











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Calakmul Biosphere Reserve is one of the largest natural protected areas in México with 7,231 km2 (Comisión Nacional de Áreas Naturales Protegidas 2023). It belongs to the Mayan rainforest, the biggest extension of tropical forest of North and Central America, and the second in America, just below the Amazon basin (Secretaria de Medio Ambiente y Recursos Naturales 2018; Reyna-Hurtado and Sanchez-Pinzón 2019). Emblematic large mammals such as the jaguar (Panthera onca), the white lipped peccary (Tayassu pecari), and the Central American tapir (Tapirus bairdii) still inhabit this forest (Carabias-Lillo et al. 1999; Reyna-Hurtado and Sanchez-Pinzón 2019), as well as species of high cultural and alimentary importance for local communities, such as the white-tailed deer (Odocoileus virginianus) and two species of brocket deer (Odocoileus pandora and Mazama temama; Weber 2000, 2005; Reyna-Hurtado and Tanner 2005). However, this protected area is subjected to water extraction, logging, road construction, agriculture and extensive livestock farming (Weber 2008). In addition, in recent years, land clearing has increased due the construction of the “Tren Maya” megaproject and the touristic-urban complex “Hotel Maya” within the reserve, and these projects are expected to generate barrier effects (blocking of movement and trampling) and edge effects (due to noise; Alayón-Gamboa et al. 2019; Benítez et al. 2021).
The white-tailed deer (Figure 1c) is the most widely distributed deer in the Americas, historically ranging from southern Canada (60° North Latitude), to southern Peru (15° South Latitude), and has been introduced to New Zealand, Czechoslovakia and Finland (Ceballos 2014; Gallina et al. 2014; Gallina and López-Arévalo 2016). Its wide distribution is due to its high adaptability, evident in its high morphological, physiological, biochemical and behavioral variation (Weber 2014; Gallina et al. 2014; Gallina and López-Arévalo 2016). The tropical subspecies have a dark coat color in humid rainforest, being reddish in subtropical environments (Weber 2014; Gallina and López-Arévalo 2016). They are usually medium to small sized, with males weighting less than 60 kg and females less than 35 kg (Gallina et al. 2010; Weber 2014). In the greater Calakmul Region, in areas with little hunting it has been described as an “opportunistic”, using habitat by its availability, showing preference for secondary vegetation and agricultural areas (Weber 2005; Reyna-Hurtado and Tanner 2005; Weber 2008; González-Marín et al. 2008), preferring the low-flooded forest in areas with high hunting pressure (Reyna-Hurtado and Tanner 2005; Briceño-Méndez and Contreras-Moreno 2020).
Yucatán brown brocket deer (Figure 1b), recently classified within the genus Odocoileus (Gutiérrez et al. 2017), is a medium-sized cervid. In the Great Calakmul Region the average weight of 13 captured male brocket deer was 20.5 kg, and that of eight adult females was 17.3 kg (Weber and Medellín 2010; Weber 2014). The dorsal coat is uniform brown to brown grayish, becoming paler in the ventral area. It has a tuft of bristle-like stiff hair between the eyes and in the forehead. The antlers have massive pedicels and are large, divergent, often curving so the tips are convergent, but with a wide space between them (Weber 2014). The brown brocket deer is distributed in the Yucatán Peninsula, in the states of Yucatán, Campeche, and Quintana Roo, México and in northern Guatemala and northern Belize (Weber and Medellín 2010; Weber 2014; Weber et al. 2016). It has been described as a generalist species, using habitat due to its availability, showing no preference for a vegetation type (Weber 2005; Weber 2014).
Central American red brocket deer (Figure 1a) is the smallest of the three deer species we discuss (Weber 2014) weighting 12 to 32 kg; in the Greater Calakmul Region the average weight of five adult males was 16.3 kg and that of four adult females was 13.2 kg (Bello-Gutiérrez et al. 2010). Its dorsal coat its deeply red, with darker tones on neck and head, and whitish ventral parts of the body and tail. Antlers are simple, lacking ramifications, growing no more than two inches (Bello-Gutiérrez et al. 2010; Weber 2014). The limits of its distribution are unclear, having records in México, in the states of Tamaulipas, San Luis Potosí, Veracruz, Puebla, Oaxaca, Chiapas, Tabasco, Campeche, and Quintana Roo, northern and central Belize, Guatemala, El Salvador, Costa Rica, Nicaragua, Honduras, Panama, and western Colombia (Bello-Gutiérrez et al. 2010; Bello et al. 2016; Moreira-Ramírez et al. 2019). It is a habitat specialist, occupying well-preserved humid forest (Medellín and Equihua 1998; Weber 2005; Tejeda-Cruz et al. 2009; García-Marmolejo et al. 2013; García-Marmolejo et al. 2015), occasionally using secondary vegetation, as long as there are well-preserved surrounding forest or vegetation that constitutes a high percentage of its diet, like the palm Sabal mexicana (Tejeda-Cruz et al. 2009; García-Marmolejo et al. 2015), in areas with high hunting pressure, it uses low-flooded forest (Reyna-Hurtado and Tanner 2005).

Figure 1 a) Central American red brocket deer (Mazama temama). b) Yucatan brown brocket deer (Odocoileus pandora). c) White tailed-deer (O. virginianus).
Camera-trapping has been an important tool for the study of these and other medium-sized and large mammals, covering aspects ranging from species richness, abundance indices, density, occupancy, activity patterns, and even more complex aspects such as intraspecific, interspecific and environmental interactions (Mandujano 2019). In recent years, occupancy models are being used more frequently to evaluate the presence or absence of cryptic species and the relationship of the species with several variables of the landscape (MacKenzie et al. 2011). The advantage of occupancy models is that they incorporate the probability of detection, a parameter that can vary when species are cryptic, shy, exist in low densities, or when the environment changes make it difficult to sample the presence of a specific species (MacKenzie et al. 2011).
Through occupancy models and several others ecological estimators of wildlife populations parameters, knowledge about fauna has expanded, allowing for more structured conservation strategies (Bruner et al. 2001). Food, water sources, type and vegetation cover, distance from roads and human presence are factors that limits the presence, movement and size of populations of ungulates. The effects of those vary among species and from regions (Lira-Torres and Naranjo Piñera 2003; Weber 2008; Pérez-Cortez et al. 2012; García-Marmolejo et al. 2013). However, basic ecological aspects as activity patterns, occupancy and the factors involved in these, have been poorly studied for these deer species (Ramírez 2016; Moreira-Ramírez et al. 2019). The aim of this study was to evaluate the probabilities of occupation, local colonization, local extinction, detection, activity patterns, activity overlap and relative abundance of the three species of deer in the ponds of Calakmul Biosphere Reserve, through nine years of photo-trapping, contributing to the understanding of the ecological patterns and conservation of Neotropical deer that live in sympatry.
Materials and methods
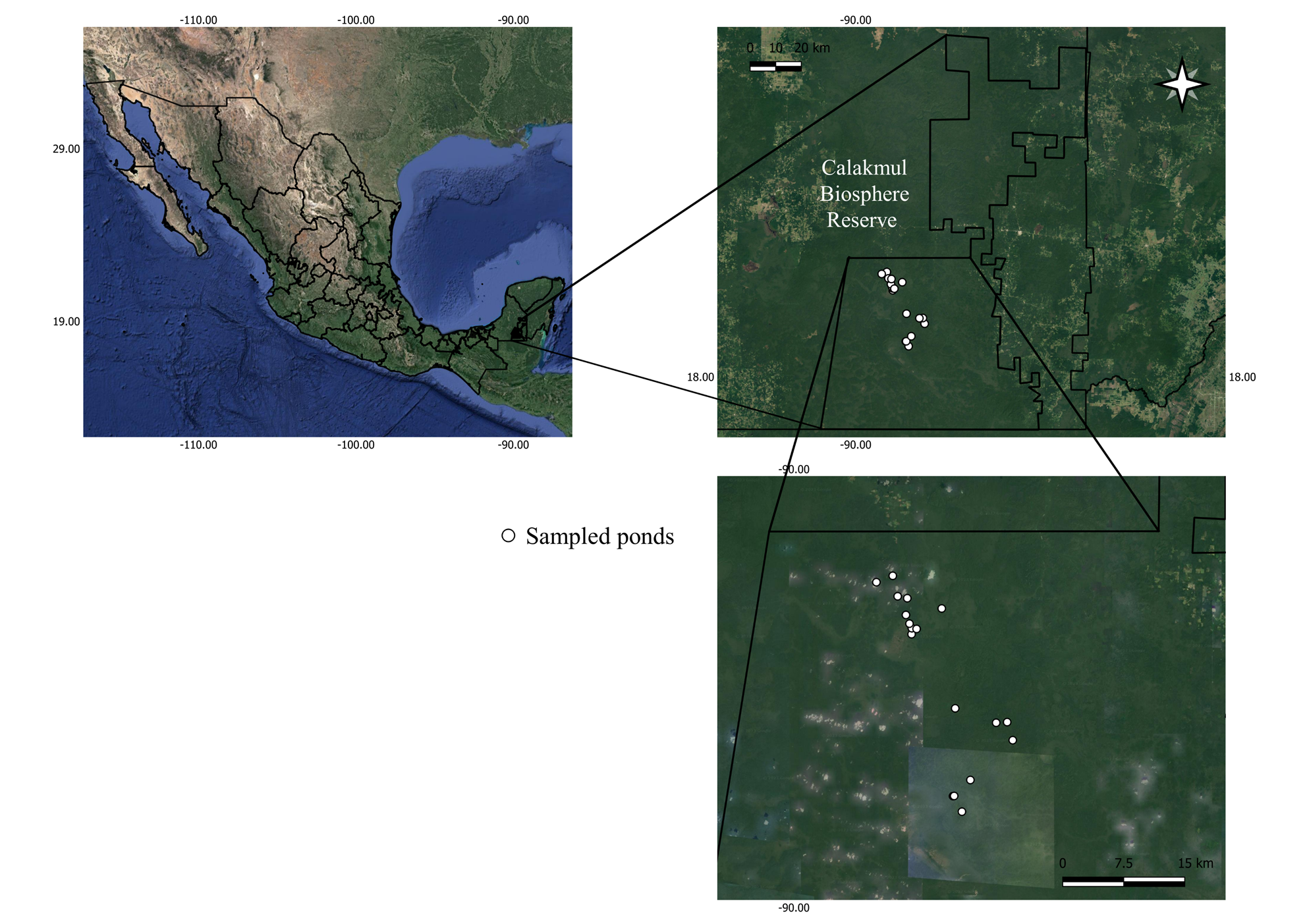
Study area. Calakmul Biosphere Reserve (CBR) is a protected area decreed in 1989, with an extension of 7,231 km2, located in the municipality of Calakmul, southeast Campeche, 19° 15’ 17” N, -90° 10’ 89” W (Carabias-Lilloet al.1999). The reserve presents a tropical sub-humid climate, with a rainy season in summer (Jun to November), having an annual mean temperature of 24.6 °C, and an annual mean precipitation of 1,076 mm (Martínez and Galindo-Leal 2002). The reserve's hydrography is determined by the amount of rainfall, which is capable of flooding lowlands and forming temporary and permanent ponds and surface stream beds Carabias-Lillo et al. 1999). The ponds are vital components of the landscape for wildlife, being one of the few water sources of the region in the dry season (Reyna-Hurtado et al. 2010; Pérez-Cortez et al. 2012). The main vegetation types in Calakmul are high evergreen forest, medium sub evergreen forest, low evergreen and deciduous forest, savanna and hydrophytes (Pennington and Sarhukan 1998; Carabias-Lillo et al. 1999; Martínez and Galindo-Leal 2002).
Camera trapping. As part of a long-term study focused on wildlife uses of ponds in CBR conducted by R. Reyna-Hurtado and collaborators, camera trapping survey was conducted from February 1st 2014 to December 31st 2022 in as many as 17 ponds (Figure 2). A single camera trap was installed in each of a set of 10 to 17 ponds that were monitored along these years (Reconyx PC800 Hyperfire professional Reconyx, Inc., Cuddeback Inc and Browning Strike Force Co.). Cameras were positioned 50 cm above the ground, and programmed to take five pictures every time the sensor detected movement. The cameras were checked every two months on average to change batteries and memory cards.
Relative abundance. To ensure the independence of the records, photographs obtained of the same species where individuals could not be recognized were selected in a 60-minute period, considering them as a single event (Tobler et al. 2008). To estimate the relative abundance index, we took the number of independent records of a selected species divided by the number of camera-nights, multiplied by 100 (O’Brien 2011; Mandujano 2019).
Occupancy. Following the methodology used by Louvrier et al. (2017) to calculate the probability of occupancy, local colonization, local extinction, and detection in the ponds over the nine years of camera-trapping, we used the dynamic model of occupancy designed by MacKenzie et al. (2003). The sampling units were the years from 2014 to 2022 and the secondary occasions were defined according to the months of the year (1 to 12). We created a detection history per species, where we pooled the records of detection (1) and non-detection (0) per periods. Assuming that occupancy (Ψ) is the probability that a site is occupied by the target species, p tj is the probability of detecting the target specie, given presence, in survey j within primary period t (MacKenzie et al. 2003). Local colonization (γ) is the probability that an unoccupied site in a primary sampling period (t) is occupied in t + 1, and local extinction ( ϵ ) is the probability that an occupied site in a primary sampling period (t) is unoccupied in t + 1 (MacKenzie et al. 2003). For model selection we used the Akaike’s information criterion (AIC; Burnham and Anderson 2002), models were performed using the Unmarked package in R (Kellner et al. 2023).
To limit the number of models we followed a hierarchical approach (Olson et al. 2005; Betts et al. 2008), and to evaluate the goodness of fit and the dispersion parameter (c-hat) of the best single species model we followed the procedure used by Andrade-Ponce et al (2022). First, we created a model for each of the covariates to see their effects on the detection of the species, applying the Akaike's selection criterion to select the best one, similarly we performed these steps for occupation, local colonization and local extinction. The resulting models were contrasted with Akaike's selection criterion to choose the best model(s).
The covariates used in this study were selected according to the possibility of their importance for the species at the time of visiting the ponds. a) The distance from the ponds to the road. b) The type of vegetation that predominates around the ponds following the classification of Pennington and Sarhukan (1998). c) Disturbance, measured as the influx of people to the ponds, being a "medium" level, when they are visited by this or another research group and/or having a medium level of tourism and being a "low" level, when they are only visited by our research group. d) The presence or absence of water. e) The year as a linear effect.
To determine a possible relationship between deer species occupancy and the presence of water, we used a multispecies model, since single-season or multi-season occupancy models consider that the variables affecting occupancy do not change during the entire study period (MacKenzie et al. 2003), while the presence or absence of water in the ponds varies over months and years. Therefore, to see its possible effect, water presence was analyzed as another species, solely for analysis purposes in the unmarked program (Kellner et al. 2023). The multispecies model allows estimating the probability of two or more species occurring simultaneously, as a function of covariates, using a multivariate Bernoulli distribution (Rota et al. 2016). For the analysis, a detection history was created for the presence of water and a null occupancy model was performed, to visualize if there was any type of interaction, and if it was significant. Finally, the species interaction factor (SIF) was estimated to corroborate the type of interaction between deer and water, where values equal to one suggest independence, values less than one indicate an inversely proportional relationship and values greater than one indicate a positive relationship (Richmond 2010). The analysis was performed through the Unmarked package in R (Kellner et al. 2023).
Activity patterns. Following the methodology used by Andrade-Ponce et al. (2022) we used the Kernel density estimation method, with the Von Mises distribution for circular data to estimate the overlap activity patterns (Ridout and Linkie 2009). To compare the overlap of daily activities of a pair of species we use the overlap coefficient (Δ), where values close to one reflect a similar activity pattern and values close to cero mark different activity patterns. In addition, we calculated the 95 % CI of each overlap coefficient with 10,000 bootstrap tests (Ridout and Linkie 2009). To assess whether the overlap coefficient was statistically significant, a randomized test was performed. If the resulting P-value is less than 0.05 it can be inferred that the Kernel circular probability distributions differ and therefore the resulting overlap is the result of chance. The packages Activity and Overlap were used to perform the tests in R version 2023.09.1+494 (Ridout and Linkie 2009; Rowcliffe 2023; R Core Team 2023).
Results
Relative Abundance. During the nine years of camera-trapping, with an effort of 31,217 camera-nights, 225 independent records were obtained for Yucatan brown brocket deer, only 16 for Central American red brocket deer and 2,026 for white-tailed deer, the last species having the highest relative abundance (RAI. = 6.49; Table 1), followed by the Yucatan brown brocket deer (RAI. = 0.72; Table 1), and Central American red brocket deer having the lowest value (RAI. = 0.05; Table 1).
Of the nine study years, the one with the highest relative abundance for Odocoileus virginianus was 2022 (RAI. = 13.29; Table 1) and the lowest was 2014 (RAI. = 3.47; Table 1). For Odocoileus pandora the highest year was 2019 (RAI. = 1.51; Table 1) and the lowest was 2020 (RAI. = 0.21; Table 1). Mazama temama was not recorded in six of the nine camera-trapping years, the one with the highest relative abundance was 2021 (RAI. = 0.81; Table 1) and the lowest 2022 (RAI. = 0.03; Table 1).
Table 1 Relative abundance index in each of the nine years of photo-trapping.
| Species | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | 9 years |
|---|---|---|---|---|---|---|---|---|---|---|
| Odocoileus pandora | 0.48 | 1.17 | 0.88 | 0.96 | 0.96 | 1.51 | 0.21 | 0.41 | 0.89 | 0.72 |
| Mazama temama | 0.00 | 0.00 | 0.00 | 0.078 | 0.00 | 0.00 | 0.00 | 0.81 | 0.039 | 0.05 |
| Odocoileus virginianus | 3.47 | 10.63 | 3.97 | 7.53 | 7.29 | 4.96 | 3.64 | 5.50 | 13.29 | 6.49 |
Occupancy. Odocoileus pandora. The probability of occupancy (Ψ = 0.98), colonization (γ = 0.0001), and extinction ( ϵ = 0.05) of O. pandora was not related to any of the variables under analysis, however, the probability of detection was negatively influenced by low to medium disturbance of the ponds respectively, with values ranging from p = 0.500 (β = 0.002, P-value = 0.99) to p = 0.077 (β = -2.47, P-value = 0.001; Table 2; Figure 4a). The more human visitation the water pond received, the lower the detection of the species by the cameras. It presented a slightly positive interaction with respect to the presence of water in the ponds (SIF. = 1.01, P-value = 2.85E-43; Figure 4c).
Odocoileus virginianus. Similar to Odocoileus pandora, the probability of occupancy (Ψ = 0.36), colonization (γ = 0.86), and extinction ( ϵ =0.06) were not affected by any of the covariates analyzed for O. virginianus. Detection probability was affected by the type of vegetation surrounding the ponds, with values ranging from p = 0.50 (β = 0.18, P-value = 0.82) in lowland dry forest, p = 0.41 (β = -0.28, P-value = 0.71) in the mid-humid forest and flooded lowland ecotone, p = 0.23 (β = -1.16, P-value = 0.08) in mid-humid forest, and p = 0.19 (β = -1.73, P-value = 0.01) in mid-dry forest (Table 2; Figure 4b). While interaction with water was also slightly positive (SIF. = 1.03 P-value = 7.24E-08; Figure 4e).
Table 2 Dynamic occupancy models for the three deer species in Calakmul Biosphere Reserve from 2014 to 2022. Covariates: Disturbance (Pert); Year; Distance to road (Dist_road); Presence of water (Water); Vegetation type (Veg). Ψ = Occupancy, γ = Colonization, ϵ = Extinction, p = Detection, K = Number of parameters, AICc = Akaike Information Criterion, ΔAICc = Delta Akaike, AICcw = Akaike weight.
| Model | K | AICc | ΔAICc | AICcw |
|---|---|---|---|---|
| Odocoileus pandora | ||||
| Ψ(1);γ(1); ϵ (1);p(Pert) | 6 | 309.54 | 0 | 0.95 |
| Ψ(1);γ(1) ;ϵ (Pert);p(1) | 6 | 316.99 | 7.45 | 0.02 |
| Ψ(1);γ(1); ϵ (1);p(1) | 4 | 318.38 | 8.84 | 0.01 |
| Ψ(1);γ(Year); ϵ (1);p(1) | 5 | 318.38 | 9.26 | 0.01 |
| Ψ(Pert);γ(1); ϵ (1);p(1) | 6 | 321.27 | 11.73 | 0.00 |
| Ψ(1);γ(1 + Year); ϵ (1 + Pert);p(Pert) | 9 | 334.46 | 24.92 | 0.00 |
| Mazama temama | ||||
| Ψ(1);γ(1); ϵ (1);p(1) | 4 | 34.77 | 0 | 0.40 |
| Ψ(1);γ(Year); ϵ (1);p(1) | 5 | 37.20 | 2.43 | 0.12 |
| Ψ(1);γ(Dist_road); ϵ (1);p(1) | 5 | 37.20 | 2.43 | 0.12 |
| Ψ(1);γ(1); ϵ (1);p(Year) | 5 | 37.85 | 3.09 | 0.08 |
| Ψ(1);γ(1); ϵ (1);p(Water) | 5 | 37.85 | 3.09 | 0.08 |
| Ψ(1);γ(1); ϵ (1);p(Pert) | 6 | 37.85 | 3.09 | 0.08 |
| Ψ(1);γ(1); ϵ (1);p(Dist_road) | 5 | 37.85 | 3.09 | 0.08 |
| Ψ(1);γ(Pert); ϵ (1);p(1) | 6 | 40.59 | 5.83 | 0.02 |
| Ψ(1);γ(1); ϵ (Pert);p(1) | 6 | 40.62 | 5.86 | 0.02 |
| Ψ(1);γ(1 + Year + Dist_road + Pert); ϵ (1 + Pert);p(1 + Year + Water + Pert + Dist_road) | 15 | 288.96 | 254.19 | 0 |
| Odocoileus virginianus | ||||
| Ψ(1);γ(1); ϵ (1);p(Veg) | 8 | 418.35 | 0 | 0.93 |
| Ψ(1);γ(1); ϵ (1);p(1) | 4 | 424.24 | 5.89 | 0.05 |
| Ψ(1);γ(1); ϵ (Year);p(1) | 5 | 426.80 | 8.44 | 0.01 |
| Ψ(1);γ(1); ϵ (Pert);p(1) | 6 | 430.44 | 12.08 | 0.00 |
| Ψ(1);γ(1); ϵ (1 + Year + Pert);p(Veg) | 11 | 449.66 | 31.31 | 0.00 |
Mazama temama. Probably due to the few independent records obtained for the Central American red brocket deer, none of the variables under analysis had any effect on the probability of occupancy (Ψ = 0.001), colonization (γ = 0.99), extinction ( ϵ = 0.65), and detection (p = 0.01; Table 2). Being the interaction with water slightly negative (SIF. = 0.93, P-value = 1.84e-01; Figure 4d).
Activity patterns. We detected a predominant diurnal activity in two of the three deer species, white-tailed deer showed two peaks of activity (7:00 to 10:00 and 13:00 to 16:00; Figure 3d), and the Yucatan brown brocket deer showed a relatively constant activity during the day with a large peak from 11:00 to 13:00 (Figure 3a), while the Central American red brocket deer showed a cathemeral activity, having several activity peaks during the whole day (7:00 to 8:00, 10:00 to 11:00, 14:00 to 13:00, 17:00 to 18:00 and 23:00 to 24:00; Figure 3b). We found a high overlap in the daily activities of the three species (Δ > 0.7), the pair of species with the highest overlap coefficient were white-tailed deer and Yucatan brown brocket deer (Δ = 0.86, P-value = 0.63; Figure 3d), followed by white-tailed deer and Central American red brocket deer (Δ = 0.78, P-value = 0.7; Figure 3f), and finally Yucatan brown brocket deer and Central American red brocket deer (Δ = 0.73, P-value = 0.6; Figure 3e).
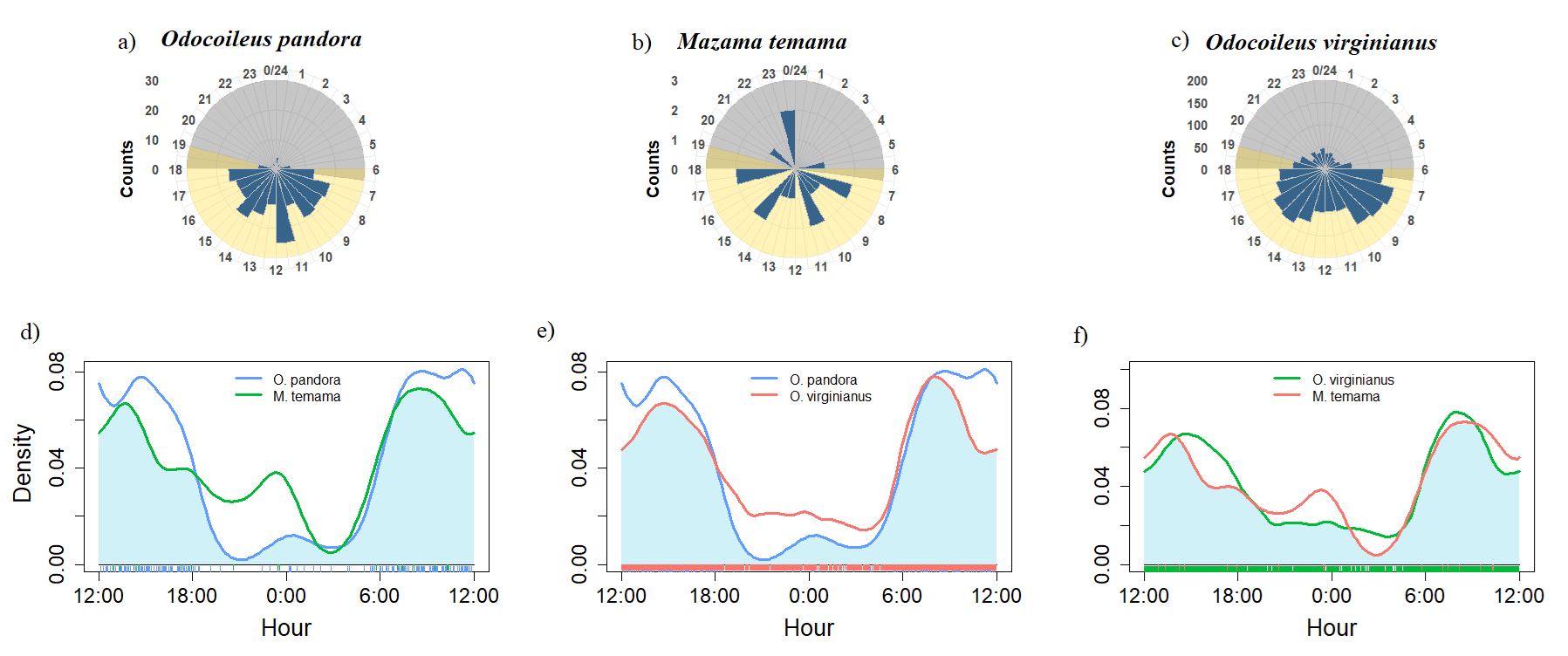
Figure 3 a-c) Number of records per hour for Odocoileus pandora, Mazama temama and O. virginianus. d-f) Kernel activity overlaps between each pair of species in Calakmul Biosphere Reserve, México.
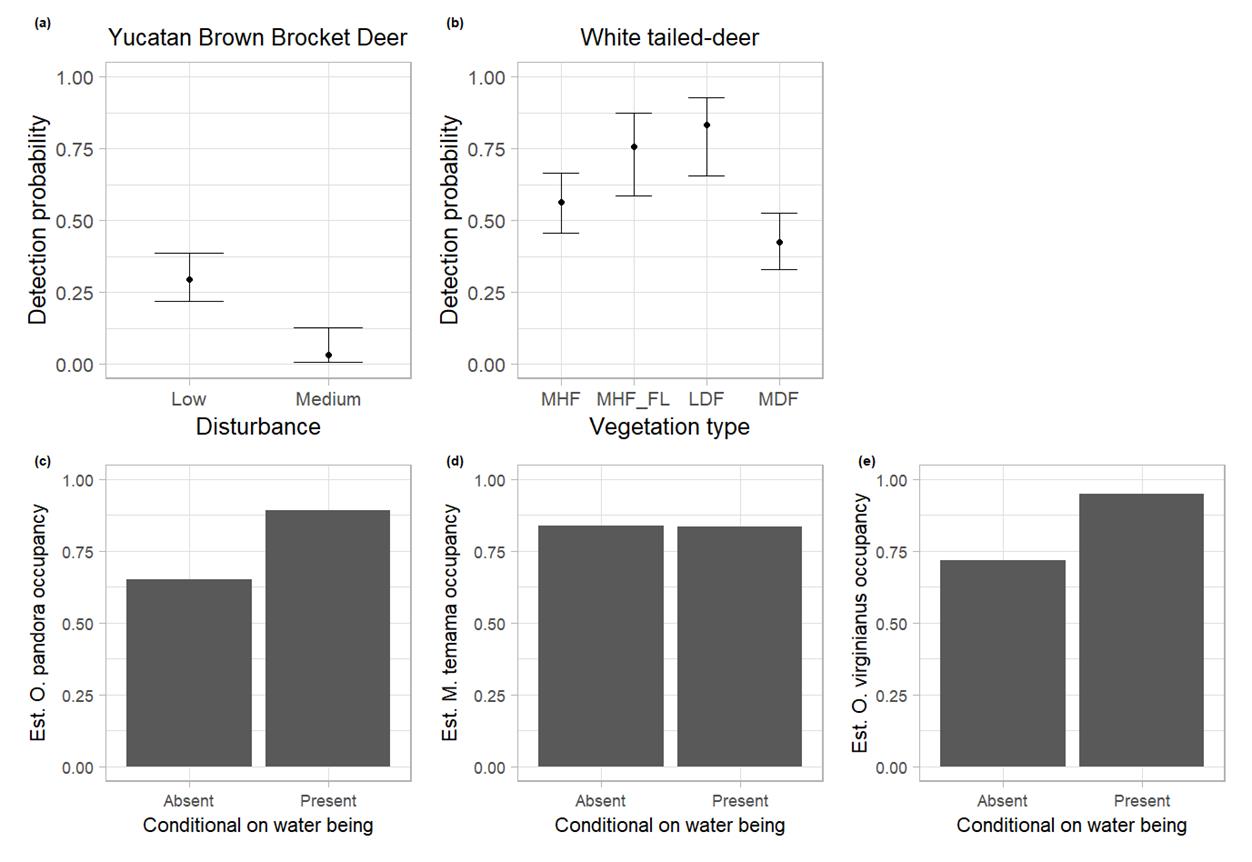
Figure 4 a) Relation between O. pandora Detection and level of disturbance. b) Relation between O. virginianus detection and vegetation type. c-e) Interac tion between the presence or absence of water and the occupancy of the three deer species, in ponds of Calakmul Biosphere Reserve, México. MHF = Mid Humid Forest, MHF_FL = Mid Humid Forest and Flooded Lowland, LDF = Low Dry Forest, MDF = Mid Dry Forest.
Discussion
The type of human disturbance, behavior, habitat and parameters of the deer biology can explain the results obtained for the relative abundance and occupancy of the ponds. The White-tailed deer, despite not occupying a large number of ponds, presented greater abundance, probably due to the ponds being open environments, with a large amount of food in their surroundings. On the other hand, both Gray and Red brocket deer showed a low abundance throughout the nine years, which may be due to their elusive behavior and their more frugivorous diet (Weber 2005), and especially for M. temama, due to their preference for closed environments with dense humid vegetation (Bello-Gutiérrez et al. 2010). This differs from studies conducted in the same area with transects and track counts, in which a greater abundance of Brocket deer is reported relative to White-tailed deer, but because they are different methods, the results often differ from each other (Weber 2005; Reyna-Hurtado and Tanner 2007).
Similarly, other studies throughout México report abundances of M. temama, being the highest in the El Triunfo Biosphere Reserve in Chiapas (Lira-Torres and Naranjo-Piñera 2003) and in the ejido of Nuevo Becal, Campeche (Ramírez 2016), with the species been more abundant in humid well-conserved areas. In the case of O. virginianus, its abundance is very fluctuating, due to its ability to adapt to disturbed areas, for example in San Luis Potosí, it presented an abundance of 5.6 in ecosystems of early successional stages and an abundance of 2.8 in those of medium successional stages. The abundance of O. pandora has been poorly studied outside of Campeche, and most studies lump the two brocket deer species into a single group (Weber 2005; Reyna-Hurtado and Tanner 2007), making it difficult to compare the abundance of these species. However, the results obtained in this study should be taken with caution, as they may be biased, because the positioning of the cameras was not random, but were placed specifically in ponds (Reyna-Hurtado et al. 2019).
The Yucatan brown brocket deer had the highest probability of occupancy in the ponds out of the three deer species throughout the nine years (despite not being very abundant), and therefore presented very low probabilities of colonization and extinction; however, none of the variables analyzed had an effect on these. This could be due to the fact that this species has been defined as a generalist species (Weber 2005), adapting well to the diverse types of environments that the CBR has (Reyna-Hurtado and Tanner 2005), and to the different degrees of disturbance, either by human presence and/or distance to the roads. Previous studies have shown the adaptability of O. pandora to these environments, presenting high abundances for the species (Weber 2005; Reyna-Hurtado and Tanner 2007; Ramírez 2016), however, it has also been reported that this species is susceptible to hunting and that the presence of this activity has a negative effect on occupancy (Urquiza-Haas et al. 2009). The probability of detection was affected by the disturbance at the watering holes, decreasing with increased human presence, probably due to the species solitary and nervous nature, leading them to a selective use of watering holes with little human influx (Weber 2014).
White-tailed deer showed a low occupancy of the ponds, but with a high probability of colonization, and a low probability of extinction, this can be explained by the high amount of food found in these forest clearings, where herbaceous plants are abundant and are important resources for this species (García-Marmolejo et al. 2015; Ramírez 2016), although none of the variables had an effect on these, as in studies conducted in Nicaragua (Zeller et al. 2011). On the other hand, detection probability was influenced by the type of vegetation, varying among the four types of vegetation analyzed. Although this species is classified as a generalist (Weber 2005) and has great plasticity, allowing it to inhabit a wide variety of ecosystems (Weber 2014; Gallina et al. 2014; Gallina and López-Arévalo 2016), it has shown a preference for dry and flooded lowland in the CBR. These habitats presented the highest probability of detection, followed by medium-humid and dry forests, which the species uses in proportion to their availability (Reyna-Hurtado and Tanner 2005).
In the case of Central American red brocket deer, both the probability of occupancy and detection were very low and the probability of colonization and extinction were high, probably due to the low abundance of this species in the reserve in this and other studies (Weber 2005; Ramírez 2016), as well as to the prevalence of non-optimal environments for this species, which prefers tropical forests with high humidity and high and dense vegetation cover, as a means of protection against predators (Bello-Gutierrez et al. 2010; Ramírez 2016). In studies conducted in a nearby community forest (Ejido Nuevo Becal) the presence of this species was higher (Ramírez 2016) and there is even a sport hunting company focusing entirely on legally harvesting this species (Reyna-Hurtado pers. Obs.). Studies conducted in Los Chimalapas, Oaxaca, showed that the distance to the villages had a positive effect on the occupancy of M. temama and that the probability of detection is low, due to its cryptic behavior (Pérez-Irineo and Santos-Moreno 2016). Similarly, it has been detected that the presence of hunting has a negative effect on the occupancy of this species (Urquiza-Haas et al. 2009). On the other hand, in Costa Rica, it was determined that the colonization by M. temama was favored in areas with ecologically similar species as the temperature increased. Conversely, in areas with ecologically dissimilar species, colonization was favored as the temperature decreased (Beaudrot et al. 2019).
The presence of water in the ponds was an environmental variable of special interest in this work, because it has been demonstrated that it is a vital resource for other ungulate species in the reserve (Reyna-Hurtado and Tanner 2005; Reyna-Hurtado et al. 2012; Pérez-Cortez et al. 2012; Sandoval-Seres et al. 2016; Sánchez-Pinzón et al. 2020). White-tailed deer as well as Yucatan brown brocket deer showed a slight positive interaction with the presence of water, allowing us to infer that the presence of the resource may be a factor that influences the visit to the ponds, however it is uncertain if it is for consumption or for some other reason, such as a greater amount of some herbaceous species. In other regions, water has been found to be a determining factor in the activities of white-tailed deer, altering their behavior (Gallina et al. 1998) or extending their home range when the resource is scarce (Kroll 1992; Contreras-Moreno et al. 2021). The importance of water availability in the behavior and survival of this species (and probably of other cervids) is clear (Bello et al. 2001). The interaction of the Central American brocket deer with water was slightly negative. This finding is inconsistent with other studies in the tropics, which report the importance of water for the brocket deer species (Zeller et al. 2011). The CBR represents the driest portion of the area and M. temama prefers humid areas, such as the adjacent community areas to the CBR, where the species seems to be more abundant (Ramírez 2016; Reyna-Hurtado unpublished data). This discordance also may be due to the low number of records we obtained for this species.
Two of the three deer species studied, the white-tailed and the Yucatan brown brocket deer showed daily diurnal activity; the white-tailed deer had activity peaks during most of the day, but avoiding the hottest hours (11:00 to 13:00), agreeing with other studies for this species in the east and center of Mexico (Hernández-SaintMartín et al. 2013; Soria-Díaz and Monroy-Vilchis 2015; Ramírez 2016; Soria-Díaz et al. 2016; Moreira-Ramírez et al. 2019) and Costa Rica (Botts et al. 2020), although contrasting with studies done in arid environments and in Sierra Madre Occidental (Beier and McCullough 1990; Gallina et al. 2005), where activity peaks were reported at dawn and dusk. Likewise, the Yucatan brown brocket deer was active during the day, presenting a large peak between 11:00 and 12:00, agreeing with previous works in Campeche and Guatemala (Ramírez 2016; Moreira-Ramírez et al. 2019). In contrast, the Central American red brocket deer showed cathemeral activity, which is consistent with most studies for this species, both in México (Pérez-Irineo and Santos-Moreno 2016; Moreira-Ramírez et al. 2019; Pérez-Irineo et al. 2021; González et al. 2021; Salazar-Ortiz et al. 2022) and Central America (Arroyo-Arce et al. 2016; Arroyo-Arce et al. 2017; Moreira-Ramírez et al. 2019; Botts et al. 2020), varying at diurnal or nocturnal times in few cases (Ramírez 2016; Pérez-Irineo et al. 2021; Moreno et al. 2023).
Activity overlap was high among the three deer species (Δ > 0. 7), which lead us to hypothesize that they are able to coexist in the same environment due to the clear separation in behavior and diet specialization. The white-tailed deer being a habitat generalist browser with high consumption of leaves and stems throughout the year and a few fruits during the wet season; the Yucatan brown brocket deer is also a habitat generalist but is mainly frugivorous during the dry season and a browser in the wet season; and the Central American red brocket deer is a specialist frugivorous of well-conserved and dense habitats (Weber 2005).
In this study it was shown that among the variables analyzed there is not one that influences deer occupancy in the ponds, so it is likely that this is due to a combination of multiple factors and interactions among these species, however, it was seen that these species, like other ungulates, have a positive interaction with the presence of water. Ponds are vital resources in the Calakmul Biosphere Reserve, being one of the few semi-perennial water sources for wildlife during the dry season. Increasing temperatures, desiccation, changing rainfall patterns and human disturbance in the ponds threaten these unique microenvironments, endangering the natural habitats of the species and the biotic community in general, making them priority areas for biodiversity conservation. The CBR is a unique place that hold population of three sympatric species of deer. We hope that this research helps to understand the ecological requirements of these interesting species and how they have coexisted in this tropical, semidry environment.

