











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Los bosques conformados por taxones del género Pinus son de los más abundantes y con amplia distribución en la Sierra Madre Occidental de México (González-Elizondo et al., 2012). Estos ecosistemas han sido ampliamente estudiados, principalmente por las actividades de aprovechamiento forestal (Sánchez et al., 2003). De los 24.7 millones de hectáreas de la superficie total del estado de Chihuahua (norte de México), 7.08 millones (28.66 %) corresponden a superficies de bosques templados (Instituto Nacional de Estadística y Geografía [Inegi], 2023). Los bosques templados de esta zona del país, además de las especies del género Pinus, están conformados por taxa de los géneros Abies, Cupressus, Juniperus, Pseudotsuga y Quercus (Challenger & Soberón, 2008). Debido a la alta demanda de bienes y servicios ambientales por las comunidades rurales y urbanas, los principales ecosistemas aprovechados se integran por bosque de pino y pino-encino (García García et al., 2019), en los cuales se busca un aprovechamiento forestal sostenible (Monárrez-Gonzalez et al., 2020).
El conocimiento de la estructura, composición y diversidad de especies arbóreas en estos bosques es de suma importancia para planear un manejo sustentable y de conservación (Delgado-Zamora et al., 2016). Existen factores que influyen sobre la composición y estructura de los bosques templados; por ejemplo, el clima, el suelo, la precipitación y la forma de aprovechamiento de los árboles (Schaub et al., 2020). De esta manera, la evaluación de la diversidad permite conocer los efectos del aprovechamiento maderable sobre las especies arbóreas (Aguirre Calderón et al., 2008; García García et al., 2019).
La caracterización de la estructura permite estimar parámetros de la estructura horizontal, tales como el Índice de Valor de Importancia, dominancia, frecuencia y abundancia (Jiménez et al., 2001). También, incluye variables de la estructura vertical como diámetro, altura y cobertura, lo que genera índices cuantitativos que aportan información con fines de conservación (Tadeo-Noble et al., 2019).
Para lograr los objetivos del aprovechamiento maderable en el manejo forestal, aquellos atributos del bosque que más se modifican son su estructura, composición y diversidad (Ramírez-Santiago et al., 2019). La comprensión profunda de la estructura, diversidad y dinámica del bosque, junto con el impacto del aprovechamiento forestal en estos aspectos, es crucial cuando se trata de tomar decisiones informadas sobre el manejo forestal y defender su integridad ecológica (Lutz & Halpern, 2006). Es por ello, que una correcta protección forestal en la producción de madera requiere prácticas de manejo apropiadas y reservas en áreas disponibles para el manejo forestal (Aguirre-Calderón, 2015). Por lo tanto, la presente investigación tuvo como objetivo describir la estructura, composición y producción de un bosque de pino bajo aprovechamiento forestal, en el suroeste del estado de Chihuahua, México.
Materiales y Métodos
Área de estudio
La investigación se llevó a cabo en una propiedad forestal privada, con una superficie aproximada de 189.1 ha, correspondiente a un bosque templado frio dominado por Pinus arizonica Engelm., en el municipio Guachochi, Chihuahua, México; dentro de la cadena montañosa Sierra Madre Occidental. El lugar está enmarcado geográficamente entre los 26°45′45.69″ y 26°55′33.28″ norte y 107°7′15.97″ y 107°8′26.23″ oeste (Figura 1).
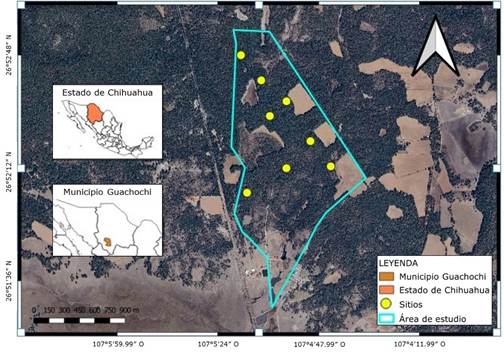
Los suelos son de tipo Cambisol éutrico, Regosol éutrico y Planasol éutrico, de textura media a fina (Inegi, 2024).
El clima se clasifica como C(E)(w2)(x'), correspondiente a un clima húmedo-semifrío, con una temperatura promedio anual de 5 a 12 °C (Instituto Nacional de Estadística, Geografía e Informática [INEGI], 2008).
Evaluación en campo
La información proviene de un inventario forestal realizado en ocho parcelas permanentes de monitoreo de forma circular (1 000 m2), de evaluación y seguimiento silvícola, establecidas en un área destinada a la producción de maderas blandas. De cada individuo presente se obtuvo la especie, altura total con un hipsómetro Suunto ® modelo PM-5/1520 y el diámetro a la altura del pecho (DAP) se estimó con una cinta diamétrica Forestry Suppliers ® modelo 283D.
Para este análisis se definieron tres etapas del manejo forestal. La primera se denominó Condición Inicial [CI (2012)], en la que se consideraron árboles que permanecieron en pie después de la cosecha en el otoño de 2012. La segunda se describe como Reclutamiento [Rec (2022)] en la que se incluyó a individuos no aprovechados y los incorporados que tienen una dimensión comercial (DAP>7.6 cm); esta información se registró en el verano del año 2022. La tercera etapa se denominó Condición Final [CF (2022)] y estuvo conformada por individuos no seleccionados para la tala o árboles residuales de otoño del año 2022.
Análisis de la información
La caracterización horizontal de la comunidad vegetal se hizo mediante el cálculo del Índice de Valor de Importancia (IVI), para lo cual se estimó la dominancia (área basal ocupada por especie), abundancia (número de individuos de la especie) y frecuencia (presencia de la especie). Con estos valores se determinó la importancia de cada especie dentro de la comunidad vegetal (Curtis, 1959). Además, se calculó la producción por medio de la estimación del volumen por especie (Jiménez, 1990).
La riqueza de especies se obtuvo con el Índice de Margalef (Mg) en el que cuanto mayor sea el valor, mayor es la riqueza relativa de especies en el área (Mulya et al., 2021). Para cuantificar la diversidad de especies se utilizó el Índice de Shannon (H′) (Magurran, 2004) que se usa para evaluar cambios en la diversidad a través del tiempo; así, un valor alto indica más diversidad en el área. Como complemento, se calculó el Índice de Equitatividad (E), que proporciona información sobre la uniformidad en la distribución de individuos entre las especies en un intervalo que varía de 0 a 1. Para medir la probabilidad de que dos individuos seleccionados al azar en el área de estudio pertenezcan a la misma especie, se estimó el Índice de Simpson (D), con el cual un valor alto indica una comunidad dominada por pocas especies y un valor bajo corresponde a una mayor diversidad de especies. Para más claridad en los datos, se calculó el índice complementario expresado como 1-D, con el que un valor cercano a 1 representa mayor diversidad y un valor cercano a 0 indica menor diversidad (Magurran, 2004). Las fórmulas utilizadas para determinar los índices estructurales, diversidad y similitud se presentan en el Cuadro 1.
Cuadro 1 Fórmulas utilizadas para determinar los índices estructurales, diversidad y similitud de las especies.
| Fórmula | Descripción de componentes |
|---|---|
|
|
A
i
=Abundancia absoluta AR i = Abundancia relativa por especie N i = Número de individuos de la especie S = Superficie del muestreo (ha) |
|
|
D
i
= Dominancia absoluta DR i = Dominancia relativa de la especie respecto a la dominancia total G i = Área basal de la especie S = Superficie (ha) |
|
|
F
i
= Frecuencia absoluta FR i = Frecuencia relativa de la especie respecto a la frecuencia total P i = Número de sitios en los que está presente la especie NS = Número total de sitios de muestreo |
|
|
IVI = Índice de Valor de
Importancia AR i = Abundancia relativa por especie respecto a la densidad total DR i = Dominancia relativa de la especie respecto a la dominancia total FR i = Frecuencia relativa de la especie respecto a la frecuencia total |
|
|
I = Índice de
Margalef S = Número de especies ln = Logaritmo natural N = Número total de individuos |
|
|
H′ = Índice de diversidad de
Shannon S = Número de especies P i = Proporción de las especies ln = Logaritmo natural N = Número total de individuos n i = Número de individuos de la especie |
|
|
E = Índice de Equitatividad
H′ = Índice de diversidad de Shannon S = Número de especies P i = Proporción de las especies ln = Logaritmo natural N = Número total de individuos n i = Número de individuos de la especie |
|
|
D = Índice de
Simpson P i = Proporción de las especies N = Número total de individuos n i = Número de individuos de la especie |
|
|
A = Área basal
π = Valor constante (Pi=3.1416) d = Diámetro |
|
|
V = Volumen g = Superficie circular del diámetro basal CM = Coeficiente mórfico |
Resultados
Estructura horizontal
En total se registraron cuatro especies de árboles. Existen tres taxones con 218 individuos en la etapa de CI, cuatro con 297 individuos en la etapa Rec y cuatro con 195 individuos en la etapa CF (Cuadro 2).
Cuadro 2 Especies inventariadas en el área de estudio.
| Familia | Nombre científico | Nombre común |
Origen | Etapas |
|---|---|---|---|---|
| Pinaceae | Pinus arizonica Engelm. | Pino blanco | Nativo | CI, Rec y CF |
| Pinaceae | Pinus leiophylla Schiede ex Schltdl. & Cham. | Pino chino | Nativo | CI, Rec y CF |
| Ericaceae | Arbutus xalapensis Kunth | Madroño | Nativo | CI, Rec y CF |
| Cupressaceae | Juniperus deppeana Steud. | Táscate | Nativo | Rec y CF |
La especie con el mayor valor de abundancia fue Pinus arizonica con 194 individuos ha-1 en la etapa CI, que representan 71.10 % del arbolado, con un área basal (abundancia) de 75.77 % y una frecuencia de 50 %, en los ocho sitios. Se contabilizaron 250 individuos ha-1 de la misma especie en la etapa Rec, equivalente a 67.34 % del arbolado, con un área basal de 15.9 m2 ha-1 (73.07 %), en los ocho sitios con una frecuencia de 44.44 %. Por otro lado, se identificaron 166 individuos ha-1 de esta especie en el inventario CF, que suman 68.21 % del arbolado, y 10.22 m2 ha-1 de área basal (73.93 %), con 44.44 % de frecuencia (presencia en los ocho sitios).
Pinus leiophylla Schiede ex Schltdl. & Cham. registró una abundancia de 28.44 % para la etapa CI con un área basal de 24.13 % y una frecuencia de 43.75 % en siete sitios. Para Rec, la abundancia fue de 31.99 %, con un área basal de 26.67 %, una frecuencia de 44.44 %, y se le observó en ocho sitios. Por otro lado, para la etapa de CF, su abundancia fue de 30.77 %, con un área basal del 26.67 %, se le consignó en ocho sitios con un porcentaje de 44.44 %.
Arbutus xalapensis Kunth tuvo un porcentaje de abundancia y área basal >1 % en las tres etapas, con una frecuencia en un sitio de 6.25 % para la etapa Inicial, 5.56 % para la etapa de Reclutamiento y Final. Los mismos datos se confirmaron para Juniperus deppeana Steud. que se detectó solamente en la etapa de reclutamiento y en la etapa final (Cuadro 3).
Cuadro 3 Parámetros ecológicos en un bosque de pino durante tres etapas de manejo forestal.
| Especie | Individuos | Abundancia | Área basal | Frecuencia | IVI | |||
|---|---|---|---|---|---|---|---|---|
| N ha -1 | % | m2 ha-1 | % | Núm. sitios | % | |||
| Etapa de Condición Inicial [CI (2012)] | ||||||||
|
Pinus
arizonica Engelm. |
155 | 194 | 71.10 | 10.24 | 75.77 | 8 | 50 | 65.62 |
|
Pinus leiophylla Schiede
ex Schltdl. & Cham. |
62 | 78 | 28.44 | 3.26 | 24.13 | 7 | 43.75 | 32.11 |
|
Arbutus xalapensis Kunth |
1 | 1 | 0.46 | 0.01 | 0.10 | 1 | 6.25 | 2.27 |
| Total | 218 | 273 | 100 | 13.51 | 100 | 16.00 | 100 | 100 |
| Etapa de Reclutamiento [Rec (2022)] | ||||||||
|
Pinus arizonica Engelm. |
200 | 250 | 67.34 | 15.90 | 73.07 | 8 | 44.44 | 61.62 |
|
Pinus leiophylla Schiede ex Schltdl. & Cham. |
95 | 119 | 31.99 | 5.81 | 26.67 | 8 | 44.44 | 34.37 |
|
Arbutus xalapensis Kunth |
1 | 1 | 0.34 | 0.05 | 0.21 | 1 | 5.56 | 2.03 |
|
Juniperus deppeana Steud. |
1 | 1 | 0.34 | 0.01 | 0.05 | 1 | 5.56 | 1.98 |
| Total | 297 | 371 | 100 | 21.76 | 100 | 18.00 | 100 | 100 |
| Etapa de Condición Final [CF (2022)] | ||||||||
|
Pinus
arizonica Engelm. |
133 | 166 | 68.21 | 10.22 | 72.93 | 8 | 44.44 | 61.86 |
|
Pinus
leiophylla Schiede ex Schltdl. & Cham. |
60 | 75 | 30.77 | 3.74 | 26.67 | 8 | 44.44 | 33.96 |
|
Arbutus xalapensis Kunth |
1 | 1 | 0.51 | 0.05 | 0.33 | 1 | 5.56 | 2.13 |
|
Juniperus deppeana Steud. |
1 | 1 | 0.51 | 0.01 | 0.07 | 1 | 5.56 | 2.05 |
| Total | 195 | 244 | 100 | 14.01 | 100 | 18.00 | 100 | 100 |
En las tres etapas resultaron valores similares para el IVI, lo que supone mayor importancia para las primeras dos especies que se describen a continuación: Pinus arizonica 65.62 %, 61.62 % y 61.86 %, respectivamente por etapa; Pinus leiophylla con 32.11 %, 34.37 % y 33.96 %, respectivamente por etapa. Con menor IVI se posiciona a Arbutus xalapensis, los resultados rondan entre 2 y 2.30 % en cada una de las tres etapas; asimismo, Juniperus deppeana mostró resultados de 1.98 % y 2.05 % para la etapa de Reclutamiento y Condición Final, respectivamente (Cuadro 3).
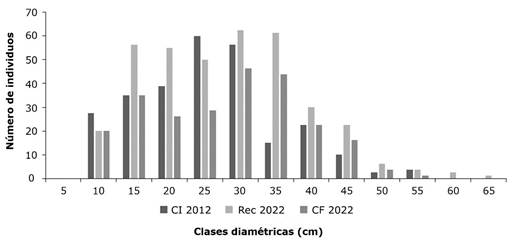
Un parámetro clave es el diámetro del fuste, cuyo análisis, basado en las clases diamétricas registradas según el número de individuos, reveló patrones significativos. En la etapa de Condición Inicial, las categorías de 25-29.99 cm y 30-34.99 cm destacaron por concentrar el mayor número de individuos, con 48 y 45 ejemplares, respectivamente. Para la etapa de Reclutamiento la mayor cantidad de individuos se encontró entre las categorías de 30-34.99 cm con 50 individuos y 35-39.99 cm con 49 individuos, concentrándose la mayor cantidad de los individuos entre los 15-39.99 cm. Para la etapa de Condición Final, la mayor cantidad de individuos se concentró en las categorías 30-34.99 cm con 37 individuos y 35-39.99 cm con 35 individuos, mostrando valores muy similares en las tres etapas donde la mayor cantidad de individuos recae en las categorías que oscilan entre los 30 cm de diámetro con una curva descendente pasando los 40 cm. Solo la etapa Inicial no presentó valores altos en la categoría 35-39.99 cm, presentando una comunidad de arbolado no maduro para las tres etapas (Figura 2).
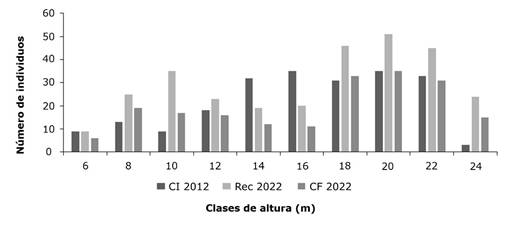
Los resultados de las clases de altura de la comunidad vegetal arbórea mostraron para la etapa Inicial una concentración de la mayor cantidad de individuos entre los 12-23.99 m con 166 de 218 individuos inventariados. Para las etapas posteriores (Rec y CF), la mayor concentración de individuos muestra una altura entre los 16 y los 21.99 m, con 142 y 99 individuos, respectivamente, en cada una de las etapas inventariadas, siguiendo la misma tendencia que las clases diamétricas (Figura 3).
Producción
La mayor producción en volumen se registró para Pinus arizonica durante la etapa de reclutamiento, con 151.38 m3 ha-1, aunque este valor representó un porcentaje similar al de las otras etapas, y varió entre 1 y 2 %. De manera similar, Pinus leiophylla mostró un volumen de 44.80 m3 ha-1 en la etapa de Reclutamiento, con una variación inferior de 2 % respecto a las demás etapas evaluadas. Arbutus xalapensis presentó una contribución inferior a 0.20 % en las tres etapas, mientras que Juniperus deppeana tuvo valores por debajo de 0.02 % tanto en las etapas de Reclutamiento como en la de corta final (Cuadro 4).
Cuadro 4 Producción en volumen para cada especie y por etapa.
| Especie | CI (2012) | Rec (2022) | CF (2022) | |||
|---|---|---|---|---|---|---|
| m3 ha-1 | % | m3 ha-1 | % | m3 ha-1 | % | |
| Pinus arizonica Engelm. | 91.30 | 79.20 | 151.38 | 77.09 | 96.92 | 76.64 |
| Pinus leiophylla Schiede ex Schltdl. & Cham. | 23.95 | 20.78 | 44.80 | 22.82 | 29.36 | 23.22 |
| Arbutus xalapensis Kunth | 0.03 | 0.02 | 0.16 | 0.08 | 0.16 | 0.13 |
| Juniperus deppeana Steud. | - | - | 0.02 | 0.01 | 0.02 | 0.02 |
Riqueza, diversidad y Equitatividad
Los índices muestran variaciones notables en la riqueza, diversidad y equitatividad de especies entre las etapas de Condición Inicial y las etapas de Reclutamiento y Condición Final en 2022. La riqueza estimada y evaluada mediante el Índice de Margalef reveló un incremento progresivo desde CI hacia CF, con una leve mejora en la diversidad específica con el avance en las etapas de recuperación. De forma similar, los valores de diversidad de especies, representados por el Índice de Shannon, también muestran un ligero aumento de CI a CF, lo que sugiere una tendencia positiva en la complejidad ecológica de la comunidad vegetal.
En cuanto a la Equitatividad (E), los valores refieren una distribución relativamente uniforme de especies en CI, que disminuye levemente en Rec y CF, lo cual podría indicar una ligera dominancia de ciertas especies en las etapas de recuperación. Asimismo, el Índice de Simpson (D) y su complementario (1-D) corroboran esta baja diversidad, con valores consistentes entre Rec y CF, develando una estructura comunitaria con predominancia de pocas especies (Cuadro 5).
Cuadro 5 Número de especies, riqueza (Mg), diversidad (H′), Equitatividad (E), diversidad verdadera (D) y 1-D.
| Atributo | Etapas inventariadas | ||
|---|---|---|---|
| CI (2012) | Rec (2022) | CF (2022) | |
| Número de especies | 3 | 4 | 4 |
| Riqueza (Mg) | 0.37 | 0.52 | 0.57 |
| Diversidad (H′) | 0.62 | 0.66 | 0.67 |
| Equitatividad (E) | 0.56 | 0.48 | 0.48 |
| Diversidad verdadera (D) | 0.59 | 0.56 | 0.56 |
| 1-D | 0.41 | 0.44 | 0.44 |
Discusión
Las especies con el mayor IVI fueron Pinus arizonica y Pinus leiophylla, lo que sugiere que el género Pinus es el más importante; forma bosques con tendencia a masas puras, pues sus especies constituyen la mayor parte de la composición vegetal, aunque pueden coexistir con otros taxa en menor proporción. En esta misma región, Rascón-Solano et al. (2023) registraron a este género con la mayor importancia ecológica (70.81 %); Rascón-Solano et al. (2022) también calcularon valores superiores a 50 % para dicho género cerca del área de estudio. En contraste, García García et al. (2021) argumentaron que el género Pinus alcanza una importancia ecológica menor en los bosques templados del norte de México (30 %), de alto valor ecológico que se identifican con especies en estado de conservación.
La abundancia de estas especies puede estar asociada a la fisiografía en la zona, la precipitación media anual y el manejo forestal que favorece a las especies comerciales más notables.
Destaca Pinus arizonica como una especie representativa de los bosques templados del norte de México; llega a ser la principal de los ecosistemas de Pino y Pino-Encino (Hernández-Salas et al., 2018). Martínez-Salvador et al. (2013) la destacan como poseedora de un alto potencial productivo, ya que permite sostener la producción maderera con métodos selectivos e intervenciones silvícolas intensivas aplicadas para su desarrollo y aumento de la producción.
La presencia de Juniperus deppeana en la etapa de Reclutamiento y ausente en la etapa anterior, podría estar asociada a procesos de disturbio, dado su carácter oportunista y capacidad para colonizar áreas alteradas como especie primaria (Rubalcava-Castillo et al., 2020). Si bien su incorporación incrementa el número de especies registradas, su presencia no necesariamente refleja una mayor diversidad funcional o resiliencia del ecosistema, sino más bien una respuesta a las condiciones generadas tras las intervenciones silvícolas selectivas aplicadas (Rascón-Solano et al., 2023). Esto muestra que los tratamientos silvícolas pueden favorecer el establecimiento de especies secundarias, aunque aún sin producir una transformación sustancial en la composición general del bosque.
El DAP y la altura total de las especies arbóreas son variables predictoras del volumen de los bosques (Pacheco Escalona et al., 2007). En la distribución de individuos por clases diamétricas, el mayor número se agrupa entre las clases 15 y 35 cm en las tres etapas. A partir de diámetros de 40 cm y conforme esta variable aumenta, la estructura del bosque decrece en densidad, lo cual supone la presencia de algunos individuos jóvenes que en un futuro reemplazaran a los ejemplares de mayores dimensiones. Esto describe masas irregulares porque que se reflejan en curvas de distribución normal, lo que es similar a lo registrado por Delgado Zamora et al. (2016) para bosques templados de Durango. Si bien existen todas las categorías de altura, la mayor proporción de individuos se reúne entre los 18 y los 22 m. Dicho patrón estructural corresponde a una comunidad en transición hacia una estructura más madura y constata que los tratamientos aplicados permiten mantener un proceso de renovación natural, sin que se altere significativamente la distribución vertical y horizontal.
Como se esperaba, la especie con mayor producción en las tres etapas fue Pinus arizonica, la cual registró mayor abundancia y área basal. Algunos autores señalan que la producción en volumen aumenta con el incremento en la densidad de individuos (Návar-Cháidez & González-Elizondo, 2009); no obstante, es importante considerar que, a medida que la densidad crece, puede surgir una competencia significativa por los recursos luz, agua y nutrientes, lo que eventualmente podría limitar la producción. Lo anterior coincide con los resultados de este estudio, en los que la etapa de Reclutamiento, caracterizada por una alta densidad de individuos jóvenes, mostró el mayor número de individuos sin evidenciar aún signos de competencia intensa. Esto sugiere que, si bien el aprovechamiento ha permitido una regeneración densa, será necesario monitorear el crecimiento futuro para evitar efectos negativos por competencia intraespecífica.
La riqueza de especies de 0.37 a 0.57 sugiere baja diversidad, de acuerdo con los valores de referencia de Margalef (1977), lo que exhibe las condiciones ambientales del bosque analizado del norte de México, caracterizados por una baja diversidad y dominados por especies de pino y encino (Alfaro-Reyna et al., 2020). En estudios de bosques templados de Chihuahua, se han reportado valores mayores, como 1.52 por García García et al. (2020) y 1.23 por Hernández-Salas et al. (2013). Rascón-Solano et al. (2023) documentaron valores de 0.61 a 1.75 en tres etapas de manejo: un periodo inicial (2012), una etapa de recuperación tras tratamiento silvícola (2022) y una etapa de manejo de masas forestales (CF, 2022). En el ejido Aboreachi, Chihuahua, en un bosque de pino-encino conservado durante 30 años, se reportó un valor de 1.76, lo que indica que la riqueza puede incrementarse significativamente con la conservación a largo plazo (Rascón-Solano et al., 2022). Sin embargo, otros estudios señalan que el manejo forestal no siempre conlleva variación en la riqueza de especies a lo largo del tiempo (Hernández-Salas et al., 2013; Rascón-Solano et al., 2023). En este estudio, la estabilidad en la riqueza y diversidad sugiere que los tratamientos aplicados mantienen la composición sin generar efectos significativos a corto plazo, lo cual podría interpretarse como una señal de estabilidad estructural, pero también como una limitación para fomentar mayor complejidad ecológica.
Conclusiones
La composición del bosque se mantuvo relativamente estable antes y después de los tratamientos silvícolas, con una marcada dominancia de Pinus arizonica y Pinus leiophylla. Estas especies conservaron sus proporciones en las tres etapas de manejo forestal, lo que indica una estructura forestal centrada en estos taxa. La aparición de Juniperus deppeana en la etapa de Condición Final sugiere una ligera incorporación de especies secundarias, aunque sin afectar significativamente la composición general. Los índices de riqueza y diversidad registraron valores bajos a lo largo de las etapas evaluadas. Aunque la riqueza mejoró ligeramente con la incorporación de Juniperus deppeana, no se observaron cambios significativos en la Equitatividad, ni en la diversidad general del ecosistema. Esto indica que el aprovechamiento forestal actual no ha promovido una diversificación sustancial de especies a corto plazo.
La estructura horizontal se caracterizó por la predominancia de individuos en clases diamétricas medias (CF, 2022: 24.38 cm), mientras que verticalmente se observaron alturas mayores (CF, 2022: 19.5 m), lo que supone que el rodal está en transición hacia una estructura más madura. La distribución estable del volumen (m3 ha-1) entre etapas significa que no hubo un incremento importante en biomasa, reflejando una estructura estable, pero con baja complejidad. Esto sugiere que, si bien el manejo actual mantiene la producción y la estabilidad del bosque, aún no promueve de forma significativa una mayor diferenciación estructural.

