package (Barbosa 2015), available in R (R Core Team 2022). By using the Pearson method with a threshold value set at 0.8 and a variance inflation factor, this function effectively identifies and excludes highly correlated variables (Barbosa 2015). To establish a calibration area (M), we intersected a buffer around species occurrences (2.5 °) with ecoregions defined by Olson et al. (2001). This approach considers both dispersal and ecological limitations, enhancing the robustness of our modeling process.
Occurrence records. We generated a database comprising 400 georeferenced records for L. curasoae and after a thorough examination, we excluded 63 % of the records due to inconsistent or duplicated coordinates. Furthermore, we implemented a distance-based filter of 5 km to mitigate sample bias, using the spThin function (Aiello-Lammens et al. 2015) of the Wallace package (Kass et al. 2018, 2023). As a result of these procedures, we obtained 148 valid records.
Niche-based species distribution model. We employed the ensemble modeling approach implemented in the biomod2 package (Thuiller et al. 2009) within the R software (R Core Team 2022) to develop the niche-based distribution model for L. curasoae. Ensemble modeling combines multiple individual models to produce a more robust prediction by leveraging the strengths of each constituent model while mitigating the effects of their inherent uncertainties and errors (Araujo and New 2007).
Out of the ten algorithms available in biomod2, we selected seven for our analysis: MaxEnt (MAXENT.Phillips), Generalized Additive Models (GAM), Generalized Boosted Regression Model (GBM), Artificial Neural Networks (ANN), Surface Range Envelope (SRE), and Random Forest (RF). Because our occurrence datasets only included presence-derived information, we randomly generated 10,000 pseudo-absences within the calibration area to balance the dataset. Prevalence was set to 0.5 to assign equal importance to presences and pseudo-absences during the calibration process. Each model was executed ten times, with each run using a different selection of calibration and evaluation datasets, with 70 % of the data allocated for calibration and the remaining 30 % for evaluation in each run.
To evaluate model performance, we used the area under the curve of the receiver operating characteristic (AUC). Models were assembled using a total consensus rule, where algorithms and the ten replicates were assembled based on the weighted sum of evaluations greater than AUC > 0.8 (Araujo and New 2007). Subsequently, an ensemble model comprising seven algorithms × 10 repetitions was used to project the potential distribution of L. curasoae under current climatic conditions. Given the thorough revision of all occurrence records (Peterson et al. 2011), we are confident that each point contributes to representing the distribution of L. curasoae. Therefore, we obtained a binary map by employing the minimum training presence threshold (value of 240).
Pressures and threats. We conducted an overlay analysis, superimposing the current and potential spatial distribution of L. curasoae with layers depicting land use transformation and protected areas, and mapped current threats such as mining, wind farms, tourism, and vandalism of roosts across the geographic range of this species. The land-use transformation data were obtained from a global map of land use/land cover (LULC) from 2023 derived from ESA Sentinel-2 imagery at 10 m resolution (Karra et al. 2021). This map layer was created based on a large dataset of over 5 billion human-labeled Sentinel-2 pixels by developing and deploying a deep-learning segmentation model on Sentinel-2 data. The algorithm generates LULC predictions for nine classes: water, trees, flooded vegetation, crops, built area, bare ground, snow/ice, clouds, and rangeland.
The polygons representing protected areas were sourced from Protected Planet (Protected Planet 2023), a comprehensive data repository on protected areas and other effective area-based conservation measures (OECMs). This database is updated monthly and maintained by the United Nations Environment Programme World Conservation Monitoring Centre (UNEP-WCMC 2024). Only protected areas categorized by the IUCN were considered. Protected areas designated as ‘Protective Zones’ and ‘Critical Areas with Treatment Priority’ were excluded from the analysis because they only exist in Venezuela, and the anthropogenic activities allowed include exploitation of natural resources incompatible with the aims of protected areas (García and Silva 2013). We compiled 340 georeferenced locations of mining, wind farms, and tourism threats obtained from Colombian governmental platforms such as the Mining and Energy Planning Unit (2023) and the Ministry of Commerce, Industry, and Tourism (2022), respectively. For Aruba, Bonaire, Curaçao, and Venezuela, threat information came from published data (Petit et al. 2006; Molinari et al. 2012; Nassar and Simal 2019; Simal et al. 2021) and data observed directly in the field between 1997 and 2015 (J. Nassar pers. obs.). Similarly, roost vandalism records for Colombia were obtained from our field observations from 2020 to 2023.
By integrating spatial data on land cover change, we identified regions where habitat degradation and fragmentation might pose significant threats to L. curasoae populations. Additionally, overlaying data of protected areas allowed us to assess the extent to which current conservation measures cover the bat's distribution range and identify potengial gaps in its protection. Mapping current threats enabled us to pinpoint specific areas where human activities pose immediate risks to the survival and habitat integrity of L. curasoae.
Results
Current geographic distribution of L. curasoae. Most of the occurrence records of L. curasoae came from Venezuela (41.2 %) and Colombia (39.9 %), and the remaining occurrence data (18.9 %) correspond to the ABC islands (Supplementary material, Table S1). In Colombia, we captured 200 individuals from six localities in the three geographic areas sampled: the Chicamocha River Basin (5 individuals), the Caribbean Coast (124), and La Guajira Peninsula (71; Table S2). Five localities represent new records of L. curasoae in the country (Table S1).
Throughout the geographic range of L. curasoae, only 23 locations (15.5 %) correspond to roosts. Ten roosts are in Venezuela, and ten are in the ABC islands. During the field expeditions in Colombia, we explored 23 roosts, including caves, natural shelters, and human-made buildings (Supplementary material, Table S3). We discovered three roosts of L. curasoae in Colombia. One roost is in a cave on La Guajira Peninsula, which hosts a large maternity colony (Table S3). The other two roosts are in historical fortresses on Tierra Bomba Island where a smaller colony resides. This colony was observed only in February 2020 and December 2022, using the fortresses as mating roosts (Table S3). Eighteen shelters are maternity roosts: Venezuela (7 shelters), Bonaire (4), Aruba (3), Curaçao (3), and Colombia (1). Mating roosts were found in Venezuela (5), Bonaire (2), and Colombia (2; Supplementary material, Table S1). Our data indicated that the Extent of Occurrence (453,300 km2) was reduced by almost half and the Area of Occupancy (167,500 km2) by nearly a third in the revisited current distribution compared to the previous distribution published in the last IUCN Red List assessment (EOO = 1,028,661km2 and AOO = 614,428 km2; Figure 1).
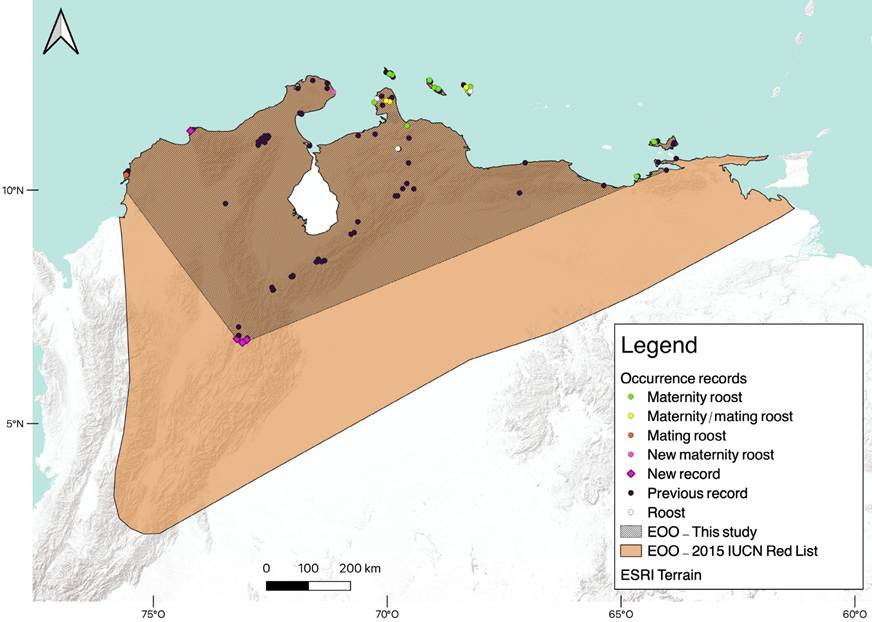
Figure 1 Records of L. curasoae used for the Species Distribution Model (SDM). New localities and types of roosts are shown. The Extent of Occurrence (EOO) assessed in this study and from the IUCN Red List (Nassar 2015) for L. curasoae are shown.
Niche-based species distribution model. Out of the 60 models created (six algorithms and ten replicates), 45 models exhibited AUC values greater than 0.8. The average sensitivity across these models was 96.5 %, indicating a high proportion of correctly predicted presences, while the average specificity was 89.4 %, reflecting a high proportion of correctly predicted absences (Supplementary material, Table S4).
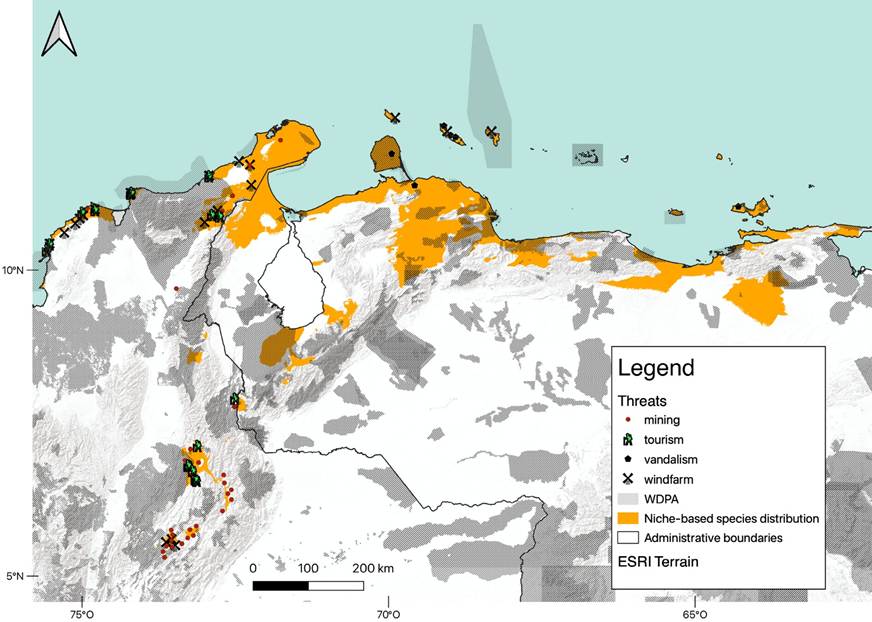
The potential distribution of L. curasoae estimated through the weighted ensemble of these models encompassed most of its currently known distribution in Colombia, Venezuela, and the ABC Islands. Predicted suitable areas were predominantly identified in Aruba, Bonaire, Curaçao, La Guajira Peninsula in Colombia, the Paraguaná Peninsula, and areas south of Falcón and north of Lara states in Venezuela, as well as several intra-Andean arid patches in Venezuela and Colombia. All these areas correspond with most of the currently known locations for this species, however, some exceptions were observed. Suitable areas for L. curasoae were identified in the northeast portion of the Sierra de Perijá, north of Monagas state, and southeast of the Paraguaná Peninsula in Venezuela, as well as in the south of Sierra del Perijá and south of the Chicamocha River Basin in Colombia. All of them are locations where L. curasoae has never been recorded in the past. Additionally, a deviation of the model was detected with no suitable area predicted for an existent record of the species in the south of the Cesar state, in Colombia (Figure 2).
The final set of non-correlated variables was seven (Table 1). Based on the model’s predictions, L. curasoae can occur in areas with relatively high annual mean temperature, low temperature seasonality, and low variation in annual mean diurnal range temperature. Also, suitable areas are characterized by low to intermediate annual precipitation, intermediate precipitation seasonality, and low precipitation in the driest month and the coldest quarter (Table 1). The potential distribution model indicated that suitable areas for this bat species represent only 9.4 % of the total area reported in the IUCN Red List (Figure 2).
Table 1 Mean, median, maximum, and minimum of bioclimatic variables considered in the potential distribution model of L. curasoae.
| Annual mean temperature (ºC) Bio 1 | Annual mean diurnal range (ºC) Bio 2 | Temperature seasonality (CV, %) Bio 4 | Annual precipitation (mm) Bio 12 | Precipitation of driest month (mm) Bio 14 | Precipitation seasonality (CV, %) Bio 15 | Precipitation of coldest quarter (mm) Bio 19 | |
|---|---|---|---|---|---|---|---|
| Minimum | 8.35 | 0.9 | 15.5 | 276 | 1.1 | 25.4 | 6.2 |
| Median | 26.35 | 7.2 | 72.8 | 919.7 | 12.2 | 60.9 | 84.3 |
| Mean | 25.91 | 6.69 | 71.13 | 902.5 | 20.18 | 58.39 | 122 |
| Maximum | 29.05 | 11.3 | 113.6 | 2356 | 124.1 | 110.6 | 574.1 |
Pressures and threats. According to the LULC map, the most dominant land-use classes were rangeland (47.6 %) and trees (40.2 %). In comparison, the anthropized area covered 8.4 % of the potential distribution area of L. curasoae (Supplementary material, Table S5). In general, 133 protected areas overlapped with the potential distribution of L. curasoae (Figure 2, Supplementary material, Table S6). Only 40 (30.1 %) protected areas have the highest level of protection, being considered as IUCN categories Ia (1 area) and II (39), which cover only 18.41 % of the potential distribution of L. curasoae. Most of the protected areas are in Venezuela (58.6 %). In Colombia, 33.1 % of the protected areas overlapped, while in the ABC islands the overlap was 8.3 %. About 22.2 % of the suitable areas predicted are inside protected areas, most of which are in Venezuela (20.2 %), followed by Colombia (1.9 %) and the ABC islands (0.1 %; Table 2).
Table 2 Potential distribution area of L. curasoae (extension and percentage), protected areas overlapped (number and percentage) with the potential distribution of L. curasoae, and percentage of potential distribution area under protection by country.
| Country | Potential distribution area (km2) | % of potential distribution area | Number and (%) of protected areas | % of potential distribution area inside protected areas |
|---|---|---|---|---|
| Aruba | 172.21 | 0.22 | 1 (0.75) | 0.04 |
| Bonaire | 273.44 | 0.35 | 3 (2.26) | 0.06 |
| Curaçao | 419.53 | 0.53 | 7 (5.26) | 0.03 |
| Colombia | 18,454.38 | 23.28 | 44 (33.08) | 1.94 |
| Venezuela | 59,936.35 | 75.62 | 78 (58.65) | 20.17 |
| Total potential distribution inside protected areas | 22.23 | |||
| Total potential distribution without protection | 77.77 | |||
| Total potential distribution | 79,255.91 |
We identified 86 threats that overlapped with the potential distribution of L. curasoae (Figure 2). Roost vandalism is the most critical threat in Curaçao and Venezuela, while mining, tourism, and wind farms are the most frequent menaces identified in Colombia. Mining and tourism were mainly concentrated in L. curasoae suitable areas in the intra-Andean, Caribbean, and La Guajira regions of Colombia. Wind farms are mainly restricted to the ABC islands (1 wind farm in each) and northern Colombia (17; Figure 2, Supplementary material, Table S7).
Discussion
The distribution of a species is expected to occur in areas with suitable abiotic conditions and biotic interactions that have been accessible to the species via dispersal over relevant time periods (Feng et al. 2024). In this study, we found that the current Extent of Occupancy and potential distribution of L. curasoae are smaller than previously considered, with environmentally suitable areas found in the ABC islands, northern Colombia and Venezuela. Around 22.3 % of the suitable areas for the species are inside protected areas, and many threats identified occur in countries where the environmentally suitable areas are less represented in national parks or reserves, such as Colombia.
Current geographic distribution of L. curasoae. In Colombia, L. curasoae is more abundant in the coastal region (Caribbean and La Guajira Peninsula) than in the Chicamocha River Basin. Similarly, more occurrences of this species were reported by Cañón and Trujillo (2014) in La Guajira. Previous studies have documented a lower capture rate of L. curasoae in intra-Andean arid patches of Colombia and Venezuela compared to northern Venezuela (Sosa and Soriano 1993; Cadena et al. 1998; Martino et al. 1998; Sánchez and Cadena 1999; Soriano et al. 2000). Part of the sampling in the Caribbean coast and La Guajira was conducted at the recently discovered roosts, where the largest colony mainly consisted of pregnant females. On Tierra Bomba Island, the colony uses the fortresses only during the mating season (October-February) and leaves afterward (A. Otálora-Ardila, unpublished data). These reproductive behaviors may explain the large aggregations and high abundance observed of L. curasoae in northern Colombia. In contrast, no roosts were found for L. curasoae in the Chicamocha River Basin.
The high abundance of L. curasoae in the northern part of its range might also be linked with food availability. Stenocereus griseus and Cereus repandus, two columnar cacti species that are primary food resources for L. curasoae, have longer flowering and fruiting periods on the coast of Venezuela compared to the continental and intra-Andean arid areas of Venezuela and Colombia (Sosa and Soriano 1996; Ruiz et al. 2000; Nassar and Emaldi 2008). These differences in cacti reproductive patterns are probably mediated by variations in rainfall regimes between the coast and continental arid zones (Nassar and Emaldi 2008). Although specific data on reproductive phenology of cacti in the north of Colombia is lacking, similar differences in resource availability might exist between the northern regions and intra-Andean arid zones.
Most of the known roosts of L. curasoae are in northern Venezuela and the ABC islands, regions characterized by abundant karst formations (Hoekstra et al. 2010). Consequently, have more potential for cave development than continental regions in northern South America. Only in the ABC islands, the karst extension is nearly 13,000 km2 (Day 2010). This high concentration of caves likely explains the large populations of L. curasoae in this part of its range. High capture rates have been recorded in caves of ABC islands and the Paraguaná Peninsula in Venezuela, with population estimates of 14,350 bats in Bonaire and 26,517 in Aruba (Simal et al. 2015, 2021). Our data suggest a similar pattern in Colombia, where a cave in La Guajira Peninsula hosts a maternity colony of around 10,000 bats. Although the two roosts we found in Tierra Bomba are human buildings (Otálora-Ardila et al. 2022a, b), they host approximately 700 individuals. These three shelters are the only roosts known for L. curasoae in Colombia. Despite exploring roosts in the Chicamocha River Basin, including La Macaregua cave, which was previously reported as L. curasoae roost (Marinkelle and Cadena 1972), we did not find this species in any cave or artificial roost in this region.
Our data indicated that the current EOO is smaller than previously known (Nassar 2015). Upon revisiting specimens at mammal collections, we identified some specimens with incorrect localization data, confirming that the southern location of L. curasoae corresponds to the Chicamocha River Basin. Although our results indicate a reduction in EOO and AOO, the current EOO and AOO are still above the thresholds of 100 km2 or 10 km2, respectively, which would be necessary to classify L. curasoae as an Endangered species. Based solely on the current EOO and AOO values, this species would be classified as Near Threatened or Least Concern. However, the A2 criteria applied to L. curasoae in the past (inferred population reduction where the causes of reduction may not have ceased, be understood, or be reversible) remain relevant. Additionally, the threats faced by L. curasoae and knowledge about its biology and population parameters vary across the countries where it is distributed. The situation is more critical in Colombia because there is limited knowledge of roosts and populational parameters. Moreover, the identified threats are present throughout its known distribution in Colombia and are expected to increase in the near future. Consequently, considering all these factors and adopting a conservative approach, we recommend that L. curasoae be maintained as Vulnerable (VU A2c).
Our data showed that the environmentally suitable areas of the potential distribution of L. curasoae correspond to only 9.4 % of the area presented in the IUCN Red List (Nassar 2015). This discrepancy arises because the Extent of Occurrence assessed by the IUCN overestimates the actual distribution because it includes an erroneous record located 170 km to the southwest, corresponding to an individual captured in the Chicamocha River Basin. Also, the Extent of Occurrence represents the area encompassing all the known occurrences of a taxon, which includes unsuitable or unoccupied habitats (IUCN 2012). In contrast, the potential distribution refines the area occupied by a species by delimiting it to suitable areas.
Niche-based species distribution model and threats for L. curasoae. Bioclimatic variables such as temperature and precipitation are recognized as environmental factors influencing the potential distribution areas of tropical bats (da Silva et al. 2018; Debata et al. 2019; Garbino et al. 2023). Specifically, temperature seasonality (Bio 4) and precipitation seasonality (Bio 15) significantly contribute to the potential distribution of other tropical nectarivorous bats associated with arid environments (Burke et al. 2019). These climatic variables are positively correlated with primary productivity, water availability, and phenology in tropical dry forests (Stan and Sanchez-Azofeifa 2019). For instance, temperature promotes flowering production in seasonally dry forests (Pau et al. 2013), while precipitation regimes influence the flowering and fruiting patterns of columnar cacti, which are primary dietary sources for L. curasoae (Nassar et al. 2003; Nassar and Emaldi 2008). Furthermore, temperature and precipitation contribute to modulate the species and functional composition of chiropterophilous cacti used by L. curasoae in the inhabited dry ecosystems (Ruiz et al. 2002; Nassar et al. 2013). Consequently, these two bioclimatic factors are key drivers of the seasonal availability of food resources for this species.
Additionally, suitable areas for L. curasoae were associated with low mean diurnal range, suggesting that temperature is an extrinsic factor that might exert physiological constraints on this species. As noted by Ortega-García et al. (2017), L. curasoae has the narrowest thermal niche among Neotropical nectar-feeding bats, ranging from 16.6 to 32.6 °C. Thus, areas with temperature stability and values within these limits provide optimal thermal conditions for this species.
The spatial distribution of threats affecting L. curasoae suggests that, despite the relatively small anthropized area (8.4 %), there are marked differences in protected area coverage and the impacts of other threats among countries. Our data indicate that wind farms, mining, and tourism pose important threats in Colombia. For instance, mining has had more impact, with approximately 90 % of Colombia’s coal production occurring at La Guajira, and 90 % of gypsum and 30 % of limestone mining operations located in the Chicamocha River Basin (UPME 2022, 2023a). Wind farms represent another potentially critical threat, particularly in the ABC islands and northern Colombia, where 67 wind farm projects are planned along the Caribbean coast and from the La Guajira Peninsula to northern Cesar state (UPME 2023b). Although the long-distance flying routes of L. curasoae in Colombia are not well known, it is likely that some of the planned wind farms could overlap with the presumed migratory corridor that this species might use from La Guajira Peninsula to the east towards the Caribbean coast and south towards the intra-Andean dry areas (UPME 2023b).
Our data indicated that only 22.2 % of the suitable areas of L. curasoae are within protected areas. Extending or creating new protected areas is a high-priority conservation action for dry forests (Sánchez-Azofeifa et al. 2014; da Silva et al. 2018; Prieto-Torres et al. 2018). We suggest the creation of new protected areas in La Guajira Peninsula, the Caribbean coast near Cartagena and Tierra Bomba Island, the intra-Andean arid zones of Colombia and Venezuela, and Lara and Falcón states in Venezuela. These efforts are crucial for preserving local bat populations and protecting critical mating and maternity roosts, stopover foraging grounds, and migratory routes.
Although SDM models are valuable tools in conservation planning, they have limitations. For instance, the SDM assumes that species are at equilibrium with the environment, and biotic interactions are usually not considered due to the Eltonian Noise Hypothesis (Peterson et al. 2011). Our study did not include cave occurrences in the SDM due to the limited number of known L. curasoae roosts and the scarcity of cave inventories. Future research should extend the use of SDM to examine how food resource availability, migratory patterns, and climate change could impact L. curasoae populations.
Implications for conservation. Nectar-feeding bats like L. curasoae play a crucial role in plant pollination and seed dispersal, essential for maintaining dry forests and arid habitats (Arita and Wilson 1987). Therefore, preserving L. curasoae will contribute to maintaining biotic interactions, plant diversity, and the ecosystem services this species provides in the dry and xerophytic ecosystems of the ABC islands, northern Colombia, and Venezuela. Although representation of dry forests within Colombia’s protected areas increased by 68.4 % from 2010 to 2020, these ecosystems remain the least represented in the Colombian protected areas' system (Corzo et al. 2023). Connectivity between protected areas is also lower in the Caribbean ecoregion due to human transformation and because it has the smallest remnants of natural dry forests (Castillo et al. 2020).
Given the possible migratory behavior of L. curasoae, we recommend not only expanding existing natural areas or creating new ones but also establishing conservation corridors (Belote et al. 2016) and a network of connected private reserves across political boundaries (Kark et al. 2009). Conservation corridors should be prioritized along protected areas on the Caribbean coast of Colombia and Venezuela and in Falcón and Lara states in Venezuela. This approach is crucial to guarantee the seasonal movements and connectivity of L. curasoae populations throughout its distribution.
Conserving mating and maternity roosts is crucial since L. curasoae is strongly cave-dependent (Cole and Wilson 2006). Currently, seven important areas for bat conservation (AICOMs by its acronym in Spanish) and four important sites for bat conservation (SICOMs by its acronym in Spanish) have been recognized by the Latin American and Caribbean Network for Bat Conservation (RELCOM, https://www.relcomlatinoamerica.net/) as of high value to protect areas and caves or human buildings that serve as maternity and mating roosts of L. curasoae (Bárquez et al. 2022). Despite these efforts, our study indicates that roost vandalism remains a significant threat, menacing the species in several locations. For instance, as we completed the preparation of this article, we received a report from the coordinators of the Venezuelan Program for Bat Conservation (PCMV, the Venezuelan node of RELCOM) of a presumed case of massive poisoning of the bat colonies roosting in the Butare Tunnel at Falcón State (Venezuela), one of the identified roots used by thousands of L. curasoae bats (ID L385, Table ST1; A. García-Rawlins, Asociación Civil Topotepuy, pers. comm.). More than 200 adults and pups of L. curasoae and Mormoops megalophylla were found dead on the floor of the tunnel and hanging on the walls (date of sighting May 31st, 2024); however, it is unknown what substance was used. Roosts discovered in Colombia are particularly sensitive because one of them is the only known maternity roost for L. curasoae, and the other two are used by a colony only during mating season. Since its rediscovery in 2020, this colony has returned in 2022 and 2023 (A. Otálora-Ardila, unpublished data). Consequently, these two roosts are vital to maintaining the presumed long-distance movements of this species between the western part of its distribution and the north. Therefore, it is critical to establish effective management actions to protect bat colonies in those sites (see examples in Simal et al. 2021).
Our findings suggest that L. curasoae populations along the Caribbean coast of Colombia and Venezuela and in intra-Andean dry areas face more threats than those in the ABC islands. In the latter, ongoing bat protection initiatives have been implemented or are in the process of being established, having as targets the main bat roosts identified in the three islands (Simal et al. 2021). Identifying roosts in intra-Andean areas, determining their lapse of use, and assessing routes of migratory movements between coastal and intra-Andean zones is crucial. This is particularly important as planned wind farm developments in Colombia (UPME 2023b) could disrupt bat navigation (Jonasson et al. 2024), and interfere with these not yet identified flight routes. We also recommend creating intergovernmental action plans to protect this bat species at a regional level, addressing gaps in knowledge and guiding conservation efforts in a coordinated way to protect L. curasoae, with special focus on protection of maternal and mating roosts. Despite being included in general national-level strategies focused on migratory species (MAVDT et al. 2009), L. curasoae lacks specific conservation measures (Rojas-Díaz and Saavedra-Rodríguez 2014) and is often omitted from strategies aimed at protecting pollinator species (MADS et al. 2021).

