











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
For large herbivores living in dry environments, water can be a limiting resource that affects their distribution and abundance in periods when water requirements are not met through forage (Villarreal-Espino and Marín 2005; Nagy and Gruchacz 1994; Eliades et al. 2022). In these circumstances, many animals depend on access to surface freshwater sources, especially during certain critical periods of the year (Moro-Ríos et al. 2008). Water availability is one of the factors that influence the spatial distribution of wildlife to different degrees (Paredes et al. 2017); in some cases, it causes changes in the behavior of animals (Pacifici et al. 2015). In habitats where water is a limiting resource, animals usually gather around available water sources (Redfern et al. 2003).
Worldwide, the implementation of artificial water troughs for wildlife has been reported as a successful strategy to mitigate the consequences of water shortages in drought periods (Epaphras et al. 2008). The implementation of these artificial water sources includes various strategies; for example, in some cases, it is sought to concentrate the populations of wild fauna within selected areas, such as protected natural areas (PNA; Sutherland et al. 2018), thus preventing the animals from entering inhospitable habitats outside the PNA where they could die. On the other hand, water troughs have been conceived as a strategy that can contribute to the maintenance of wildlife populations in the short term by allowing access to water during the drought season (Mandujano-Rodríguez and Hernández 2019; Borges-Zapata et al. 2020).
To counteract the effects of prolonged drought in some areas of México, artificial water sources such as jagueyes and water troughs have been used (Villarreal 2006), which have contributed to habitat enrichment and the maintenance of wildlife populations (Bello et al. 2004). Particularly, the implementation of water troughs for wildlife in México has been reported to be a successful strategy to mitigate the consequences of water shortages in drought periods, mainly in places where water is a limiting resource (Mandujano-Rodríguez and Hernández 2019; Borges-Zapata et al. 2020).
In recent years, increasingly more extreme temperatures have been recorded in the Yucatán Peninsula, with prolonged events of high temperatures (Mardero et al. 2020), forcing wildlife to resort to water sources for thermoregulation, as observed in semiarid areas (Bello et al. 2004). This trend has affected Calakmul in southeastern México, a region that lacks large rivers or surface water bodies (García-Gil et al. 2002). This is important for ungulates inhabiting the Maya Forest (a forest area in the states of Campeche, Chiapas, and Quintana Roo, México), as habitat selection by these mammals has been reported to be strongly influenced by water availability (Pérez-Cortéz et al. 2012; Reyna-Hurtado et al. 2019). This close relationship could intensify in the coming years, as an imbalance in precipitation patterns has been recorded throughout the region (Mardero et al. 2020), reducing the availability of surface water (Reyna-Hurtado et al. 2022) and could lead to changes in population dynamics and the behavior of species in the Maya Forest region (Contreras-Moreno and Torres-Ventura 2018).
In the particular case of deer, the water-trough strategy has been used for decades in northern México in Wildlife Management Units (WMU), especially in semiarid and xeric scrub sites (Villarreal 2006; Mandujano-Rodríguez and Hernández 2019). Since 2018, water supply in artificial troughs has been conceived as a strategy that contributes to the maintenance of wildlife populations in the short term by facilitating access to water during the drought season (Borges-Zapata et al. 2020; Contreras-Moreno et al. 2019b; 2020; Delgado-Martínez et al. 2021; Pérez-Flores et al. 2021).
Identifying the functionality of artificial water troughs in improving the habitats of deer has become a research priority in México. In this regard, studies in the Maya Forest have recorded that the movements of deer and the size of their home range change during the drought season (Contreras-Moreno et al. 2019a, 2021a). Knowing the functionality of water availability for deer in artificial troughs would help improve conservation and management programs for the various deer species (Fulbrigth and Ortega-S. 2007). The objective of this study was to estimate the use of water supplied in artificial troughs by three deer species (Odocoileus virginianus, Mazama pandora, and Mazama temama) in the Maya Forest region of Campeche, in southeastern México.
Materials and methods
Study Area. The Calakmul Biosphere Reserve (CBR, in Spanish) is located in the Yucatan Peninsula, in southeast Campeche (Figure 1). It is part of the Great Calakmul Region, which includes the Maya Biosphere Reserve in Guatemala and the Rio Bravo Dos Milpas Conservation Area in Belize. It stretches across 728,908 ha (Reyna-Hurtado et al. 2022). The CBR has a warm and subhumid climate (Aw) with a mean annual temperature of 24.6 °C; the maximum height above sea level is 390 m on Mount Champerico, and the minimum altitude ranges from 100 to 150 m. The dominant vegetation types are medium semi-evergreen forests, medium subdeciduous forests, and low subdeciduous forests (Martínez and Galindo 2002; Martínez-Ku et al. 2008).
Installation of Water Troughs. As part of the efforts of CBR in collaboration with the Global Environmental Facility (GEF) Species at Risk project and the World Wildlife Fund (WWF México), artificial water troughs have been installed in the region since 2018 to counteract the effects of climate change in the region. Approximately 70 water troughs were installed, mainly within the CBR core areas. The artificial water troughs installed in the CBR are black plastic structures (Rotoplast®) with a capacity of 300 liters each. These troughs were distributed along the access road to the CBR, separated by a minimum distance of 2 km between them. At the beginning of the drought season, water was generally supplied twice monthly (every 15 days); however, as the drought season progressed and became dryer, water could be supplied once per week.
Data Recording with Camera Traps. Photographic recording of mammals with camera traps is a widely used and efficient method in southeastern México (Hidalgo-Mihart et al. 2017), particularly in the Calakmul region (Borges-Zapata et al. 2020; Contreras-Moreno et al. 2019b; 2020, 2021; Delgado-Martínez et al. 2021).
Between December 2018 and August 2021, eight Cudddeback camera traps (Non Typical Inc., de Pere, WI, USA) were installed in eight artificial water troughs placed throughout the study area (Figure 1) to record the presence of deer; the data from these eight cameras were used to perform all analyses. The study area comprised areas adjacent to the road leading to the Calakmul archeological zone (from km 20 to 47). The exact location of the water troughs is shown in Figure 1.
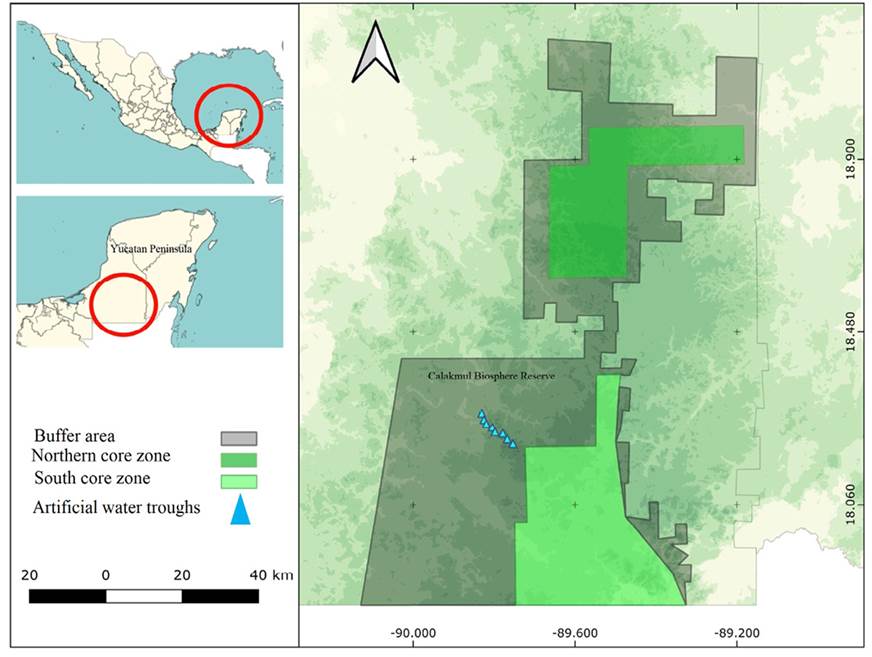
Figure 1 Map showing the location of water troughs monitored in the Calakmul Biosphere Reserve, México, where the study was carried out.
The sites where camera traps were placed were the same sites where water troughs were installed (Table 1). One camera trap was placed at each site. The number of days that each trap remained operating varied depending on the characteristics of each device; camera traps were in operation for 143 days minimum. Camera traps were installed 50 cm from the ground in trees adjacent to the water trough; they were set to capture photographs 24 hours a day, with 5-second intervals between captures. A minimum distance of 2 km was maintained between the cameras (same as for water troughs). Operating cameras were reviewed every three weeks. In each review, the photographs were downloaded and assigned a record code number; finally, the proper operation of the cameras was checked, and the batteries were replaced as needed (Hidalgo-Mihart et al. 2017). Species were identified manually, entering the following data into a spreadsheet: station, camera-trap name, date, time, species, image name, and number of individuals. After the photographs were captured, a database was built using the CamtrapR package in R 3.4.0 (R Core Team 2017; Niedballa et al. 2019).
Data Analysis. The Photograph Visit Index (PVI) was captured with the formula IFV=C/EM*100 trap-days, where C = number of captures or independent events photographed, EM = sampling effort (number of camera traps per monitoring day) and 100 trap days (standard unit; Hernández-Pérez et al. 2020). To avoid overestimating the number of recorded individuals, only independent records were considered for the analyses (Nichols and Karanth 2011).
Independent records were defined as consecutive photographs of individuals of different species and photographs of individuals of the same species captured with a separation of more than 24 hours. If more than one individual was recorded in an independent record, this was noted as one record. In photographs in which several individuals of the same or different species appeared, these cases were also considered single records. Kruskall-Wallis tests were performed to compare the visit rates of a species to each monitoring station and to compare each sampling year, and a Mann-Whitney test (Hernández-Pérez et al. 2020) to compare the independent records of the three years between species. These analyses were performed in R 3.4.0 (R Core Team 2017).
Activity patterns. The activity pattern in the study area was evaluated for two deer species (O. virginianus and M. pandora) with circular statistics to evaluate differences between visiting times. During the different sampling months, a Watson-Williams test was performed with the Oriana 4.0 software (Kovach Computing Service 2011). The degree of overlap in the activity patterns of deer species was assessed using a Kernel density analysis, and the statistical differences between the hours of high activity between species were evaluated with a Wald test supported by the packages Activity version 3.5.1 and overlap in R 3.4.0 (Ridout and Linkie 2009; Rowcliffe 2016).
Results
With a sampling effort of 4,672 trap/nights, we captured 477 separate records of O. virginianus, 229 of M. pandora, and 3 of M. temama using artificial water troughs in the CBR (Table 1). Due to the scarce records of M. temama in water troughs, comparative analyses were performed only with the other two deer species. Regarding the records for O. virginianus, highly significant differences were found between the three sampling years (H = 12.575, df = 2, P < 0.001; Figure 2a); similarly, M. pandora showed highly significant differences between sampling years (H = 9.29, df = 2, P = 0.001; Figure 2b). When the records for O virginianus were compared between sampling stations, no significant statistical differences were observed (H = 4.73, df = 7, P = 0.69); this same result was observed for M. pandora (H = 4.22, df = 7, P = 0.75). Similarly, no significant differences were found when comparing independent records of the three monitoring years between both species (W = 229, P = 0.17; Table 1).
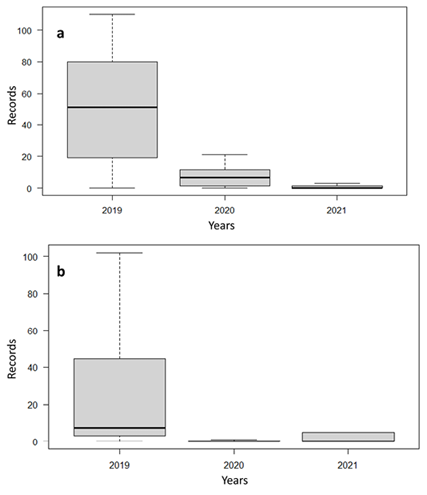
Figure 2 Independent records of O. virginianus (2a) and M. pandora (2b) during the three sampling years in artificial water troughs in the CBR.
Table 1 Independent records of deer and photographic visit index (PVI) for the three sampling years in the Maya Forest, México.
| 2019 | 2020 | 2021 | TOTAL | |||||
|---|---|---|---|---|---|---|---|---|
| Records | PVI | Records | PVI | Records | PVI | Records | PVI | |
| Odocoileus virginianus | 411 | 145.13 | 60 | 44.15 | 6 | 12.47 | 477 | 102.10 |
| Mazama pandora | 212 | 74.86 | 2 | 1.47 | 15 | 31.19 | 229 | 49.02 |
| Mazama temama | 0 | 0 | 3 | 2.21 | 0 | 0 | 3 | 0.64 |
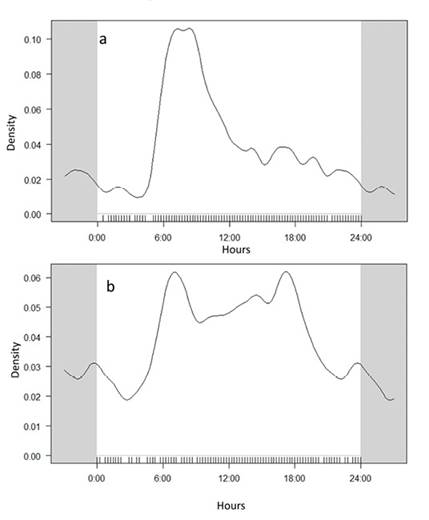
Activity patterns. For O. virginianus, the records showed a circular mean obtained at 8:15 h. and an angular dispersion ranging from approximately 8:00 h to 9:00 h. It was found that O. virginianus is mainly diurnal, with activity peaks in the early hours of the day, contrasting with a lower number of nighttime records (Figure 3a). M. pandora was also found to be mainly diurnal, showing activity peaks in the early hours of the day and before dusk (Figure 3). The circular mean of M. pandora activity was recorded at 6:00 h, with an angular dispersion ranging from approximately 6:00 h to 18:00 h. (Figure 3b). In 2019, a high degree of overlap was recorded in the use of water troughs by O. virginianus and M. pandora (IP = 0.79, P= 0,023; Figure 4); a similar pattern was observed in 2021 (IP = 0.75, P = 0,037; Figure 5).
Discussion
Two deer species, O. virginianus and M. pandora, regularly use water troughs, as shown by the number of records obtained (Table 1). The PVI showed differences between years for O. virginianus and M. pandora; the number of records in 2019 was significantly higher than in 2020 and 2021 (Table 1). In this regard, some analyses showed a trend of lower precipitation prior to 2019 (Pérez-Flores et al. 2021); particularly, 2019 showed drought precipitation conditions in terms of intensity and duration (SMN 2019). The greater presence of deer in water troughs in 2019 responds to their need for thermoregulation through water (Bello et al. 2004), so deer visited water troughs more frequently to meet their water needs (Fuller et al. 2014). Similarly, 2019 recorded a noticeable increase in the occurrence of tapirs outside the forest (Contreras-Moreno 2020; Pérez-Flores et al. 2021).
In arid regions, water is a limiting resource for various vertebrate species (McKee et al. 2015). In this study, the average PVI for O. virginianus and M. pandora for the three years was higher than the values reported in natural water holes (holes in rocky soil produced by erosion and that accumulate rainwater; Delgado-Martínez et al. 2018). Similarly, when the findings in the present study are compared with the records of Moreira-Ramírez et al. (2019), O. virginianus presented higher PVI values here compared to those recorded in Nuevo Becal (11) and the CBR (32.1), while M. pandora showed higher values only in two years (2019 and 2021); in contrast, M. temama showed lower values in the present study compared to Nuevo Becal. In this study, O. virginianus individuals were observed regularly in water troughs, even exceeding the frequency recorded for natural water bodies. This species has been considered a generalist that is able to benefit from disturbed habitats (Gallina-Tessaro et al. 2019), and on multiple occasions has displayed a positive behavior toward artificial water supply (Villarreal 2006). M. pandora is an ungulate that is considered an opportunist in the Calakmul region because it is able to use the habitat according to its availability. In this regard, an opposite trend was observed in 2020, probably due to the high availability of water in natural water bodies that year (Hernández-Cerda et al. 2021). Regarding M. temama, the values recorded in the present study (Table 1) were lower than those recorded in Nuevo Becal but were similar for Calakmul, since records were scarce or null in both cases. In this regard, it has been suggested that M. temama is more abundant in wet forests within the Maya Forest, such as Nuevo Becal, which contrasts with the characteristics of Calakmul (Moreira-Ramirez et al. 2019), where subdeciduous (dry) forests predominate toward the central part of the CBR (Martínez and Galindo 2002).
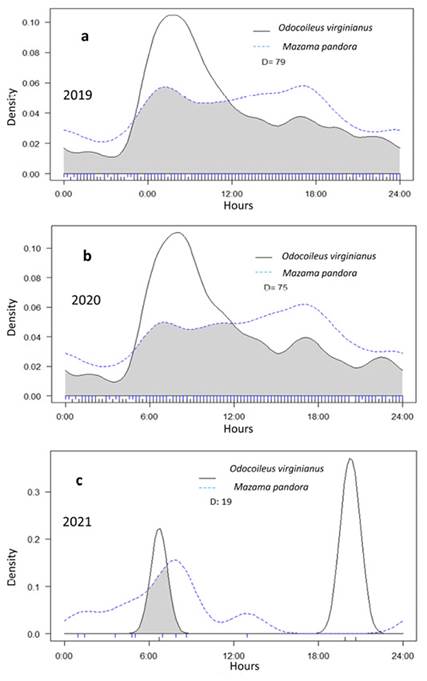
Figure 4 Overlap of the daily activity patterns of O. virginianus and M. pandora for the three years of records (a: 2019, b: 2020, c: 2021) at the CBR, Campeche, México. The 95 % coefficient of overlap is the area under the curve of both density estimates (marked in gray).
Similarly, the visit rate was higher than that estimated for O. virginianus in a WMU in the Tehuacán-Cuicatlán Biosphere Reserve (RBT-C) where the species is utilized (Mandujano- Rodríguez and Hernández 2019). In the present study in the Maya Forest, the visit rate of deer decreased significantly in 2020 and 2021. This result differs from the finding reported by Mandujano-Rodríguez and Hernández (2019), who recorded a larger number of records in the last sampling year (2018). These authors commented that this finding could be a result of the fact that, over time, deer have learned to recognize water troughs as alternative water sources and resort to them once they have located them (Berbert and Fagan 2012). Additionally, these authors considered that 2018 had more adverse climatic conditions (higher temperature and lower precipitation), so deer visited the water troughs more frequently to meet their water needs (Fuller et al. 2014).
The lower number of deer records in 2020 and 2021 is likely due to the high water availability in natural reservoirs since 2020 was a relatively rainy year where the first rains occurred in April (Hernández-Cerda et al. 2021), followed by tropical storm systems in May (Arthur and Bertha); however, it was Tropical Storm Cristobal (June 1 to 8) that saturated the soils of the region (NOAA 2020), flooding a large area of the Calakmul Biosphere Reserve, so natural water bodies supplied water until the middle of 2021 (Contreras-Moreno pers. comm.).
The use of water troughs has been scarcely analyzed in México. However, mammals have been reported to respond positively (Delgado-Martínez et al. 2018; Mandujano-Rodríguez and Hernández 2019; Mandujano-Rodríguez and Hernández-Gómez 2019; Borges-Zapata et al. 2020), and are a common practice within wildlife management units (WMU; Gastelum-Mendoza et al. 2014).
In the CBR, deer used water troughs regularly during the peak drought season when natural water bodies were dry, confirming that troughs effectively supply water to these herbivores and are a viable option to mitigate the effects of water scarcity in the Calakmul region, as is the case for other mammals in the region (Contreras-Moreno et al. 2019; Borges-Zapata et al. 2020). No significant differences were observed in the use of water troughs between O. virginianus and M. pandora, indicating the importance of water troughs for both species. Therefore, the water management and supply carried out likely contribute to the conservation of both species.

Figure 5 Photographs of the three deer species recorded in artificial water troughs in the present study. 5a: O. virginianus, 5b: M. pandora, 5c: M. temama.
A low number of records of M. temama in water troughs were captured. This finding has generally been observed within the CBR for this species (Ramírez-Ortiz 2016), since the distribution of this species is associated with wetter areas, such as those adjacent to the reserve (Ramírez-Ortiz 2016). M. temama is considered a frugivorous species, at least to a greater extent than white-tailed deer (Weber 2008), which would also explain why M. temama visits water bodies, including water troughs, less frequently.
In this regard, climate changes may affect the structure of ecological communities, triggering changes in resource distribution, abundance, and phenology (Kardol et al. 2010). This situation becomes adverse in places where water has been identified as a limiting resource since rainfall is a driver of many animal movements (Bello et al. 2004). The activity pattern of deer when visiting water troughs was mainly diurnal (Figures 4 and 5), probably because it is much safer for both species to move during the day. Predators and meso predators have been frequently recorded in the study area, which are active mainly during nighttime hours (Sima-Panti et al. 2020; Contreras-Moreno et al. 2019b, 2020, 2021). Deer often adapt their behavior to manage their time between food search and protection (Schmitz 1991).
Several studies have documented that prey adapt their behavior to minimize the risk of predation by their natural predators (Mukherjee and Heithaus 2013). In some cases, hunting can force ungulates to modify their visits to water troughs from day to night, but the magnitude of this change may be limited by the risk of predation imposed by large nocturnal carnivores (Crosmary et al. 2012).
The activity patterns of O. virginianus observed in the present study are similar to those recorded in Laguna de Términos, Campeche (Rodríguez 2015) and Calakmul (Ramírez-Ortiz 2016). It is worth mentioning that in both studies, camera traps were not directed to water troughs, suggesting that deer activity patterns are not affected by artificial water troughs. Similarly, our findings in the present study for O. virginianus are similar to those found in the RBT-C (López-Tello et al. 2015), which is noteworthy since RBT-C is a semiarid site. However, in some other cases, it has been observed that deer are usually active in twilight hours in the dry season and in daytime hours during the rainy season (Beier and McCullough 1990; Cornicelli et al. 1996; Sánchez-Rojas et al. 1997; Galindo-Leal and Weber 1998; Gallina et al. 2005). Adjustments of deer to their activity pattern may be related to resource availability (Sánchez-Rojas et al. 1997).
In the case of M. pandora, the activity pattern recorded in the present study showed activity peaks at dawn and dusk. However, in Calakmul, M. pandora recorded daytime activity, with peaks of activity at noon (Ramírez-Ortiz 2016). To note, these species forage throughout the day and probably also during the night to minimize the risk of predation by large felines (Mandujano-Rodríguez and Hernández 2019). The degree of overlap recorded between two species of deer (O. virginianus and M. pandora) suggests that both species are active at the same hours of the day.
Furthermore, it was observed that in addition to allowing access to fresh water, water troughs facilitate interactions between individuals of different populations. Hence, these could be considered sites that favor socialization and predation, and a specific approach is needed to understand these behaviors. Similarly, additional studies are needed to address both the consumption patterns of deer in all seasons and ways to improve the effectiveness of the current water trough network.
For the Yucatan Peninsula in particular, several models have been generated that suggest a disruption in precipitation patterns (Mardero et al. 2020), and it is considered that deer will be directly affected by these alterations derived from climate change (Contreras-Moreno and Torres-Ventura 2018). These models suggest that temperature will rise and precipitation will decrease in the Maya Forest in the near future (O'Farrill et al. 2014). It has been proposed that water scarcity could lead to greater intra- and inter-specific negative interactions between fauna and increase metabolic costs (as animals are forced to move further away to find water supplies; Delgado-Martinez et al. 2018).
Recently, events have been reported of tapirs going into villages, livestock pastures, hive plots, and roads searching for water, which has raised the level of interaction with people, leading to conflicts where these ungulates can be injured or killed (Pérez-Flores et al. 2021). In this sense, there is high pressure to hunt deer in some ejidos of the Maya Forest (Reyna-Hurtado and Tanner 2007), and water shortages may foster the hunting of these ungulates. Artificial water troughs could play a key role in mitigating these adverse effects of drought on deer in the Maya Forest, as they complement the functions of natural water bodies. However, further in-depth research is needed to understand and rule out potential adverse effects on deer populations since it has been suggested that water troughs, being artificial structures arbitrarily introduced into natural systems, may lead to alterations in deer behavior, ecology, and health (i. e., they could be potential foci of zoonoses).

