











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
En el ámbito del mejoramiento genético de especies forestales, los ensayos de procedencias y progenies tienen como objetivo principal obtener ganancias genéticas con base en la selección de los mejores fenotipos y genotipos (White et al., 2007). Estos ensayos inician con la selección de árboles superiores en función de su fenotipo; es decir, aquellos que presentan rendimiento sobresaliente en características evaluadas y de interés comercial en comparación con el promedio de la población natural (Zobel & Talbert, 1988).
La selección fenotípica permite elegir características de interés y predecir la ganancia genética para su posterior utilización en la formación de bosques más productivos y mejor adaptados (White et al., 2007). Estas características no se limitan únicamente al crecimiento rápido, sino que también abarcan aspectos relevantes como supervivencia, resistencia a factores ambientales adversos, calidad de la madera y producción de semillas (Flores-Flores et al., 2014). La cuantificación de la variabilidad del carácter, estimación de parámetros genéticos y heredabilidad por diferencial de selección son elementos clave en el proceso de mejoramiento genético de especies forestales (Flores-Flores et al., 2014). Además, proporcionan información valiosa para el manejo de huertos semilleros y guían a los programas de mejoramiento genético forestal (White et al., 2007).
La evaluación de la variación morfológica en ensayos de progenies es relevante para identificar individuos con características fenotípicas deseables y conocer su adaptabilidad en diferentes condiciones (Quijada, 1984; White et al., 2007). La interacción genotipo-ambiente resulta especialmente importante en especies con una amplia distribución natural como Cedrela odorata L., ya que las progenies o procedencias pueden comportarse de manera diferente en ambientes distintos (Flores-Flores et al., 2014). Por lo tanto, es deseable que los genotipos mantengan estabilidad en sus caracteres morfológicos en diferentes ambientes, ya que al tener un comportamiento relativamente bueno y previsible demuestran potencial para responder de manera favorable en diferentes sitios considerados para establecer plantaciones más productivas (Falconer & Mackay, 1996; Quijada, 1984; Zobel & Talbert, 1988). Sin embargo, obtener genotipos con estabilidad en diferentes ambientes es complicado. Por citar algún estudio, en ensayos de progenie establecidos en dos condiciones contrastantes, con más de 40 familias de Pinus elliottii Engelm. var. elliottii, solo se identificaron cuatro familias con alta estabilidad en caracteres morfológicos de interés, lo cual indica el nivel de adaptación de las familias a su ambiente y limita su potencial productivo a la condición del sitio de plantación (Pagliarini et al., 2016).
El comportamiento de las familias, en los ensayos de progenie, durante los primeros años refleja la variación de la expresión de diversas características, las cuales están influenciadas por la calidad del sitio, el manejo y la interacción con factores bióticos y abióticos (Rodríguez-Vásquez et al., 2021).
En áreas tropicales y subtropicales, C. odorata tiene un alto valor económico por la calidad de su madera (Olvera-Moreno et al., 2022). En México, ocupa el segundo lugar en términos de establecimiento de plantaciones forestales comerciales, después del género Eucalyptus spp., con una extensión de 20 705 ha (Velázquez Martínez et al., 2011). En conjunto, representan 60 % de la superficie plantada en el país, especialmente en los estados de Veracruz, Tabasco y Campeche (Palma-López et al., 2007). Por lo anterior, los objetivos del presente estudio fueron: (1) evaluar la variación en características morfológicas y de crecimiento entre familias establecidas en tres ensayos de progenie de C. odorata bajo diferentes condiciones ambientales, y (2) identificar las familias con mejor desempeño.
Materiales y Métodos
Los ensayos se establecieron en el año 2018, en las localidades Tamazunchale, en el estado de San Luis Potosí; Tihuatlán en Veracruz; y Venustiano Carranza en Puebla (Cuadro 1), que corresponden a lugares con características contrastantes entre sí (Instituto Nacional de Estadística, Geografía e Informática [INEGI], 2010a, 2010b, 2010c). En cada ensayo se consideraron 64 familias procedentes de Veracruz (23 familias), Puebla (17 familias), San Luis Potosí (11 familias), Tamaulipas (3 familias) e Hidalgo (10 familias).
Cuadro 1 Ubicación geográfica y características de los sitios de establecimiento de los ensayos de progenie de Cedrela odorata L.
| Ubicación | Latitud N y longitud O |
Altitud (msnm) |
Temperatura media anual (°C) |
Precipitación media anual (mm) |
Tipo
de suelo |
Exposición |
|---|---|---|---|---|---|---|
| Venustiano Carranza, Puebla |
20°28'26.3" 97°41'59.0" |
330 | 22-26 | 1 400 a 1 600 | Vertisol | Cenital |
| Tihuatlán, Veracruz |
20°33'44.9" 97°29'6.2" |
164 | 24-26 | 1 100 a 1 300 | Regosol | Cenital |
| Tamazunchale,
San Luis Potosí |
21°28'39"
98°58'45" |
140 | 20-24 | 1 500 a 3 000 | Leptosol | Cenital-norte |
Entre marzo y mayo de 2018 se establecieron los tres ensayos con plantas de las 64 familias de entre 6 y 8 meses de edad, con una altura de 30 a 40 cm y un diámetro al cuello de la raíz de 35 a 50 mm.
La planta se produjo en tubetes de plástico de 680 mL, llenados con una mezcla de Peat moss ( (60 %), vermiculita (20 %), perlita (20 %) y Osmocote ( 9-45-15 (3 g por cada 680 g de sustrato). El riego se aplicó cada tres o cuatro días, mediante un sistema de riego robotizado, con fertilización programada una vez por semana con fertilizante marca Peters ( 20-10-20 (3 g L-1). Se siguió un diseño experimental de bloques completos al azar (BCA), con una planta por familia por bloque como unidad experimental, a un espaciamiento de 4×4 m en 16 bloques. Por lo tanto, en cada bloque se incorporaron 64 plantas y por ensayo, un total de 1 024 ejemplares.
Variables evaluadas
Al momento de la evaluación, solo se consideraron 57 de las 64 familias en el análisis, ya que siete se descartaron por presentar 100 % de árboles muertos.
En cada ensayo se determinó el porcentaje de supervivencia (SUP) (vivo: árboles en pie con hojas verdes y brotes en crecimiento en las copas; muerto: árboles sin tejidos vivos en pie o derribados con ausencia de hojas y brotes en las copas), la altura total (ALT; m) (pértiga telescópica±1 cm, sin marca registrada), altura de fuste limpio (AFL; cm) (pértiga telescópica±1 cm, sin marca registrada) y diámetro normal (DN; mm) (vernier digital±0.05 mm, Mitutoyo ® modelo 573-282). La forma del fuste (FOR) se determinó con base en cuatro categorías: (1) Muy torcido, (2) Torcido, (3) Ligeramente torcido, y (4) Recto; bifurcaciones (BIF): (1) Bifurcada desde un tercio inferior, (2) Bifurcada desde un tercio medio, (3) Bifurcada solo en un tercio superior, y (4) No bifurcada; e incidencia de ataque por Hypsipyla grandella (Zeller) (NIV) con base en la cantidad de perforaciones en el árbol: (1) Alta: en todo el fuste, brotes y ramas, (2) Media: en brotes y ramas, y (3) Baja: solo en brotes.
Análisis estadístico
La verificación de los supuestos de normalidad y homogeneidad de varianzas se determinaron mediante la prueba Shapiro-Wilk con el procedimiento UNIVARIATE de SAS © versión 9.1 (SAS Institute Inc., 2004) para las variables SUP (p=0.9342), ALT (p=0.9556) y DN (p=0.9678). Se realizó un análisis de varianza (α<0.05) con el procedimiento GLM de SAS © (SAS Institute Inc., 2004). El modelo estadístico utilizado fue:
Donde:
Y ijk = Valor observado en el individuo de la k-ésima familia, en el j-ésimo bloque anidado en el i-ésimo sitio
µ = Media general
S i = Efecto fijo del i-ésimo sitio
β j(i) = Efecto fijo del j-ésimo bloque anidado en el i-ésimo sitio
F k = Efecto aleatorio de la k-ésima familia ~NID (0, σ 2 f )
SF ik = Efecto aleatorio de la interacción sitio×familia ~NID (0, σ 2 sf )
е ijk = Error experimental ~NID (0, σ 2 e )
i = 1, 2 y 3 (sitios)
j = 1, …16 (bloques)
k = 1, …57 (familias)
Posteriormente, se llevó a cabo la separación de medias mediante la prueba de Tukey (α<0.05) (SAS Institute Inc., 2004). La clasificación de las familias se hizo mediante el procedimiento sugerido por Dlamini et al. (2017) y Edward et al. (2014) y a cada variable evaluada (SUP, ALT, AFL, DN, FOR, BIF y NIV) se le asignó un valor categórico continuo, los cuales variaron desde el mejor (con 1 punto asignado) y sucesivamente hasta el peor (con 57 puntos asignados) por cada familia. Entonces, los datos por variable fueron promediados para obtener el promedio general (MEDIA) de las siete variables en cada familia. Posteriormente, las familias se agruparon y clasificaron (CLAS) con base en los promedios (MEDIA), de manera descendente, usando el procedimiento PROC RANK de SAS © (SAS Institute Inc., 2004).
Resultados y Discusión
El factor sitio influyó significativamente (p<0.0001) sobre la supervivencia, altura y diámetro; sin embargo, no se obtuvo significancia entre familias, ni en la interacción entre ambos factores (Cuadro 2).
Cuadro 2 Valores de probabilidad (P) obtenidos en el análisis de varianza para las variables de supervivencia, altura y diámetro de familias de Cedrela odorata L.
| FV | GL | Supervivencia | ALT | DN |
|---|---|---|---|---|
| Sitio | 2 | <0.0001 | <0.0001 | <0.0001 |
| Familia | 56 | 0.3297 | 0.6775 | 0.9572 |
| Sitio×Familia | 112 | 0.1106 | 0.9364 | 0.9897 |
FV = Fuente de variación; GL = Grados de libertad; ALT = Altura total; DN = Diámetro normal.
Efecto del sitio en la supervivencia
El porcentaje promedio de supervivencia entre los tres ensayos fue superior a 60 %. Los árboles de los ensayos de Venustiano Carranza y Tamazunchale tuvieron mayor supervivencia (>70 %) que los de Tihuatlán (35 %) (Figura 1). En estudios anteriores se ha observado que la mortalidad en las plantaciones está, principalmente, asociada con las condiciones ambientales y prácticas de silvicultura de cada sitio, así como con la falta de protección adecuada (Grignola et al., 2014).
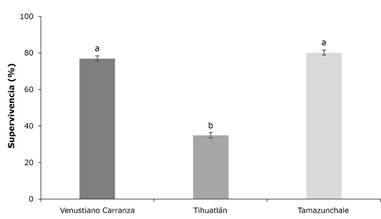
Medias con letra diferente son estadísticamente diferentes con p≤0.05. Error estándar representado en las líneas ubicadas por encima de las barras.
Figura 1 Efecto del sitio en los porcentajes de supervivencia de los árboles de Cedrela odorta L.
En el ensayo de Tihuatlán se registró una supervivencia de 35 % debido a las condiciones del terreno, en donde se presentaron problemas de anegamiento excesivo del suelo, lo que provocó hipoxia y anoxia que afectan negativamente las funciones de la planta, incluida la fotosíntesis (Lee et al., 2011; Peña-Castro, 2014). Otros factores asociados fueron la competencia con malezas y la presencia de plantas trepadoras, las cuales suprimieron el crecimiento de los árboles de C. odorata, los cuales una vez debilitados fueron más susceptibles al ataque de plagas y enfermedades (Da Ponte Canova & Salas, 2009).
Efecto del sitio en altura y diámetro
Los árboles establecidos en Tamazunchale alcanzaron alturas significativamente mayores que los de Venustiano Carranza y Tihuatlán, con una diferencia de aproximadamente 2.3 y 3.3 m, respectivamente (Figura 2A). En cuanto al DN, los árboles en el ensayo de Tamazunchale exhibieron diámetros superiores a 5.5 cm, y superaron en más de 2.8 y 4.6 cm a los árboles de Venustiano Carranza y Tihuatlán, respectivamente (Figura 2B).
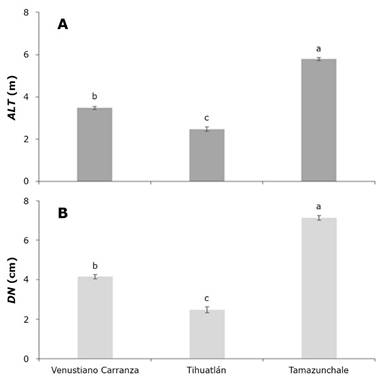
A = Altura total (ALT; m); B = Diámetro normal (DN; cm). Medias con letra diferente son estadísticamente diferentes con α≤0.05. Las líneas ubicadas por encima de las barras representan el error estándar.
Figura 2 Efecto del sitio sobre la altura total y el diámetro normal de árboles de Cedrela odorata L.
Las condiciones de menor altitud, mayor precipitación anual, suelo con buen drenaje (Leptosol) y la exposición norte en Tamazunchale favorecieron el crecimiento en altura (ALT) y diámetro (DN) de los árboles, en contraste con los de Venustiano Carranza y Tihuatlán. En condiciones óptimas de precipitación (1 200 a 3 000 mm), la especie presenta altas tasas de crecimiento en altura (Alonso-Báez et al., 2020).
El crecimiento de los árboles y su adaptación a las condiciones ambientales son aspectos importantes para comprender tanto su respuesta al clima, como las estrategias de adaptación genética (Mondino, 2014). Además, en este mismo ensayo se realizaron labores culturales en lo relativo al control de malezas y podas, a diferencia de los otros dos ensayos. Es recomendable llevar a cabo prácticas regulares de control de malezas para evitar la competencia entre individuos durante los primeros tres años de establecimiento de la plantación (Toro Vergara, 2004).
La presencia del barrenador Hypsipyla grandella representa un riesgo significativo para las plantaciones, ya que afecta el crecimiento de los árboles y la calidad de la madera (Cornelius & Watt, 2003; Santos Murgas et al., 2015). El monitoreo regular y la implementación de medidas de control como las prácticas culturales son fundamentales para mitigar este problema. En particular, en Tihuatlán, con menor precipitación, prácticas culturales escasas y algunas zonas de la plantación con drenaje deficiente, que permiten inundaciones frecuentes durante la temporada de lluvias, generaron condiciones estresantes para los árboles. Aunque C. odorata puede crecer en diferentes ambientes, se sabe que las características del suelo determinan su establecimiento, crecimiento y desarrollo; los mejores crecimientos de la especie se han registrado en suelos con buen drenaje y fertilidad (Cintron, 1990).
Clasificación ordinal de familias
En la clasificación general, 39 % de las familias están dentro de los primeros cinco lugares (Cuadro 3). Esto indica una respuesta importante y positiva de la mayoría de las familias a los tres sitios; a pesar de que la selección fenotípica de los árboles superiores se realizó con los residuos de las poblaciones naturales de C. odorata, en ocasiones con densidades de árboles reducidas y en otros casos, solo con relictos donde no había muchos individuos para hacer una mejor selección.
Cuadro 3 Clasificación ordinal de las familias en los tres ensayos de progenie de Cedrela odorata L.
| Familia | SUP | ALT | AFL | DN | FOR | BIF | NIV | MEDIA | CLAS |
|---|---|---|---|---|---|---|---|---|---|
| 62 | 11 | 2 | 9 | 3 | 11 | 7 | 7 | 7 | 1 |
| 7 | 21 | 4 | 5 | 6 | 5 | 5 | 5 | 7 | 1 |
| 45 | 27 | 1 | 6 | 2 | 1 | 9 | 6 | 7 | 1 |
| 20 | 24 | 4 | 3 | 8 | 5 | 5 | 5 | 8 | 2 |
| 37 | 14 | 5 | 10 | 6 | 7 | 5 | 9 | 8 | 2 |
| 71 | 39 | 1 | 4 | 2 | 6 | 5 | 2 | 8 | 2 |
| 26 | 10 | 3 | 19 | 5 | 11 | 6 | 5 | 8 | 2 |
| 48 | 3 | 6 | 21 | 6 | 8 | 6 | 9 | 8 | 2 |
| 18 | 25 | 6 | 1 | 7 | 15 | 1 | 9 | 9 | 3 |
| 51 | 19 | 6 | 12 | 7 | 9 | 6 | 5 | 9 | 3 |
| 63 | 35 | 7 | 7 | 10 | 2 | 2 | 2 | 9 | 3 |
| 34 | 1 | 6 | 14 | 10 | 18 | 5 | 11 | 9 | 3 |
| 77 | 29 | 3 | 17 | 3 | 2 | 4 | 9 | 10 | 4 |
| 59 | 3 | 8 | 31 | 8 | 8 | 3 | 7 | 10 | 4 |
| 65 | 23 | 8 | 8 | 10 | 9 | 5 | 5 | 10 | 4 |
| 72 | 17 | 7 | 29 | 6 | 5 | 2 | 3 | 10 | 4 |
| 80 | 25 | 7 | 11 | 9 | 9 | 3 | 9 | 10 | 4 |
| 8 | 46 | 5 | 2 | 4 | 10 | 4 | 4 | 11 | 5 |
| 10 | 6 | 6 | 25 | 13 | 9 | 6 | 11 | 11 | 5 |
| 3 | 2 | 8 | 35 | 13 | 7 | 8 | 5 | 11 | 5 |
| 61 | 16 | 7 | 28 | 8 | 9 | 1 | 9 | 11 | 5 |
| 55 | 25 | 6 | 15 | 9 | 12 | 3 | 9 | 11 | 5 |
| 33 | 13 | 8 | 20 | 10 | 16 | 7 | 9 | 12 | 6 |
| 78 | 4 | 9 | 33 | 14 | 16 | 5 | 4 | 12 | 6 |
| 19 | 18 | 4 | 36 | 10 | 10 | 7 | 1 | 12 | 6 |
| 46 | 20 | 7 | 22 | 8 | 10 | 8 | 11 | 12 | 6 |
| 29 | 9 | 5 | 39 | 8 | 8 | 9 | 10 | 13 | 7 |
| 54 | 8 | 7 | 40 | 12 | 10 | 9 | 2 | 13 | 7 |
| 13 | 7 | 8 | 45 | 12 | 7 | 5 | 5 | 13 | 7 |
| 5 | 15 | 8 | 32 | 10 | 9 | 4 | 11 | 13 | 7 |
| 12 | 22 | 8 | 23 | 14 | 9 | 4 | 11 | 13 | 7 |
| 42 | 35 | 7 | 13 | 9 | 10 | 10 | 8 | 13 | 7 |
| 36 | 38 | 5 | 26 | 1 | 14 | 5 | 7 | 14 | 8 |
| 58 | 26 | 11 | 26 | 14 | 13 | 3 | 5 | 14 | 8 |
| 69 | 44 | 9 | 18 | 9 | 12 | 3 | 4 | 14 | 8 |
| 23 | 35 | 11 | 16 | 16 | 11 | 1 | 11 | 14 | 8 |
| 60 | 5 | 6 | 52 | 7 | 13 | 7 | 11 | 14 | 8 |
| 39 | 30 | 7 | 30 | 13 | 7 | 4 | 11 | 15 | 9 |
| 76 | 43 | 5 | 27 | 6 | 13 | 4 | 4 | 15 | 9 |
| 70 | 35 | 10 | 24 | 13 | 8 | 7 | 6 | 15 | 9 |
| 66 | 12 | 9 | 56 | 12 | 8 | 11 | 5 | 16 | 10 |
| 35 | 28 | 5 | 42 | 11 | 11 | 7 | 11 | 16 | 10 |
| 28 | 39 | 9 | 34 | 7 | 17 | 6 | 4 | 17 | 11 |
| 43 | 34 | 9 | 49 | 11 | 7 | 6 | 4 | 17 | 11 |
| 79 | 25 | 10 | 46 | 15 | 12 | 8 | 4 | 17 | 11 |
| 68 | 37 | 9 | 41 | 11 | 12 | 9 | 5 | 18 | 12 |
| 22 | 33 | 10 | 44 | 11 | 14 | 7 | 7 | 18 | 12 |
| 11 | 40 | 10 | 43 | 12 | 11 | 4 | 8 | 18 | 12 |
| 47 | 41 | 9 | 38 | 15 | 11 | 8 | 7 | 18 | 12 |
| 21 | 49 | 6 | 47 | 10 | 3 | 6 | 10 | 19 | 13 |
| 49 | 31 | 10 | 48 | 17 | 9 | 6 | 11 | 19 | 13 |
| 32 | 42 | 9 | 51 | 14 | 6 | 7 | 5 | 19 | 13 |
| 64 | 32 | 9 | 50 | 13 | 15 | 10 | 5 | 19 | 13 |
| 50 | 47 | 5 | 55 | 4 | 12 | 8 | 4 | 19 | 13 |
| 41 | 45 | 7 | 37 | 13 | 18 | 11 | 4 | 19 | 13 |
| 25 | 48 | 4 | 54 | 8 | 4 | 7 | 11 | 19 | 13 |
| 24 | 36 | 9 | 53 | 16 | 5 | 8 | 11 | 20 | 14 |
SUP = Supervivencia; ALT = Altura total; AFL = Altura de fuste limpio; DN = Diámetro normal; FOR = Forma de fuste; BIF = Bifurcaciones; NIV = Nivel de incidencia de Hypsipyla grandella; MEDIA = Promedio de los siete atributos; CLAS = Clasificación. La sombra indica las 22 mejores familias (arriba) y las 17 peores (abajo), sin sombras familias intermedias.
Los resultados revelaron que las familias 7, 45 y 62 exhibieron una notable estabilidad en los tres sitios analizados, principalmente en Venustiano Carranza y Tamazunchale. La familia 45 se mantuvo dentro de los primeros cinco lugares, y mostró consistencia en cada uno de los tres sitios; destacaron en ALT, DN y FOR (con las primeras posiciones 1, 2, 1, respectivamente) que corresponden a las variables de mayor interés comercial. También resalta la familia 62, la cual sobresalió en ALT y DN (posiciones 2 y 3, respectivamente), y conservó una posición relevante en ambos lugares. La familia 7 exhibió la mayor consistencia en las características de crecimiento, bifurcación, forma de fuste e incidencia de Hypsipyla grandella, siempre entre las posiciones 4 a 6 (Cuadro 3).
Las características destacadas de estas tres familias en los sitios evaluados indican su capacidad de respuesta a las diferentes condiciones presentes en cada uno de ellos. Sin embargo, es necesario complementar estos resultados con la estimación de parámetros genéticos y la evaluación de interacción genotipo-ambiente. En estudios similares de ensayos de C. odorata, pero con planta injertada, se indicó que la heterogeneidad ambiental entre sitios fue la causa principal de la baja heredabilidad en variables de crecimiento, además de que la interacción genotipo-ambiente resultó de baja a moderada (Olvera-Moreno et al., 2022).
Estos parámetros ayudan a recomendar planes de manejo en los ensayos y a seleccionar genotipos superiores con fines de mejoramiento genético de la especie en cada uno de los sitios. El desarrollo y crecimiento de los genotipos está relacionado con su genética y todas las condiciones ambientales presentes de cada ensayo, particularmente los factores edáficos, climáticos y bióticos (Kimmins, 2004). Por lo tanto, la variabilidad observada representa la diversidad de genotipos presentes y es un parámetro fundamental para los programas de mejoramiento genético (Murillo et al., 2017). Esta variabilidad se manifiesta a nivel morfológico en los árboles, afecta características como la altura, diámetro, rectitud y calidad del fuste.
En México, con estudios como el aquí documentado, se están logrando avances significativos en el campo del mejoramiento genético de C. odorata, con el objetivo de desarrollar materiales mejorados que presenten características estables y distintivas en diferentes ubicaciones (Rodríguez-Vásquez et al., 2021). En el presente estudio, las evaluaciones realizadas en los tres sitios con condiciones ambientales contrastantes permiten conocer el comportamiento y variación en características de crecimiento y morfológicas del fuste en las familias. Lo anterior contribuirá a determinar la adaptabilidad y estabilidad del material plantado (Olvera Moreno et al., 2022); pero estudios de variación genética serán necesarios para realizar los aclareos genéticos en los ensayos y su transformación a huertos semilleros.
En el presente estudio, a los tres años de edad de establecidas las plantaciones, a nivel de familias, los árboles de C. odorata no exhibieron una marcada diferenciación morfológica.
Conclusiones
A los tres años de establecidos los ensayos, la variación morfológica esperada entre las familias de C. odorata no se verificó, a pesar de la diversidad de condiciones de su origen en Veracruz, Puebla, San Luis Potosí, Tamaulipas e Hidalgo. Sin embargo, el factor sitio influyó en la supervivencia y caracteres de crecimiento de los árboles. En particular, los árboles establecidos en Tamazunchale, San Luis Potosí destacaron en relación con los otros dos ensayos por su alta supervivencia y características comerciales, como altura y diámetro normal; el sitio presenta una precipitación media anual muy superior al de los otros dos, y tiene un suelo tipo Leptosol, diferente al Vertisol y al Regosol existentes en Venustiano Carranza y Tihuatlán, respectivamente; ambos son factores que inciden en la variación descrita. Finalmente, las familias 7, 45 y 62 se identifican como las de mejor desarrollo y crecimiento, de acuerdo a la clasificación basada en los atributos morfológicos considerados en este estudio.
La continuidad de la evaluación será importante para corroborar los resultados en el mediano y largo plazo. Además, se recomienda estudiar y analizar el grado de control genético presente en los caracteres morfológicos, con la finalidad de conocer su relevancia en la adaptación y domesticación de las familias en diferentes ambientes.

