











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
The tribe Oryzomyini is one of the most diverse groups of rodents on the planet, with 33 genera and over 152 species (Weksler 2015; Pardiñas et al. 2017). However, it is also one of the taxa that has undergone numerous recent taxonomic changes. These proposals include the split of some genera into distinct new genera, new species, and the revival of old names (Weksler 2006; Percequillo et al. 2011; Pine et al. 2012; Brito et al. 2019; Brito et al. 2020; Caccavo and Weksler 2021; Prado et al. 2021; Semedo et al. 2021). Recent phylogenetic studies, combined with detailed morphological and morphometric analysis, have revealed previously hidden diversity of rodents within the tribe Oryzomyini. The recognized diversity is expected to continue growing, particularly in the tropical regions of South America, where these rodents have been historically understudied, but are known to be highly diverse. Within the tribe Oryzomyini, the genus Oryzomys has undergone significant reorganization in the last two decades, with morphological and molecular analyses leading to its division into at least 11 different genera (Weksler 2006; Weksler et al. 2006). Nevertheless, it remains one of the most widely distributed genera within the family, found from the southeastern United States of America to Colombia and Venezuela, including several islands (Hall 1981; Musser and Carleton 2005; Weksler 2015).
Oryzomys, after being redefined, has also undergone recent taxonomic changes, including the placement of taxonomic forms that were previously considered subspecies at the species level (Carleton and Arroyo-Cabrales 2009; Hanson et al. 2010; Indorf and Gaines 2013). Since of the revision of Musser and Carleton (2005), four species were recognized within the genus (O. couesi, O. dimidiatus, O. gorgasi, and O. palustris), later an ancient species from the island of Jamaica, O. antillarum, was confirmed (Morgan 1993; McFarlane et al. 2002; Weksler et al. 2006). Through a morphological and morphometric study, three additional species (O. nelsoni, O. peninsulae, and O. albiventer) were recognized (Carleton and Arroyo-Cabrales 2009). Subsequently, Hanson et al. (2010) conducted a molecular phylogenetic analysis of the genus in Central America and North America, leading to the separation of O. texensis and O. mexicanus. The taxonomic situation of O. argentatus is also complex and widely discussed, but current evidence indicates that it is a distinct species facing serious conservation problems (Goodyear 1991; Wang et al. 2005; Indorf and Gaines 2013). Actually, at least 11 species of the genus Oryzomys are known, with two additional species, one from Panama and other from Costa Rica, pending formal descriptions (Hanson et al. 2010).
The species Oryzomys couesi (Alston, 1877) is the most widely distributed species of the genus, found on the coastal plains of the Gulf of México from Tamaulipas in México to northern Colombia, including most of the Yucatán Peninsula, as well as on the Cozumel Island (Reid 1997; Hanson et al. 2010; Vázquez-Domínguez et al. 2020). Oryzomys mexicanus Allen, 1897 distributes in tropical regions of the coastal plains of the Mexican Pacific from Sonora to at least to El Salvador (Hanson et al. 2010; Percequillo 2015). Ramírez-Pulido et al. (2014), after formally restricting its type locality to the "Valle de Mexico", proposed that this species must be called O. fulgensThomas, 1893 because this name has priority name over O. mexicanus.
Oryzomys fulgens is an enigmatic form known from only a few specimens. The main problem is that it was described with a single specimen without a precise type locality, only as "México", and the holotype has never been compared with other specimens from México (Merriam 1901; Goldman 1918; Carleton and Arroyo-Cabrales 2009); these facts have prevented the name from being correctly assigned to any known population and as such contrasted with populations of other taxa, making it a name that practically have not being used, despite the fact that the name is one of the oldest associated to the genus. We interpreted that the authors of the past visualized the species, or in its case the name, in at least three different ways after its description. Many researchers accepted it with caution as a distinct species, awaiting future research that would delineate it geographically and morphologically (Merriam 1901; Goldman 1918; Hall 1981). Other authors consider it taxonomically dubious (Anderson 1977), assuming that it is nomenclaturally unavailable. Most of the times it was recognized as a subspecies of “O. couesi”, lacking a clearly defined geographical distribution as expected for a subspecies (Musser and Carleton 1993, 2005). Thus it was used for more than 40 years; sometimes it was used as conspecific of O. crinitus (e. g., Ramírez-Pulido et al. 2005), another subspecies of “O. couesi” that was also described from the Basin of México and therefore theoretically occupied the same distribution area.
Rice rats of the genus Oryzomys usually live in close relationship with water or flooded areas and wetlands, both in inland wetlands and in mangroves and marshes (Wolfe 1982, 1985). However, although they can be found in other environments (Genoways and Timm 2005; Engstrom et al. 1989; Poindexter et al. 2012a, b), but that periodically include semi-aquatic microhabitats (Benson and Gehlbach 1979). Therefore, we considered that the region Xochimilco region could be the last possible area in the Basin of México with the ecological conditions where a population of O. fulgens could still exist. We sample on that area and at the light of results based on the captured specimens we made taxonomic considerations on the distinction of O. fulgens.
Material and methods
The area of the Xochimilco lake is a valuable remnant wetland within one of the most highly urbanized regions in México. It is located within the Basin of México (also known as Valle de México) in the southeastern of México City. It comprises an area of 2,657 hectares. It is internationally recognized as a Zone of Historical Monuments, Protected Natural Area, a World Natural and Cultural Heritage Zone, and a RAMSAR wetland (Wigle 2010). Xochimilco comprises a complex lake system consisting of interconnected channels, permanent, and temporary lagoons. The depth of these water bodies varies, with some channels reaching 60 cm and the San Gregorio Atlapulco lagoon reaching depths of 2 to 4 m. The entire system is estimated to have an approximate length of 203 km, and currently relies on artificial feeding through treated water sources (Figure 1).
Marginal aquatic vegetation in the Xochimilco region is diverse, consisting of emerged vegetation such as tulares (Typha latifolia) and floating vegetation including water lilies (Eichornia crassipes) and lentils (Lemna minuscula and L. trisulcata) found in channels and lagoons. In non-flooded areas, there are cultivated lands, occasionally interspersed with different pastures such as Agrostis semiverticillata and Pennisetum clandestinum. Trees in the region comprises mixed forests consisting of native species like pines (Pinus sp.), cedars (Cedrus sp.), ahuehuetes (Taxodium sp.), and willows (Salix sp.). However, there are also introduced species like casuarinas, eucalyptus, and pirules (Schinus sp.) (Jiménez-Gutiérrez 2016). The Xochimilco region has a temperate sub-humid climate characterized by summer rains. The average annual rainfall is around 620 mm and the average annual temperature is 16 °C (Jiménez-Gutiérrez 2016).
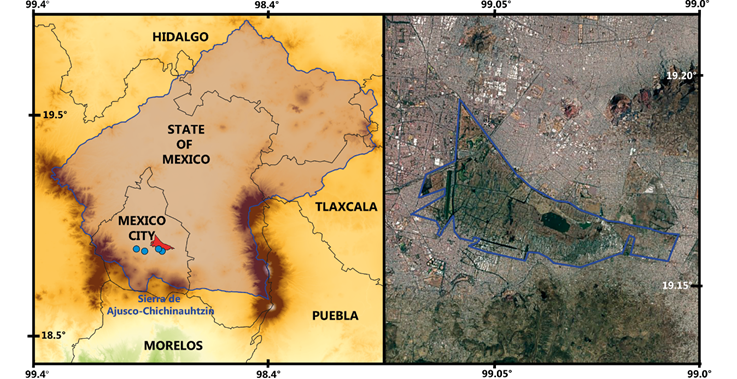
Figure 1 Region associated with the distribution of Oryzomys fulgens. Left: Limits of the Basin of Mexico with the Xochimilco region (in red). Right: Lake Xochimilco within the urban area of Mexico City. The blue dots show the localities of O. fulgens: 1) Probably the type locality “Tepepan” (Boucard 1894), 2) type locality of O. crinitus of “Tlalpan” (Merriam 1901), 3) “Xochimilco” (González-Romero 1980), and 4) 2 km N, 3.9 km W of San Gregorio Atlapulco, 2,240 m (this study).
During our study on the wild mammals that inhabit the Xochimilco Lake Zone, aimed at understanding the diversity, distribution, and abundance in the region, we conducted 12 monthly samples from February 2013 to February 2014. For rodents, we used Museum Special, Victor, and Sherman traps baited with oats. In total, 2,517 trap-nights were set across all types of plant associations.
Most of the collected rodents were released at the site of capture, while some individuals were prepared as museum specimens and deposited in the Mammal Collection of the Metropolitan Autonomous University (UAMI); no tissue sample was salved. Additionally, reference and comparison specimens, unless explicitly stated, are housed in the same collection. Collected specimens were morphologically compared with the holotypes of Oryzomys fulgens and O. couesi by two of us (NGR and JAC), which are preserved at the Natural History Museum (BMNH) in London, England.
External dimensions, in mm, were transcribed from skin tags: total length (TOTL); tail length (TL); hindfoot length (HFL); and ear length (EL). Other 17 cranial variables were measured to 0.01 mm using digital calipers, and taken according to the specifications of Carleton and Arroyo-Cabrales (2009). These measurements and their abbreviations are: total length (TOTL), tail length (TL), hindfoot length (HFL), ear (pinna) length (EL), occipitonasal length (ONL), greatest zygomatic breadth (ZB), breadth of rostrum (BR), interorbital breadth (IOB), breadth of braincase (BBC), depth of braincase (DBC), breadth of bony palate (BBP), breadth of zygomatic plate (BZP), length of diastema (LD), breadth across incisive foramina (BIF), width of the upper first molar (WM1), length of rostrum (LR), length of bony palate (LBP), postpalatal length (PPL), length of incisive foramen (LIF), coronal length of maxillary tooth row (CLM), and length of auditory bulla (LAB). All specimens examined are adults according to the criteria of Carleton and Arroyo-Cabrales (2009).
Results
With 2.517 trap-nights a total of 269 rodent specimens were collected, resulting in a capture success rate of 9.4%. Collected specimens belong to 17 species, including Reithrodontomys megalotis, Baiomys taylori, Mus musculus, Rattus sp., Microtus mexicanus, and Oryzomys fulgens. Among them, R. megalotis was the most abundant species, accounting for 54.6 % of the collected rodents, while Oryzomys fulgens was the least abundant (two specimens), comprising only 0.7 % of the specimens. Invasive species were also present in our samples, with Mus musculus representing 8.9 % and Rattus sp. accounting for 6.7 % of the collected rodents. It is worth noting that despite the genus Peromyscus being one of the most diverse and abundant in North America (Carleton 1989), no species from this genus was trapping sites during this study. However, it is important to mention that two species of the genus (P. gratus and P. melanotis) were previously recorded in the study area more than 40 years ago (González-Romero 1980).
In the locality of 2 km north and 3.9 km west of San Gregorio Atlapulco (19.2758° N, -99.0940° W), we collected two female specimens of Oryzomys fulgens. One specimen was preserved as a skin and skull and deposited under number UAMI 17346, while the other specimen was released at the collection site (Figure 2C, D). Both individuals were captured in the canals surrounding a chinampa in February 2014. The collection site was situated on the banks of a canal where there was a high abundance of tulares (Typha latifolia; Figure 2B). These areas also contained other types of vegetation, including crops. Land cover of this locality also exhibited an abundance of grass (Pennisetum clandestinum), while the tree layer consisted of ahuehuetes (Taxodium sp.), along with some eucalyptus and pirules. Additionally, Baiomys taylori, Reithrodontomys megalotis, and Rattus sp. were found in the same area. Oryzomys fulgens was the only rodent species exclusively found within the tulares, but nearby other species are found, primarily R. megalotis.

Figure 2 Photographs of the Xochimilco region and specimens of Oryzomys fulgens. A) General view of the Xochimico wetland. B) Precise site where we collected the two specimens of O. fulgens. C) A female specimen (UAMI 17346) and D) A second female specimen (released at the collection site) of O. fulgens.
Discussion
Taxonomy. Oryzomys fulgens was originally described based on a single male specimen (Holotype: British Museum Natural History, BMNH 70.6.20.3) collected by Adolphe Boucard in México. The precise locality of the specimen is unknown, as it was only recorded as "Mexico" (Thomas 1893a). Adolphe Boucard, a naturalist, along with Auguste Sallé, collected several species of amphibians, reptiles, invertebrates, and particularly birds in central and southern México between 1865 and 1867 (Flores-Villela et al. 2004). Many of these specimens did not have specific localities documented (see Papavero and Ibanez-Bernal 2001). Thomas (1893b) suggested that the type specimen of O. fulgens was likely collected alongside the holotype of the gopher Geomys merriami [Cratogeomys merriami], which is primarily found in the Basin of México. This assumption prompted Merriam (1901) and later Goldman (1918) to designate the type locality as "southern Mexico, probably in or near the Valley of Mexico [= Basin of México]." Finally, Ramírez-Pulido et al. (2014) provided additional evidence and formally restricted the type locality to the "Valle de México" (Gardner and Ramírez-Pulido 2020). Additional evidence supporting that the type specimen of O. fulgens was collected within the Basin of México is that A. Boucard, during his extensive travels throughout México, journeyed from Acapulco to México City (Boucard 1894:72). Within the Basin of México, he passed through [San Miguel] Topilejo, which is approximately 19 km from the collection location of our O. fulgens specimen from Xochimilco. He also reached Tepepa[n], which is 9.7 km from our collection site. It is plausible that it was in Tepepan where he collected both the type specimens of Cratogeomys merriami and O. fulgens, as in the past, it likely had suitable conditions for both species. However, presently, Tepepan is integrated into the urban area of México City. Defining the type locality was crucial because O. fulgens, after O. couesi and O. palustris, is the third oldest name among all species and subspecies associated to the genus Oryzomys, and then is of much relevance to name any lineage of Oryzomys present in México.
We had the opportunity to compare the type of O. fulgens with the specimen collected at Xochimilco. Although the holotype skull is missing most of braincase and the zygoma is broken, they are very similar, except that the type has the interorbital region not completely convergent anteriorly and the frontal extension of the zygomatic plate is more pronounced (Figure 3). Some of these differences and, even more, the similarities, had already been noticed by Merriam (1901) when interpreting the O. fulgens description and comparing it with the holotype of O. palustris crinitus. In fact, this author mentioned that they are “strongly marked species appears to be closely related” (see also Goldman 1918). Our specimen is female and the type of O. fulgens is male, which is relevant given that the species of this group present sexual dimorphism (Carleton and Arroyo-Cabrales 2009). In addition, even when the type misses its posterior part of the skull, we can interpret that the two specimens are very similar and likely belong to the same species. We also compared the skulls of O. fulgens with those of O. albiventer and O. mexicanus, the other species southern and western México to understand and morphologically define the characteristics of the species of the genus Oryzomys in México. In the comparison of O. fulgens with O. mexicanus we observed that the former has the previous enterostyle of the first upper molar; the upper molars are noticeably larger and more robust, particularly the anterocone is larger (Figure 4); the posterior edge of the incisive foramen reaches the posterior edge of the upper first molars (in most specimens of O. mexicanus it does not reach that edge); the lateral extension of the lacrimal bone forms only a small notch whereas in O. mexicanus it is well developed; the interorbital region convergent anteriorly but with a contraction in the proximal part, in O. mexicanus this contraction regularly does not occur (Figure 3); the posterior exposure of the parietal is large and square while in O. mexicanus it is small and triangular. In addition, the hair is long, it has a fine, dense and silky coat, while in O. mexicanus it is short and harsh. In O. fulgens, unlike O. albiventer, the mesopterygoid fossa extends forward before the middle of the pterygoid fossa, while in albiventer it extends beyond the middle of the pterygoid fossa; the posterior part of the rounded occipital and nape extends behind the condyles of the foramen magnum, in O. albiventer it is flat and extends further behind the condyles (Figure 1); the interparental bone thick and elongated versus small and narrow. O. albiventer is notably larger in overall skull and body size (Table 1; see also Carleton and Arroyo-Cabrales 2009). The dorsal hair of O. fulgens is long and silky like that of O. albiventer, possibly as an adaptation of the two species to temperate regions at higher elevations (see below; Figure 3).
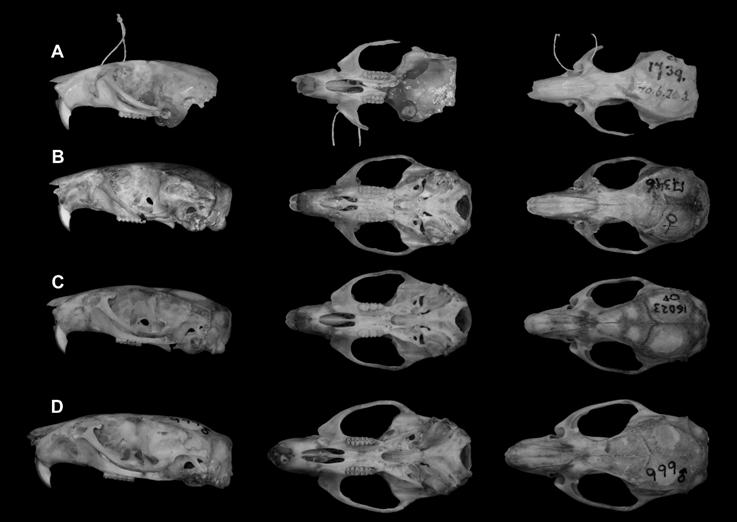
Figure 3 Lateral, ventral and dorsal view of some of the specimens examined. A) Holotype, a young adult male, of Oryzomys fulgens (BMNH 70.6.20.3). B) Young adult female O. fulgens collected in Xochimilco at 2 km N, 3.9 km W San Gregorio Atlapulco, 2240 m (UAMI 17346). C) Old adult male O. mexicanus from El Salto, 4 km WSW Minatitlán, Colima (UAMI 16023). D) Young adult male O. albiventer from 3 km W Venustiano Carranza, Michoacán (UAMI 999).
Craniodental measurements are also similar between the holotype of O. fulgens and the Xochimilco specimen, although the latter is slightly larger in the breadth of zygomatic plate and length of bony palate (Table 1), but these small differences could be interpreted as sexual dimorphism. However, the two O. fulgens specimens are consistently larger in all variables than the average O. mexicanus specimen, and both species are markedly smaller than O. albiventer (Table 1).
Due to the morphological differences of O. fulgens, and its inhabitant of highlands and temperate climate that strongly contrasts with the habitats of other species of the genus Oryzomys, we consider that O. fulgens represents, as originally proposed by Thomas (1893a), a distinct species of the genus. We agree with the taxonomic proposal of Ramírez-Pulido et al. (2014) but with the difference that we consider that O. fulgens that restrict to the Basin of México. Therefore, the populations of the Pacific coastal plains, distributed from Sonora to El Salvador, correspond to O. mexicanusMerriam, 1901 as it is the oldest name in the region (see also Carleton and Arroyo-Cabrales 2009; Hanson et al. 2010).
Our taxonomic hypothesis should be further tested with the study of additional specimens and the analysis of genetic variation. The later would also allow testing the phylogenetic position of O. fulgens.
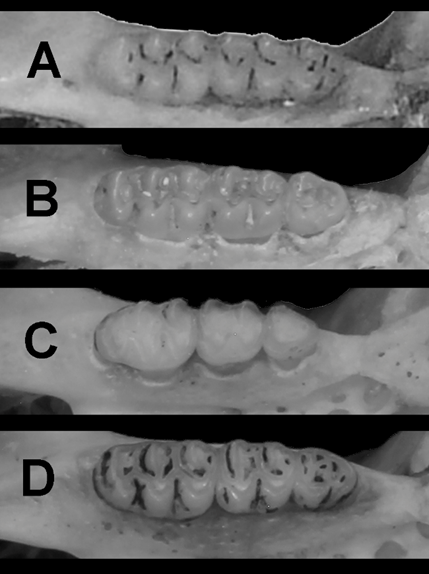
Figure 4 Occlusal views of right upper molar of some of the specimens examined. The specimens are the same as in figure 2.
Distribution and habitat. The first locality or region where O. fulgens was recognized is the "Valle de México," collected between 1865 and 1867 (Boucard 1894; Ramírez-Pulido et al. 2014; Gardner and Ramírez-Pulido 2020). The second locality of the species corresponds to the type locality of O. palustris crinitus "Tlalpam [Tlalpan], Federal District [Ciudad de México], Mexico," collected by E. W. Nelson and E. A. Goldman in 1892 (Merriam 1901; Goldman 1918). The third location was almost 90 years later, González-Romero (1980), in his work on rodent pest from México City, mentions O. couesi crinitus from "Xochimilco" without a precise location. However, a one specimen of the rice rat was collected within the chinampera zone of Xochimilco, but it was not preserved in a scientific collection (A. Gonzalez-Romero, pers. comm. 2019); however, two photographs were obtained of this specimen that were later published in other works (Ceballos and Galindo 1984; López-Medellín and Medellín 2005). It is necessary to clarify that Ceballos and Galindo (1984) included in the report of O. palustris crinitus a specimen of "Xochimilco (UAMI)"; however, no such specimen is cataloged in the Mammal Collection of the Autonomous Metropolitan University (UAMI).
Table 1 External and craniodental measurements (in mm) of some specimens of Oryzomys from south-central México. The measurements correspond to holotype of Oryzomys fulgens (probably the type locality “Tepepan”, BMNH 70.6.20.3), specimen of O. fulgens collected in Xochimilco (2 km N, 3.9 km W San Gregorio Atlapulco, UAMI 17346), specimens of O. mexicanus (Michoacán: Las Campanas 6 km N Maruata, UAMI 16025; 2 km NW Presa Pucuato, UAMI 16026; El Capiro 20 km SSW Nueva Italia, UAMI 16027. Colima: El Salto 4 km WSW Minatitlán, UAMI 16023), and specimen of O. albiventer (Michoacán: 3 km W Venustiano Carranza, UAMI 999). See materials and methods for variable abbreviations.
| Variable | O. fulgens (holotype) | O. fulgens (this work) | O. mexicanus | O. albiventer |
|---|---|---|---|---|
| TOTL | - | 262.0 | 280.3 | 344.0 |
| (260.0-344.0) | ||||
| TL | - | 139.0 | 148.7 | 181.0 |
| (142.0-181.0) | ||||
| HFL | - | 36.0 | 31.7 | 39.0 |
| (30.0-39.0) | ||||
| EL | - | 18.0 | 16.7 | 19.0 |
| (15.0-20.0) | ||||
| ONL | - | 32.7 | 30.8 | 36.5 |
| (29.6-36.5) | ||||
| ZB | - | 17.7 | 16.5 | 19.9 |
| (15.7-19.9) | ||||
| BR | 6.6 | 6.6 | 5.9 | 7.4 |
| (5.6-7.4) | ||||
| IOB | 4.8 | 5.0 | 4.9 | 5.3 |
| (4.7-5.3) | ||||
| BBC | - | 13.8 | 13 | 14.3 |
| (12.6-14.3) | ||||
| DBC | - | 11.0 | 10.3 | 11.4 |
| (9.4-11.4) | ||||
| BBP | 5.9 | 6.2 | 5.7 | 6.7 |
| (5.5-6.7) | ||||
| BZP | 3.7 | 4.4 | 3.5 | 4.0 |
| (3.2-4.1) | ||||
| LD | 8.9 | 8.5 | 7.8 | 9.5 |
| (7.2-9.5) | ||||
| BIF | 2.2 | 2.5 | 2.9 | 3.5 |
| (2.3-3.5) | ||||
| WM1 | 1.5 | 1.4 | 1.4 | 2.0 |
| (1.4-2.0) | ||||
| LR | 10.2 | 11.1 | 10.2 | 12.4 |
| (9.9-12.4) | ||||
| LBP | - | 6.6 | 6.1 | 6.8 |
| (5.9-6.8) | ||||
| PPL | - | 11.2 | 10.6 | 20.7 |
| (10.1-20.7) | ||||
| LIF | 7.0 | 6.7 | 6 | 7.1 |
| (5.5-7.1) | ||||
| CLM | 5.4 | 5.3 | 4.7 | 5.5 |
| (4.6-5.5) | ||||
| LAB | - | 7.0 | 5.6 | 6.3 |
| (5.46.3) |
Until now, O. fulgens is recorded at three localities: the type locality of O. fulgens is in the Basin of México, the type locality of O. palustris crinitus in Tlalpan, and our record of O. fulgens from 2 km N, 3.9 km W of San Gregorio Atlapulco, 2,240 m. Based on the known localities of O. fulgens, its distribution can be extended to México City, or more likely, to the broader Basin of México (Figure 1). Davis (1944) and later Hall and Kelson (1959), recorded a locality from Tlaxcala, "8 km SW Tlaxcala, 7,500 ft." (see also Hall 1981; Fernández et al. 2015), which occurs outside the Basin of México. Although we did not have the opportunity to review the specimen from Tlaxcala, it is probable that it corresponds to O. fulgens for four reasons: 1) the Sierra Nevada, which separates the populations of the Basin of México from those of Tlaxcala, is formed by many volcanoes with an average altitude of 3,479 m (Tlaloc, Telapón, Iztaccihuatl, Popocatépetl, among others). The oldest of those volcanoes formed approximately 1.8 million years ago, and the most recent ones formed around 0.33 million years ago (Macías et al. 2012; Siebe et al. 2017; Martínez-Abarca 2023). This suggests that the population of Tlaxcala likely separated recently from that of the Basin of México before the formation of the Sierra Nevada. 2) Additionally, the locality of Tlaxcala shares similarities in terms of climate, vegetation, and altitude with those from the Basin of Mexico, 3) furthermore it was captured under very similar conditions to our example, at the edge of a swamp (Davis 1944). 4) Moreover, it also exhibits large molars (Davis 1944), which are one of the diagnostic characteristics of O. fulgens. However, further studies are need to evaluate the taxonomic status of the Tlaxcala populations.
The genus Oryzomys generally distributed in tropical regions from evergreen tropical forests, deciduous tropical forest, lowland forests, coastal vegetation, mountain cloud forests to and wetlands in the north of the distribution, but never inhabits temperate climates and forests (Sánchez-Cordero and Valadez 1989), although they only occasionally reach mesic habitat in the case of O. albiventer (Carleton and Arroyo-Cabrales 2009). In this sense, O. fulgens completely apart from the genus pattern, it is the only species that inhabits temperate climates but is associated with halophilous and semi-aquatic vegetation. It is the species of Oryzomys that inhabits at the highest elevation between 2,192 to 2,267 m; the species that follows it is Oryzomys albiventer (1,249 to 1,760 m) whole all other inhabit at altitudes lower than 1,380 m (Figure 5) where tropical regions are normally associated.
The distribution of O. fulgens is confined to the Basin of México, with its populations isolated from the species of the O. couesi group, especially O. mexicanus and O. albiventer, to which O. fulgens may be related. The Basin of México is surrounded by volcanoes; on the western side, there is the Sierra de la Cruces with several volcanoes (Volcán La Bufa, Catedral, Iturbide, Chimalpa, Salazar, San Miguel), on the east by the Sierra Nevada (Tlaloc, Telapón, Papayo, Iztaccihuatl, and Popocatépetl volcanoes), to the north, the Sierra de Guadalupe, and to the south, the Sierra de Ajusco-Chichinauhtzin (Wallace and Carmichael 1999). The Sierra de Ajusco-Chichinauhtzin is of particular interest because it geographically isolates O. fulgens from the southern populations of O. mexicanus in the nearby states of Morelos, southern Puebla, and Guerrero (Hall 1981). It is a mountainous system of Sierra de Ajusco-Chichinauhtzin that limits the southern part of the Basin of México, formed by more than one hundred volcanoes, including Chichinauhtzin, Tláloc, Xitle, Cerro Pelado, Cuauhtzin, and with an average height of 3,400 m (Figure 1; Bloomfield 1975; Vázquez-Sánchez et al. 1989). This Sierra was formed between 700,000 and 400,000 years ago (Bloomfield 1975; Márquez et al. 1999; Vázquez-Sánchez et al. 1989). As it developed, it physically and ecologically impeded faunal exchange towards the southern tropical regions, thus resulting in the isolation and refuge of O. fulgens in a temperate region with altitudes >2,200 m within the Basin of México. As said, this scenario should be tested with formal historical biogeographic analyses.
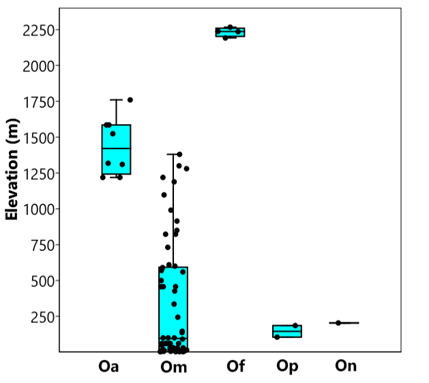
Figure 5 Variation in the elevational distribution of species of Oryzomys in southern and western México. Oa = Oryzomys albiventer, Om = O. mexicanus, Of = O. fulgens, Op = O. peninsulae, On = O. nelsoni. Most altitudinal records were obtained from Carleton and Arroyo-Cabrales (2009).
Conservation. The abundance of O. fulgens, measured by the number of specimens collected over the past 120 years or more, is remarkably low, despite México City and the Basin of México being extensively studied historically (Villa-Ramírez 1953; Ceballos and Galindo 1984; Hortelano-Moncada and Cervantes 2011), only three individuals are known to exist. Species that are rare, localized, and restricted to specific ranges are particularly vulnerable to demographic and environmental changes, such as reduced reproductive capacity and increased susceptibility to catastrophic events like floods and fires (Kunin and Gaston 1993; Gaston and Gaston 1994; Flather and Sieg 2007).
The Xochimilco region is the only known habitat where O. fulgens likely persists. This area is entirely surrounded by human densely populated regions (Figure 1), making it highly susceptible to human pressures, especially concerning resource exploitation, construction activities within the region, conversion of chinampas areas for tourism, recreation, and sports facilities, among other factors. Although further studies might reveal additional populations within the Basin of México, they would likely face similar pressures due to their association with México City and other large cities within the Basin of México.
Predators pose a significant threat to the rice rat population. Invasive predators, particularly cats and dogs, have been highly detrimental to global biodiversity (Doherty et al. 2016). Within the collection points of O. fulgens, several predator species, including domestic or feral dogs (Canis lupus domesticus), domestic or feral cats (Felis catus), rattlesnakes (Crotalus polystictus), and other types of snakes are frequently encountered. Although we do not currently have direct evidence of these predators preying on O. fulgens, their presence in abundance at the site could contribute to the extinction risk faced by the rice rat.
Invasive rodent species, notably the black rat (R. rattus) and Norway rat (R. norvegicus), are well-known for their association with the extinction or decline of native vertebrate populations, including reptiles, birds, and mammals, particularly other rodents (Harris 2009; Stokes et al. 2009; Harris and Macdonald 2007). In the same area where O. fulgens was collected, Rattus sp. were also frequently captured (capture success rate >6.7%), suggesting possible interactions between the two species, potentially due to overlapping spatial and habitat usage (Stokes et al. 2009). The abundance of the invasive black rat may intensify competition between it and the native rice rat, as observed in Florida, and has been implicated in the extinction of several Oryzomys species on various islands, including the Galapagos Islands and the Antilles (Goodyear 1992). Despite extensive evidence of rodent extinction due to the black rat on oceanic islands (Álvarez-Castañeda and Cortés-Calva 1996; Álvarez-Castañeda and Ortega-Rubio 2003; Harris 2009), the Xochimilco region can be considered an urban island, completely surrounded by urban areas, making the impact of the black rat on O. fulgens potentially similar to what occurs on oceanic islands.
The conservation problems of O. fulgens and its probable risk of extinction are summarized due to a variety of factors, including low population abundance, a restricted area of distribution, urbanization, and habitat destruction, as well as the impact of introduced predators and invasive rodents.
Xochimilco and conservation of Oryzomys fulgens. In past, the Basin of México featured a landscape dominated by five large lakes, which would merge during the rainy season into one continuous water body, all surrounded by imposing mountains that impeded water drainage (Candiani 2014; Manley et al. 2022). The major lakes in the area were Lake México-Texcoco, Lake Xochimilco, and Lake Chalco, with Zumpango and Xaltocan being smaller in size. Until pre-Hispanic times, the region surrounding these lakes likely provided favorable conditions for the survival of O. fulgens, which might have been distributed in association with the lakes and their tributaries, spanning from the southernmost in Xochimilco to the northernmost in Zumpango.
This five-lake water system has undergone countless anthropogenic alterations over two thousand years (Narchi 2014; Manley et al. 2022). Humans have occupied the Basin of México in the last 20,000 years BC, and the population grew significantly with the rise of Tenochtitlan and even more with the arrival of the Spanish from the 13th to the 17th centuries. Later, there was accelerated growth in the middle of the 20th century (Narchi 2014; Voss et al. 2015), until it became the fifth largest urban area in the world with more than 22 million people, concentrating a large part of México's industrial, commercial, and political activity (Manley et al. 2022; UN 2018). This rapid urban growth has multiple consequences at different scales, including: 1) Reduction of the original vegetation cover area, substituted by urban areas or cultivated fields (Ezcurra et al. 1999; Merlín-Uribe et al. 2013). 2) Alteration of precipitation patterns, changes in groundwater levels and drying (Bojorquez Tapia et al. 1998; Tortajada and Castelán 2003), and 3) Decrease in biodiversity (Rodríguez-Franco 2001; Ezcurra et al. 2001; Narchi 2014). Currently, some lakes no longer exist, particularly the remnants of Lake Zumpango, Texcoco, and Chalco, surrounded by urban, peri-urban areas and, in the least of cases, cultivated fields, so their surroundings do not seem to be the appropriate habitats for O. fulgens. However, Oryzomys are not of the lake itself, is more in relation to the wet areas and the marsh areas associated to these lagoons. Lake Xochimilco is a remnant water system of the Basin of México lakes composed of channels, small lakes, marsh areas, and temporary wetlands surrounded by urban regions (Pérez-Belmont et al. 2021; Merlín-Uribe et al. 2013a). Most of Xochimilco is made up of small artificial islets that ancestrally were built as farm sites called chinampas, which have the optimal condition for the species of Oryzomys.
The chinampas have been cultivated for at least six centuries and reached their maximum development between 1400 and 1600 (Merlín-Uribe et al. 2013b). This ancestral agricultural technique has been used since the Mexicas and is currently considered one of the most sustainable and productive production methods in the world (Rojas 1983; Alcántara 2020; Pérez-Belmont et al. 2021), with the addition that this system preserves great parts of the original vegetation on all three strata and the marginal subaquatic vegetation. This network of canals and vegetation produces a landscape that probably positive directly impacts the quality and availability of habitat for the rice rat O. fulgens. We believe that the ecological characteristics of the chinampas have maintained, although with low abundance, the populations of O. fulgens and promote its possibilities for study and conservation, unlike other Oryzomys species that have become extinct under similar pressures. However, even though the chinampas area is an ideal habitat for the rice rat, it is exposed to anthropogenic factors that can severely decimate their populations (see above). The anthropic effect of Xochimilco have a dual effect in the O. fulgens. On one hand the chinampas, use, build, and conservation increase the optimal habitat for the species, for which need to be abundant, but in the other hand this same activity increase the exotic species that can cause the extinction of the species (e.g., feral cats and dogs; Doherty et al. 2016). Unfortunately, at present this area of Xochimilco is constantly subjected to anthropogenic pressures that endanger the native vegetation where these rodent live, so its long-term permanence is uncertain. The rarity and limited population data for these species make them particularly vulnerable to threats, such as habitat loss, fragmentation, and invasive and exotic species. Conservation efforts are crucial to protect these unique rodents and their habitats and prevent further decline in their populations.

