Artículo científico
Aboveground and root biomass in sapling stage of Abies religiosa (Kunth) Schltdl. & Cham. in Hidalgo state
Manzur Chávez, Nancy1
 http://orcid.org/0000-0001-9549-3079
http://orcid.org/0000-0001-9549-3079
Rodríguez Laguna, Rodrigo2*
Razo Zárate, Ramón2
http://orcid.org/0000-0002-4608-3361
Acevedo Sandoval, Otilio Arturo1
http://orcid.org/0000-0003-0475-7003
Octavio Aguilar, Pablo1
http://orcid.org/0000-0002-4636-9773
http://orcid.org/0000-0001-9549-3079Rodríguez Laguna, Rodrigo2*
Razo Zárate, Ramón2
http://orcid.org/0000-0002-4608-3361Acevedo Sandoval, Otilio Arturo1
http://orcid.org/0000-0003-0475-7003Octavio Aguilar, Pablo1
http://orcid.org/0000-0002-4636-9773
-
Publication dates-
May 26, 2025
Jan-Feb , 2025
- Article in PDF
- Article in XML
- Automatic translation
- Send this article by e-mail
- Share this article +
Abstract
The initial development of the forest is known as sapling. At this stage the regeneration density is high, so the biomass accumulation rate is higher. In this context, the objective of estimating the total biomass storage capacity (aboveground-root) in trees in the initial stage of Abies religiosa in El Chico National Park, Hidalgo state, was raised. Four sampling sites of 100 m2 were established to obtain population density. 52 specimens less than 1.5 m in height and 5 cm in average base diameter were extracted with roots; a high-pressure water backpack sprayer was used to detach the soil from the root; later the plant material was placed in a drying oven at 80 °C until reaching constant weight. Results indicate that the fir trees in the sapling stage formed the basic group (height 6 to 65 cm) with 72.6 % of aboveground biomass and 27.4 % of root biomass, and another developed group (height 68 to 150 cm), with 75.8 and 24.2 % of aboveground and root biomass, respectively. The biomass stored in the fir forest in the sapling stage was 103.6 kg ha-1, with potential accumulation of aboveground/root biomass in a 3:1 ratio in canopy gaps, which favored the establishment of natural regeneration. The ratio of root biomass to aboveground biomass in the sapling stage changes with age in Abies religiosa trees.
Key words:
Natural Protected Area, aboveground biomass, root biomass, sapling, fir, natural regeneration
Introduction
Plant biomass is formed by all the organic components within an ecosystem (Litton et al., 2007) and is important as a source of energy, as well as in the storage of carbon (C) and nitrogen (N) (Peichl et al., 2012; Schuler et al., 2017). The capacity of a forest to capture atmospheric carbon tends to decrease with the increase in the age of the trees. It is known that at early or intermediate ages the rate of carbon capture is higher (López-Reyes et al., 2016). In addition, it is related to the rate of accumulation of aboveground and root biomass that forests have with the net growth of trees that are capable of sequestering more CO2 than they emit through respiration. The rate of carbon capture is directly proportional to said growth (Casiano-Domínguez et al., 2018).
-
Litton et al., 2007Carbon allocation in forest ecosystemsGlobal Change Biology, 2007
-
Peichl et al., 2012Above-and belowground ecosystem biomass, carbon and nitrogen allocation in recently afforested grassland and adjacent intensively managed grasslandPlant and Soil, 2012
-
Schuler et al., 2017Biomass estimates of small diameter planted and natural-origin loblolly pines show major departures from the national biomass estimator equationsForest Science, 2017
-
López-Reyes et al.,
2016Carbono almacenado en la biomasa aérea de plantaciones de hule (Hevea brasiliensis Müell. Arg.) de diferentes edadesMadera y Bosques, 2016
-
Casiano-Domínguez et al., 2018El Carbono de la biomasa aérea medido en cronosecuencias: primera estimación en MéxicoMadera y Bosques, 2018
Natural Protected Areas (NPA) are intended to conserve, protect and recover natural resources (González et al., 2014; Íñiguez et al., 2014). Such is the case of El Chico National Park, which is a provider of important environmental services for the mountainous region of central Mexico and is home to one of the most important relics of Abies religiosa (Kunth) Schltdl. & Cham. (fir) (Comisión Nacional de Áreas Naturales Protegidas [Conanp], 2005). This species develops in humid conditions (precipitation greater than 1 000 mm) and in low temperatures (average temperature of 7 to 12 °C), so the increase in the temperature on the planet causes effects, which are manifested in modifications at the molecular, morphological and physiological level that are reflected in negative alterations in its early stages of development (Romahn-Hernández et al., 2020).
-
González et al., 2014Las áreas naturales protegidas de MéxicoInvestigación y Ciencia de la Universidad Autónoma de Aguascalientes, 2014
-
Comisión Nacional de Áreas Naturales
Protegidas [Conanp], 2005Programa de conservación y manejo Parque Nacional El Chico, 2005
-
Romahn-Hernández et al., 2020Rango altitudinal: factor de vigor forestal y determinante en la regeneración natural del oyamelRevista Entreciencias: Diálogos en la Sociedad del Conocimiento, 2020
The initial development of the temperate forest is known as the sapling stage, which includes trees up to approximately 1.5 m tall and with an average base diameter of less than 5 cm (Aguilar, 2018; Hutchinson, 1993; Ronquillo-Gorgúa et al., 2022). In most temperate climate NPAs, the sapling stage is established during the first rainy period, after gaps are generated in the canopy that allow solar radiation to enter the forest floor (Lara-González et al., 2009). Generally, there are densities of up to 6 100 seedlings ha-1 in western exposures (Rodríguez-Laguna et al., 2015) that are reduced over time due to competition for abiotic and biotic factors that thus achieve a gradual accumulation of biomass in the aboveground and root parts of the tree (Bar-On et al., 2018; Ronquillo-Gorgúa et al., 2022).
-
Aguilar, 2018Estructura y diversidad de la vegetación arbórea de un bosque de galería en el estado de PueblaRevista Mexicana de Ciencias Forestales, 2018
-
Hutchinson, 1993Puntos de partida y muestreo diagnóstico para la silvicultura de bosques naturales del trópico húmedo, 1993
-
Ronquillo-Gorgúa et al., 2022Carbon storage during the development stages of Pinus patula Schiede ex Schltdl. & Cham. in the Sierra Alta of HidalgoRevista Chapingo Serie Ciencias Forestales y del Ambiente, 2022
-
Lara-González et al.,
2009Regeneration of Abies religiosa in canopy gaps versus understory, Cofre de Perote National Park, MéxicoAgrociencia, 2009
-
Rodríguez-Laguna et al., 2015Regeneración natural post-incendio de Abies religiosa (H. B. K.) Schl. et Cham, en el Parque Nacional “El Chico” HidalgoRevista Iberoamericana de Ciencias, 2015
-
Bar-On et al., 2018The biomass distribution on EarthProceedings of the National Academy of Sciences, 2018
-
Ronquillo-Gorgúa et al., 2022Carbon storage during the development stages of Pinus patula Schiede ex Schltdl. & Cham. in the Sierra Alta of HidalgoRevista Chapingo Serie Ciencias Forestales y del Ambiente, 2022
Studies focused on estimating the accumulation of aboveground biomass in forests (Albers et al., 2019; Oliveira et al., 2018; Pham et al., 2019; Razo et al., 2015), do not consider root biomass (Adame et al., 2017; Addo-Danso et al., 2016; Sochacki et al., 2017). Recent research suggests the need to know point estimates by component of the total forest biomass on a large scale (Bar-On et al., 2018; Djomo et al., 2011; Fu et al., 2017).
-
Albers et al., 2019Data and non-linear models for the estimation of biomass growth and carbon fixation in managed forestsData in Brief, 2019
-
Oliveira et al., 2018Above-and below-ground carbon accumulation and biomass allocation in poplar short rotation plantations under Mediterranean conditionsForest Ecology and Management, 2018
-
Pham et al., 2019Integrating Sentinel-1A SAR data and GIS to estimate aboveground biomass and carbon accumulation for tropical forest types in Thuan Chau district, VietnamRemote Sensing Applications: Society and Environment, 2019
-
Razo et al., 2015Coeficientes de Carbono para arbustos y herbáceas del bosque de oyamel del Parque Nacional El ChicoRevista Mexicana de Ciencias Forestales, 2015
-
Adame et al., 2017Mangrove root biomass and the uncertainty of belowground carbon estimationsForest Ecology and Management, 2017
-
Addo-Danso et al., 2016Methods for estimating root biomass and production in forest and woodland ecosystem carbon studies: A reviewForest Ecology and Management, 2016
-
Sochacki et al., 2017Accuracy of tree root biomass sampling methodologies for carbon mitigation projectsEcological Engineering, 2017
-
Bar-On et al., 2018The biomass distribution on EarthProceedings of the National Academy of Sciences, 2018
-
Djomo et al., 2011Estimations of total ecosystem carbon pools distribution and carbon biomass current annual increment of a moist tropical forestForest Ecology and Management, 2011
-
Fu et
al., 2017Individual tree biomass models to estimate forest biomass for large spatial regions developed using four pine species in ChinaForest Science, 2017
The machinery, tools and time to extract adult trees with roots is almost impossible, but the interest in providing information on the root component of young trees allows for complementary information to reach the total biomass stored in the forest. Fragoso-López et al. (2017) estimated the aboveground biomass component through geographic information systems; likewise, the biomass stored on the soil surface, known as necromass (Cortés-Blobaum et al., 2019), is known, so it is necessary to incorporate a part of the root component in this ecosystem. Therefore, the objective of the present research was to estimate the total biomass accumulation capacity (aboveground-root) in A. religiosa trees at the sapling stage in El Chico National Park, Hidalgo state, Mexico.
-
Fragoso-López et al.
(2017)Carbon sequestration in protected areas: A case study of an Abies religiosa (H. B. K.) Schlecht. et Cham forestForests, 2017
-
Cortés-Blobaum et al., 2019Patrones culturales de uso de leña en la primera área protegida de Latinoamérica, El Chico, MéxicoRevista Iberoamericana de Ciencias, 2019
Materials and Methods
Study area
The study was carried out in the NPA El Chico National Park, located at the western end of the Sierra de Pachuca, Hidalgo state, in the Transversal Neovolcanic Axis (Figure 1). It is located between 20°10’10’’ to 20°13’25’’ North and 98°41’50’’ to 98°46’02’’ West, with an area of 2 739 ha, and altitudes between 2 600 and 3 050 m (Conanp, 2005).
Thumbnail
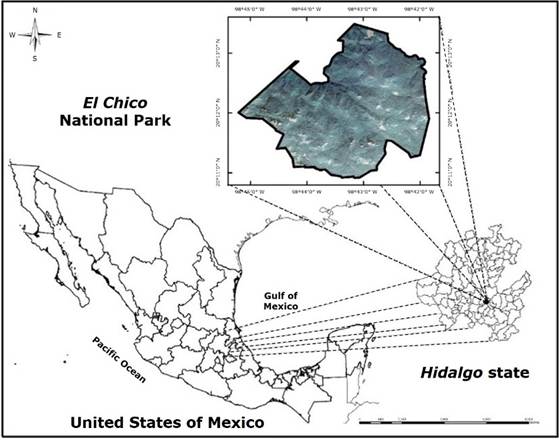
Figure 1
Geographic location of El Chico National Park, Hidalgo state, Mexico.
Geographic location of El Chico National Park, Hidalgo state, Mexico.
The climate is temperate-subhumid with summer rains (C(m)(w)b(i0)gw) and mean annual temperature between 12 and 18 °C. The predominant soils are humic Cambisol, dystric Regosol and medium-textured humic Andosol (Conanp, 2005). The largest percentage of vegetation cover (67 %) belongs to the A. religiosa forest (Fragoso-López et al., 2017); other important tree species are: Quercus spp., Pseudotsuga macrolepis Flous, Taxus globosa Schltdl. and Pinus spp. (Conanp, 2005).
-
Fragoso-López
et al., 2017Carbon sequestration in protected areas: A case study of an Abies religiosa (H. B. K.) Schlecht. et Cham forestForests, 2017
Tree density in the sapling stage
To determine density, four 100 m2 (10×10 m) plots were established in fir regeneration areas; from which the complete saplings with roots were extracted.
Tree selection for aboveground-root biomass analysis
Travels were conducted within El Chico National Park to identify spaces (clearings) within the forest that were large enough for natural fir regeneration. A total of 52 A. religiosa trees at sapling stage were selected, with average heights of 6-150 cm (Ronquillo-Gorgúa et al., 2022), free of mechanical damage, far from roads or gaps with compacted soil, and were completely extracted from the root.
-
Ronquillo-Gorgúa et al., 2022Carbon storage during the development stages of Pinus patula Schiede ex Schltdl. & Cham. in the Sierra Alta of HidalgoRevista Chapingo Serie Ciencias Forestales y del Ambiente, 2022
Organic matter was removed from each tree using a rake, then sufficient water was gradually added around the base of the specimen in order to soften the soil and remove it the next day, taking care not to damage the root. A high-pressure water backpack sprayer was used and the direction and depth of each root was followed (one at a time) until the complete extraction of the root system was ensured. A wooden support in the form of a square was implemented, fixed to the ground, which helped to keep the tree in a vertical position; in the middle part of the trunk they were tied with plastic ropes during the entire extraction process, until the root system was completely free.
Evaluated variables
The total height (cm) from the soil surface to the apex of each tree was measured with a model FCN-3M Truper® tape measure; the base diameter (cm), with a model 14388 Truper® digital vernier with millimetric precision; age was determined by the number of whorls in each individual, since these are produced at a rate of one whorl per year in Abies (Lara-González et al., 2009). The complete root system of each specimen was washed and left to air dry in the shade for approximately two hours (Fonseca et al., 2009) and the fresh weight of the entire tree (g) was taken in the field with a model BE16001 Biobase® scale. Subsequently, the aerial and root parts were separated to weigh each component and were placed each in paper bags previously labeled with the sample number and were placed in a model LW-201C GRIEVE® drying oven at 80 °C until reaching constant weight. With this, the average dry weight (g) of each component was calculated and multiplied by the number of fir plants per hectare.
-
Lara-González et
al., 2009Regeneration of Abies religiosa in canopy gaps versus understory, Cofre de Perote National Park, MéxicoAgrociencia, 2009
-
Fonseca et al., 2009Modelos para estimar la biomasa de especies nativas en plantaciones y bosques secundarios en la zona Caribe de Costa RicaBosque, 2009
In a first analysis, the variance of the dry weight of the root was determined with the following formula:
e2
n
=
(
t
2
σ
2
)
d
2
Where:
t = Standardized value of the error at 5 %, which is equivalent to 1.96
σ 2 = Variance of the attribute to be evaluated
d = Confidence interval for the quality measured (Zar, 2010).
-
Zar, 2010Biostatistical analysis, 2010
Data analysis
With the information from the 52 trees included in the sample, a cluster analysis was performed based on the variables height, dry weight of the whole tree, dry weight of the root and percentage of root biomass, using the Ward method based on Euclidean distances; for this, the Statistica v.10 program was used (StatSoft Inc., 2011). The dry root biomass was contrasted using a t-test for unbalanced samples, before a normality test. For this analysis, the Past program was used (Hammer et al., 2001). And the rest of the attributes were contrasted with parametric or non-parametric paired tests according to the type of data.
-
StatSoft Inc.,
2011STATISTICA (data analysis software system), 2011
-
Hammer et al.,
2001PAST: Paleontological statistics software package for education and data analysisPalaeontologia electrónica, 2001
Results and Discussion
The cluster analysis indicated the formation of two groups that are mainly distinguished by smaller size (initial sapling) with a height range of 6 to 65 cm, 0.07 to 0.9 cm base diameter and age of 1 to 8 years, and another of larger size (developed sapling) with a height range of 68 to 150 cm, 0.9 to 2.1 cm base diameter and age of 8 to 12 years. The root biomass data are normally distributed (Anderson-Darling, 0.4669 and 0.3743; p>0.05) with 38 individuals of initial sapling and 14 of developed sapling; on the other hand, no significant differences were found between groups of saplings (t=1.7468; p=0.09), while for height: t=5.47 and p=0.0001, dry weight of the aerial part: t=5.435 and p=0.0001, and dry weight of the root: t=5.482 and p=0.0001.
The organization of two groups is possibly due to the age of the specimens and the differences in the formed and accumulated biomass (Figure 2), in addition to the changes in the formation of roots at early ages, so it is not possible to assume a linear increase throughout the life of the plant.
Thumbnail
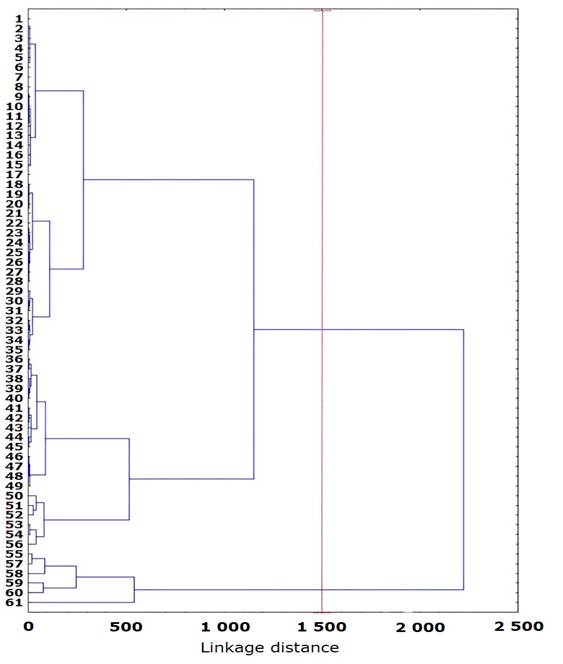
Figure 2
Grouping by morphological attributes in Abies religiosa (Kunth) Schltdl. & Cham. plants at the sapling stage in El Chico National Park, Hidalgo state, Mexico.
Grouping by morphological attributes in Abies religiosa (Kunth) Schltdl. & Cham. plants at the sapling stage in El Chico National Park, Hidalgo state, Mexico.
Some works mention that the proportion of aboveground biomass-root biomass are interdependent, which highlights the importance of the functional balance of both parts (van Noordwijk & Willigen, 1987). However, the growth rate of both elements differs depending on soil water content, biophysical processes, changes in leaf elongation rate, plant stress, crowding and competition, among others (Krizek et al., 1985; Macklon et al., 1994).
-
van Noordwijk & Willigen,
1987Agricultural concepts of roots: from morphogenetic to functional equilibrium between root and shoot growthNetherlands Journal of Agricultural Science, 1987
-
Krizek et al., 1985Comparative effects of soil moisture stress and restricted root zone volume on morphogenetic and physiological responses of soybean [Glycine max (L.) Merr.]Journal of Experimental Botany, 1985
-
Macklon et al., 1994Soil P resources, plant growth and rooting characteristics in nutrient poor upland grasslandsPlant and Soil, 1994
Biomass by component
The results for dry biomass by component indicate that the trees in the initial sapling group have 72.6 % of aboveground biomass and 27.4 % of root biomass (Figure 3), while in the developed sapling group, 75.8 % corresponds to aboveground biomass and 24.2 % to root biomass (Figure 4). It is convenient to consider this rate of change to estimate the amount of Carbon in regeneration areas, especially when the space has a greater abundance of young plants. Previous studies have shown that there is a significant increase of up to five percentage points in root production in restrictive soils (Guerra et al., 2005), which could modify the results obtained if they were compared under conditions with lower soil nutritional quality.
-
Guerra et al., 2005Análisis de la biomasa de raíces en diferentes tipos de bosques. Avances en la evaluación de Pinus radiata en ChileBosque, 2005
Thumbnail
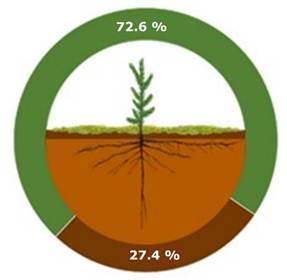
Figure 3
Aboveground and root biomass in initial sapling trees (6 to 65 cm tall).
Aboveground and root biomass in initial sapling trees (6 to 65 cm tall).
Thumbnail

Figure 4
Aboveground and root biomass in trees with developed saplings (68 to 150 cm tall).
Aboveground and root biomass in trees with developed saplings (68 to 150 cm tall).
Several authors estimate that 20 to 40 % of forest biomass is composed of roots (Brunner & Godbold, 2007; Finér et al., 2011; Sochacki et al., 2017); however, this biomass changes according to the species, the climate and the characteristics of the ecosystem in which they grow.
-
Brunner & Godbold, 2007Tree roots in a changing worldJournal of Forest Research, 2007
-
Finér et al., 2011Factors causing variation in fine root biomass in forest ecosystemsForest Ecology and Management, 2011
-
Sochacki et al., 2017Accuracy of tree root biomass sampling methodologies for carbon mitigation projectsEcological Engineering, 2017
Accumulated biomass potential
An average regeneration density of 3 925 trees ha-1 at the sapling stage was estimated, but this result could be overestimated since the sampling sites were located in clear spaces in the canopy where environmental conditions allowed the establishment of A. religiosa regeneration; however, under the canopy of adult trees the density was low due to the low solar radiation that reaches the forest floor and the plants do not regularly establish themselves (Lara-González et al., 2009). Contrasting data by Hernández et al. (2022) with the percentage of dead seedlings under a partially closed canopy was 79 % (n=803), while under light gaps it was 70.1 % (n=384). And regarding the percentage of live seedlings under a partially closed canopy it was 17.9 % (n=182), while in light gaps it was 28.1 % (n=154).
-
Lara-González et al., 2009Regeneration of Abies religiosa in canopy gaps versus understory, Cofre de Perote National Park, MéxicoAgrociencia, 2009
-
Hernández et al.
(2022)Nicho de regeneración de Abies religiosa (Kunth) Schltdl. & Cham. en el Monte Tláloc, Parque Nacional Iztaccíhuatl-Popocatépetl, MéxicoBotanical Sciences, 2022
On the other hand, the potential of total biomass (aboveground and root) stored in A. religiosa at the sapling stage was 103.6 kg ha-1 (Figure 5). It is worth noting that the aboveground part-root ratio calculated was 3:1, that is, 3 parts correspond to aboveground biomass and one part to root biomass. This contributes to maintaining organic Carbon (OC) under the soil by inducing its stability through the mineralization process that occurs over time. Xia et al. (2022) mention that the lignin contained in the wood of the roots is a recalcitrant component that stabilizes the OC in the soil for decades, and fir forests have a high lignin content as do pine forests (Avendaño et al., 2009; Leifeld & Kögel-Knabner, 2005).
-
Xia et al. (2022)Characterizing natural variability of lignin abundance and composition in fine roots across temperate trees: a comparison of analytical methodsNew Phytologist, 2022
-
Avendaño
et al., 2009Estimación de biomasa y Carbono en un bosque de Abies religiosaRevista Fitotecnia Mexicana, 2009
-
Leifeld &
Kögel-Knabner, 2005Soil organic matter fractions as early indicators for carbon stock changes under different land-use?Geoderma, 2005
Thumbnail
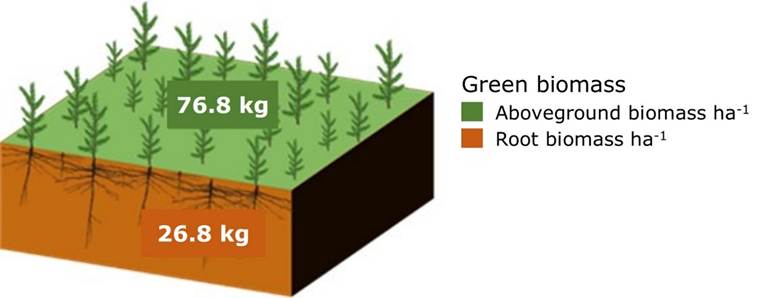
Figure 5
Potential storage of aboveground and root biomass per hectare of fir trees at the sapling stage in El Chico National Park, Hidalgo state, Mexico.
Potential storage of aboveground and root biomass per hectare of fir trees at the sapling stage in El Chico National Park, Hidalgo state, Mexico.
Root competition is greater when they grow with other root structures, so this is a factor that influences their conformation, extension and distribution in the soil and sometimes tends to reduce their depth, design and density (Bolte & Villanueva, 2006; Curt & Prévosto, 2003; Rewald & Leuschner, 2009). In this way, the percentage of root biomass during the growth of Abies religiosa in El Chico National Park could be reduced.
-
Bolte & Villanueva,
2006Interspecific competition impacts on the morphology and distribution of fine roots in European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.)European Journal of Forest Research, 2006
-
Curt & Prévosto,
2003Root biomass and rooting profile of naturally regenerated beech in mid-elevation Scots pine woodlandsPlant Ecology, 2003
-
Rewald & Leuschner,
2009Belowground competition in a broad-leaved temperate mixed forest: pattern analysis and experiments in a four-species standEuropean Journal of Forest Research, 2009
Conclusions
Abies religiosa forests at the sapling stage have the potential to accumulate aboveground-root biomass in a 3:1 ratio in canopy gaps where conditions allow the establishment of natural regeneration. Small plants aged 1 to 8 years on average store more root biomass in percentage than larger plants aged 8 to 12 years at the same sapling stage. The ratio of root biomass to aboveground biomass at the sapling stage changes with age in Abies religiosa trees.
Acknowledgements
The authors wish to express their gratitude to the National Council of Humanities, Sciences and Technologies (Conahcyt), for the financial scholarship granted to the first author, which allowed this research to be carried out.
Referencias
- Adame, M. F., Cherian, S., Reef, R. & Stewart-Koster, B. (2017). Mangrove root biomass and the uncertainty of belowground carbon estimations. Forest Ecology and Management, 403, 52-60. https://doi.org/10.1016/j.foreco.2017.08.016 Links
- Addo-Danso, S. D., Prescott, C. E. & Smith, A. R. (2016). Methods for estimating root biomass and production in forest and woodland ecosystem carbon studies: A review. Forest Ecology and Management, 359, 332-351. https://doi.org/10.1016/j.foreco.2015.08.015 Links
- Aguilar L., J. M. E. (2018). Estructura y diversidad de la vegetación arbórea de un bosque de galería en el estado de Puebla. Revista Mexicana de Ciencias Forestales, 9(47), 230-253. https://doi.org/10.29298/rmcf.v9i47.154 Links
- Albers, A., Collet, P., Benoist, A. & Hélias, A. (2019). Data and non-linear models for the estimation of biomass growth and carbon fixation in managed forests. Data in Brief, 23, Article 103841. https://doi.org/10.1016/j.dib.2019.103841 Links
- Avendaño H., D. M., Acosta M., M., Carrillo A., F. y Etchevers B., J. D. (2009). Estimación de biomasa y Carbono en un bosque de Abies religiosa. Revista Fitotecnia Mexicana, 32(3), 233-238. https://www.scielo.org.mx/pdf/rfm/v32n3/v32n3a11.pdf Links
- Bar-On, Y. M., Phillips, R. & Milo, R. (2018). The biomass distribution on Earth. Proceedings of the National Academy of Sciences, 115(25), 1-6. https://www.researchgate.net/publication/325276009_The_biomass_distribution_on_Earth Links
- Bolte, A. & Villanueva, I. (2006). Interspecific competition impacts on the morphology and distribution of fine roots in European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.). European Journal of Forest Research, 125, 15-26. https://doi.org/10.1007/s10342-005-0075-5 Links
- Brunner, I. & Godbold, D. L. (2007). Tree roots in a changing world. Journal of Forest Research, 12, 78-82. https://doi.org/10.1007/s10310-006-0261-4 Links
- Casiano-Domínguez, M., Paz-Pellat, F., Rojo-Martínez, M., Covaleda-Ocon, S. y Aryal, D. R. (2018). El Carbono de la biomasa aérea medido en cronosecuencias: primera estimación en México. Madera y Bosques, 24, Artículo e2401894. https://doi.org/10.21829/myb.2018.2401894 Links
- Comisión Nacional de Áreas Naturales Protegidas. (2005). Programa de conservación y manejo Parque Nacional El Chico. Comisión Nacional de Áreas Naturales Protegidas. http://centro.paot.org.mx/documentos/ine/chico.pdf Links
- Cortés-Blobaum, H. J., Rodríguez-Laguna, R., Otazo-Sánchez, E. M., Prieto-García, F., Fragoso-López, P. I. y Razo-Zárate, R. (2019). Patrones culturales de uso de leña en la primera área protegida de Latinoamérica, El Chico, México. Revista Iberoamericana de Ciencias, 6(2), 15-26. http://reibci.org/publicados/2019/abr/3400108.pdf Links
- Curt, T. & Prévosto, B. (2003). Root biomass and rooting profile of naturally regenerated beech in mid-elevation Scots pine woodlands. Plant Ecology, 167, 269-282. https://doi.org/10.1023/A:1023904912712 Links
- Djomo, A. N., Knohl, A. & Gravenhorst, G. (2011). Estimations of total ecosystem carbon pools distribution and carbon biomass current annual increment of a moist tropical forest. Forest Ecology and Management, 261(8), 1448-1459. https://doi.org/10.1016/j.foreco.2011.01.031 Links
- Finér, L., Ohashi, M., Noguchi, K. & Hirano, Y. (2011). Factors causing variation in fine root biomass in forest ecosystems. Forest Ecology and Management, 261(2), 265-277. https://doi.org/10.1016/j.foreco.2010.10.016 Links
- Fonseca G., W., Alice G., F. y Rey B., J. M. (2009). Modelos para estimar la biomasa de especies nativas en plantaciones y bosques secundarios en la zona Caribe de Costa Rica. Bosque, 30(1), 36-47. https://www.scielo.cl/pdf/bosque/v30n1/art06.pdf Links
- Fragoso-López, P. I., Rodríguez-Laguna, R., Otazo-Sánchez, E. M., González-Ramírez, C. A., Valdéz-Lazalde, J. R., Cortés-Blobaum, H. J. & Razo-Zárate, R. (2017). Carbon sequestration in protected areas: A case study of an Abies religiosa (H. B. K.) Schlecht. et Cham forest. Forests, 8(11), 429. https://doi.org/10.3390/f8110429 Links
- Fu, L., Zeng, W. & Tang, S. (2017). Individual tree biomass models to estimate forest biomass for large spatial regions developed using four pine species in China. Forest Science, 63(3), 241-249. https://doi.org/10.5849/FS-2016-055 Links
- González O., H. A., Cortés-Calva, P., Íñiguez D., L. I. y Ortega-Rubio, O. (2014). Las áreas naturales protegidas de México. Investigación y Ciencia de la Universidad Autónoma de Aguascalientes, (60), 7-15. https://www.redalyc.org/pdf/674/67431160002.pdf Links
- Guerra C., J., Gayoso A., J., Schlatter V., J. y Nespolo R., R. (2005). Análisis de la biomasa de raíces en diferentes tipos de bosques. Avances en la evaluación de Pinus radiata en Chile. Bosque, 26(1), 5-21. http://dx.doi.org/10.4067/S0717-92002005000100002 Links
- Hammer, Ø., Harper, D. A. T. & Ryan, P. D. (2001). PAST: Paleontological statistics software package for education and data analysis. Palaeontologia electrónica, 4(1), 1-9. https://palaeo-electronica.org/2001_1/past/past.pdf Links
- Hernández R., V., López-Mata, L., Cruz-Rodríguez, J. A. y Luna C., M. (2022). Nicho de regeneración de Abies religiosa (Kunth) Schltdl. & Cham. en el Monte Tláloc, Parque Nacional Iztaccíhuatl-Popocatépetl, México. Botanical Sciences, 100(2), 331-344. https://doi.org/10.17129/botsci.2912 Links
- Hutchinson, I. D. (1993). Puntos de partida y muestreo diagnóstico para la silvicultura de bosques naturales del trópico húmedo. Centro Agronómico Tropical de Investigación y Enseñanza, CATIE. https://www.google.com.mx/books/edition/Puntos_de_partida_y_muestreo_diagn%C3%B3stic/qRAIjOzSDAwC?hl=es&gbpv=1 Links
- Íñiguez D., L. I., Jiménez S., C. L., Sosa R., J. y Ortega-Rubio, A. (2014). Categorías de las áreas naturales protegidas en México y una propuesta para la evaluación de su efectividad. Investigación y Ciencia de la Universidad Autónoma de Aguascalientes, (60), 65-70. https://www.redalyc.org/pdf/674/67431160008.pdf Links
- Krizek, D. T., Carmi, A., Mirecki, R. M., Snyder, F. W. & Bunce, J. A. (1985). Comparative effects of soil moisture stress and restricted root zone volume on morphogenetic and physiological responses of soybean [Glycine max (L.) Merr.]. Journal of Experimental Botany, 36(1), 25-38. https://doi.org/10.1093/jxb/36.1.25 Links
- Lara-González, R., Sánchez-Velázquez, L. R. & Corral-Aguirre, J. (2009). Regeneration of Abies religiosa in canopy gaps versus understory, Cofre de Perote National Park, México. Agrociencia, 43(7), 739-747. https://www.scielo.org.mx/pdf/agro/v43n7/v43n7a8.pdf Links
- Leifeld, J. & Kögel-Knabner, I. (2005). Soil organic matter fractions as early indicators for carbon stock changes under different land-use? Geoderma, 124(1-2), 143-155. https://doi.org/10.1016/j.geoderma.2004.04.009 Links
- Litton, C. M., Raich, J. W. & Ryan, M. G. (2007). Carbon allocation in forest ecosystems. Global Change Biology, 13(10), 2089-2109. https://doi.org/10.1111/j.1365-2486.2007.01420.x Links
- López-Reyes, L. Y., Domínguez-Domínguez, M., Martínez-Zurimendi, P., Zavala-Cruz, J., Gómez-Guerrero, A. y Posada-Cruz, S. (2016). Carbono almacenado en la biomasa aérea de plantaciones de hule (Hevea brasiliensis Müell. Arg.) de diferentes edades. Madera y Bosques, 22(3), 49-60. https://doi.org/10.21829/myb.2016.2231456 Links
- Macklon, A. E. S., Mackie-Dawson, L. A., Sim, A., Shand, C. A. & Lilly, A. (1994). Soil P resources, plant growth and rooting characteristics in nutrient poor upland grasslands. Plant and Soil, 163, 257-266. https://doi.org/10.1007/BF00007975 Links
- Oliveira, N., Rodríguez-Soalleiro, R., Pérez-Cruzado, C., Cañellas, I., Sixto, H. & Ceulemans, R. (2018). Above-and below-ground carbon accumulation and biomass allocation in poplar short rotation plantations under Mediterranean conditions. Forest Ecology and Management, 428, 57-65. https://doi.org/10.1016/j.foreco.2018.06.031 Links
- Peichl, M., Leava, N. A. & Kiely, G. (2012). Above-and belowground ecosystem biomass, carbon and nitrogen allocation in recently afforested grassland and adjacent intensively managed grassland. Plant and Soil, 350(1), 281-296. https://www.researchgate.net/publication/225448541_Above-_and_belowground_ecosystem_biomass_carbon_and_nitrogen_allocation_in_recently_afforested_grassland_and_adjacent_intensively_managed_grassland Links
- Pham, M. V., Pham, T. M., Du, Q. V. V., Bui, Q. T., Tran, A. V., Pham H. M. & Nguyen, T. N. (2019). Integrating Sentinel-1A SAR data and GIS to estimate aboveground biomass and carbon accumulation for tropical forest types in Thuan Chau district, Vietnam. Remote Sensing Applications: Society and Environment, 14, 148-157. https://doi.org/10.1016/j.rsase.2019.03.003 Links
- Razo Z., R., Gordillo M., A. J., Rodríguez L., R., Maycotte M., C. C. y Acevedo S., O. A. (2015). Coeficientes de Carbono para arbustos y herbáceas del bosque de oyamel del Parque Nacional El Chico. Revista Mexicana de Ciencias Forestales , 6(31), 58-67. https://doi.org/10.29298/rmcf.v6i31.195 Links
- Rewald, B. & Leuschner, C. (2009). Belowground competition in a broad-leaved temperate mixed forest: pattern analysis and experiments in a four-species stand. European Journal of Forest Research, 128, 387-398. https://doi.org/10.1007/s10342-009-0276-4 Links
- Rodríguez-Laguna, R., Razo-Zárate, R., Fonseca-González, J., Capulín-Grande, J. y Goche-Telles, R. (2015). Regeneración natural post-incendio de Abies religiosa (H. B. K.) Schl. et Cham, en el Parque Nacional “El Chico” Hidalgo. Revista Iberoamericana de Ciencias, 2(2), 11-22. http://www.reibci.org/publicados/2015/marzo/0900104.pdf Links
- Romahn-Hernández, L. F., Rodríguez-Trejo, D. A., Villanueva-Morales, A., Monterroso-Rivas, A. I. y Pérez-Hernández, M. de J. (2020). Rango altitudinal: factor de vigor forestal y determinante en la regeneración natural del oyamel. Revista Entreciencias: Diálogos en la Sociedad del Conocimiento, 8(22), Artículo 10. https://doi.org/10.22201/enesl.20078064e.2020.22.72751 Links
- Ronquillo-Gorgúa, N., Razo-Zárate, R., Rodríguez-Laguna, R., Acevedo-Sandoval, O. A., Hernández-Ortiz, J. & Manzur-Chávez, N. (2022). Carbon storage during the development stages of Pinus patula Schiede ex Schltdl. & Cham. in the Sierra Alta of Hidalgo. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 28(3), 483-497. https://doi.org/10.5154/r.rchscfa.2022.02.009 Links
- Schuler, J., Bragg, D. C. & McElligott, K. (2017). Biomass estimates of small diameter planted and natural-origin loblolly pines show major departures from the national biomass estimator equations. Forest Science, 63(3), 319-330. https://doi.org/10.5849/FS-2016-047 Links
- Sochacki, S. J., Ritson, P., Brand, B., Harper, R. J. and Dell, B. (2017). Accuracy of tree root biomass sampling methodologies for carbon mitigation projects. Ecological Engineering, 98, 264-274. https://doi.org/10.1016/j.ecoleng.2016.11.004 Links
- StatSoft Inc. (2011). STATISTICA (data analysis software system) . Versión 10. http://www.statsoft.com Links
- van Noordwijk, M. & de Willigen, P. (1987). Agricultural concepts of roots: from morphogenetic to functional equilibrium between root and shoot growth. Netherlands Journal of Agricultural Science, 35(4), 487-496. https://doi.org/10.18174/njas.v35i4.16707 Links
- Xia, M, Valverde-Barrantes, O. J., Suseela, V., Blackwood, C. B. & Tharayil, N. (2022). Characterizing natural variability of lignin abundance and composition in fine roots across temperate trees: a comparison of analytical methods. New Phytologist, 236(6), 2358-2373. https://doi.org/10.1111/nph.18515 Links
- Zar, J. H. (2010). Biostatistical analysis. Prentice Hall. https://books.google.com.mx/books/about/Biostatistical_Analysis.html?id=LCRFAQAAIAAJ&redir_esc=y Links