










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias pecuarias
versión On-line ISSN 2448-6698versión impresa ISSN 2007-1124
Rev. mex. de cienc. pecuarias vol.16 no.3 Mérida jul./sep. 2025 Epub 08-Dic-2025
https://doi.org/10.22319/rmcp.v16i3.6778
Articles
Frequency of genes encoding antibiotic resistance in strains of Staphylococcus aureus from udder skin of cattle from southwestern Mexico
a Universidad Autónoma de Guerrero. Facultad de Ciencias Químico Biológicas. Laboratorio de Investigación en Patometabolismo Microbiano, Guerrero, México.
b Universidad Autónoma de Guerrero. Facultad de Ciencias Químico Biológicas. Laboratorio de Investigación en Microbiología, Guerrero, México.
c Universidad Autónoma de Guerrero. Facultad de Ciencias Químico Biológicas, SECIHTI-UAGro, Guerrero, México.
d Universidad Autónoma de Guerrero, Facultad de Ciencias Químico Biológicas, Laboratorio de Investigación en Microbiología Molecular y Biotecnología Ambiental, Guerrero, México.
e Secretaría de Salud Guerrero. Jurisdicción Sanitaria 02, Guerrero, México.
f Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán. Departamento de Infectología. Laboratorio de Microbiología. Ciudad de México, México.
The presence of Staphylococcus aureus in the skin of bovine udder has been proposed as a source of upward contamination to the mammary gland, which causes the development of mastitis and the presence of this bacterium in milk. The entry of S. aureus into the food chain becomes a public health problem due to the dissemination of antibiotic-resistant strains from animals to food and therefore to humans. Therefore, this study aimed to determine resistance to antibiotics and associated genes in strains of S. aureus isolated from the skin of bovine udder. Antibiotic resistance was determined by minimum inhibitory concentration and the presence of antibiotic resistance genes by endpoint PCR. The strains of S. aureus showed a high frequency of resistance to beta-lactams, macrolides, lincosamides, and tetracyclines. The strains were resistant to cefoxitin but the mecA gene was not found; of the ermA-C genes, ermB was the most frequently found gene (16.39 %); the msrA gene was found in most strains with constitutive resistance to macrolides and lincosamides (88.46 %); the tetM gene was found in 64.28 % of the tetracycline-resistant strains. In conclusion, the strains of S. aureus isolated from bovine udder skin presented different degrees of resistance to different groups of antibiotics, which can be explained by the presence of associated genes.
Keywords Staphylococcus aureus; Cow udder; Resistance genes; Antibiotics
La presencia de Staphylococcus aureus en la piel de ubre bovina se ha propuesto como una fuente de contaminación ascendente hacia la glándula mamaria, que ocasiona el desarrollo de mastitis y la presencia de esta bacteria en leche. La entrada de S. aureus a la cadena alimenticia se convierte en un problema de salud pública, debido a la diseminación de cepas resistentes a antibióticos desde los animales, a alimentos y por ende a humanos. Por lo anterior, el objetivo de este estudio fue determinar la resistencia a antibióticos y genes asociados en cepas de S. aureus aisladas de la piel de ubre bovina. La resistencia a antibióticos fue determinada por concentración mínima inhibitoria y la presencia de genes de resistencia a antibióticos por PCR en punto final. Las cepas de S. aureus presentaron elevada frecuencia de resistencia a betalactámicos, macrólidos, lincosamidas y tetraciclinas. Las cepas fueron resistentes a cefoxitina pero no se encontró el gen mecA; de los genes ermA-C, ermB fue el gen encontrado con mayor frecuencia (16.39 %); el gen msrA se encontró en la mayoría de las cepas con resistencia constitutiva a macrólidos y lincosamidas (88.46 %); el gen tetM se encontró en 64.28 % de las cepas resistentes a tetraciclina. En conclusión, las cepas de S. aureus aisladas de piel de ubre bovina presentaron diferentes grados de resistencia a diversos grupos de antibióticos, la cual puede ser explicada por la presencia de genes asociados.
Palabras clave Staphylococcus aureus; Ubre de vaca; Genes de resistencia; Antibióticos
Introduction
Bovine mastitis is the most prevalent infectious disease worldwide, which is associated with severe economic losses due to the reduction in milk production and quality, as well as the cost of treatments and animal health, increasing the risk of slaughter and replacement costs1-3. Staphylococcus aureus is recognized as the main pathogen worldwide that causes bovine mastitis, and this microorganism is considered contagious and difficult to treat due to its high rate of dissemination among infected animals and the ability to cause chronic or recurrent infections1,4,5.
Antibiotic therapy is an important measure to control bovine mastitis. Nevertheless, there has been an increasing number of studies reporting that S. aureus is resistant to multiple classes of antibiotics in response to selective pressure from their continued use, due to the overuse of these active substances in veterinary medicine and agriculture6,7.
In addition to the use of antibiotics for mastitis control, it is important to identify epidemiological patterns and identify sources of contamination. Several potential sources of contamination have been identified, from barn air, milking equipment to udder skin8. However, results about the role of bovine udder skin as a source of S. aureus in intramammary infections are not yet conclusive. On the one hand, it has been shown that strains of S. aureus from the skin and canal of the udder of the cow may be a potential source for the development of bovine mastitis because, genetically, they are the same as the strains obtained in milk and because these can promote the chronicity of the infection4,9. On the other hand, another study showed that most of the S. aureus associated with mastitis cases belong to strains that are highly adapted to the mammary gland but different from the strains that come from the skin8. Although the role of strains isolated from cow udder skin is unclear, these strains can easily be transferred to raw milk during milking. Raw milk contaminated with S. aureus can become a public health problem due to the risk of food poisoning by this microorganism, in addition to the dissemination of antibiotic-resistant strains4,10.
In a previous study, strains of S. aureus were isolated from the udder skin of cows from three dairy farms in southwestern Mexico, these strains presented the genes for enteroxins A, D and E. In addition, within these strains, clonal groups were identified that spread among the farms and remained in the rainy and dry seasons11. Due to the toxigenic potential of the strains, it is important to continue with the determination of other characteristics of epidemiological importance, such as antibiotic resistance, considering that their persistence in the farms may be related to high antibiotic resistance. Therefore, this study aimed to determine resistance to antibiotics and associated genes in strains of S. aureus isolated from the udder skin of cattle on farms in southwestern Mexico.
Material and methods
Bacterial strains
The strains of S. aureus from the teat of the cow udder were isolated in 2019 in two seasons of the year (rainy and dry) in three dairy farms located in three areas with different altitudes11. The strains are preserved in cryovials of glycerol at 15 % (v/v) at -20 °C in the strain collection of the Microbial Pathometabolism Research Laboratory (LIPM, for its initials in Spanish) of the Faculty of Chemical-Biological Sciences of the Universidad Autonoma de Guerrero. These strains were characterized under the following phenotypic profile: Gram-positive coccus, mannitol, catalase and coagulase positive and molecularly identified by the fem gene. The reactivation of the strains was carried out by resuspending 20 μl of each cryovial in 1 ml of Mueller Hinton (MH) broth, which was incubated at 37 °C for 24 h. Once the growth was obtained, they were inoculated in BHI agar plates for conservation as working strains.
Minimum inhibitory concentration
Antimicrobial resistance was assessed using the Minimum Inhibitory Concentration (MIC) assay; bacterial inocula were prepared from 24-h cultures of each of the strains in MH broth and adjusted to 0.5 McFarland. In a 96-well microplate, each antibiotic was placed in different ranges of concentrations; penicillin (PEN 0.25-2 μg/ml), ceftriaxone (CRO 0.5-32 μg/ml), ciprofloxacin (CIP 4-32 μg/ml), clindamycin (DA 4-256 μg/ml), erythromycin (E 8-256 μg/ml), kanamycin (KAN 16-256 μg/ml), gentamicin (GEN 16-256 μg/ml), tetracycline (TET 16-256 μg/ml), vancomycin (NPV 16-256 μg/ml), chloramphenicol (CLF 16-256 μg/ml), and trimethoprim (TMP 16-256 μg/ml), indicated by the CLSI (Clinical and Laboratory Standards Institute)12. The bacterial inocula were then added and incubated at 37°C for 24 ± 1 h. As a positive control, the same strains were inoculated without antibiotics and as a negative control, a well with Mueller Hinton broth and antibiotics was used. Subsequently, the growth was read by absorbance at a wavelength of 492/630 nm in a spectrophotometer (Thermo Scientific®, GENESYS 200, USA). The MIC50 was then calculated and the strains were catalogued parametrically and according to the MIC values reported by the CLSI manual as Sensitive (S) or Resistant (R).
Resistance to beta-lactams
Each strain was incubated for 18 h in MH broth at 37 °C under static conditions; the culture was adjusted to 0.5 McFarland and inoculated in MH agar using the disc diffusion method with cefoxitin (FOX 30 μg) and penicillin (PEN 10 μg) discs and incubated at 35 ± 2 °C for 24 h12; after that time had elapsed, inhibition halos were measured and categorized as sensitive (S) (PEN ≥ 29 mm or FOX ≥25 mm) or resistant (R) (PEN ≤28 mm or FOX ≤24 mm)12. Then, 10 μl of nitrocefin (Oxoid Thermo Scientific™) was placed to determine the production of beta-lactamases by observing the change in the coloration of the halo: a reddish halo allowed them to be defined as producing and when no change in the coloration of the halo was observed as non-producing.
Determination of MLSB resistance phenotypes
The phenotypes of resistance to MLSB (Macrolides-Lincosamides-Streptogramin B) were determined by the results obtained from resistance to clindamycin (C-R) or erythromycin (E-R), classifying the resistances obtained as constitutive (C-R, E-R or both) or inducible (E-R, but sensitive to clindamycin)12.
Antibiotic resistance genes
DNA extraction
The genomic DNA of all S. aureus strains was obtained from 1 ml of an 18-h static culture in brain heart infusion (BHI) incubated at 37 °C. The cells were centrifuged at 10,000 rpm for 10 min, resuspending the pellets in 300 μl of lysis buffer (10 mM Tris HCl, 1 mM EDTA pH 8.0, lysozyme 0.02 mg/ml) and incubating at 37 °C for 10 min. Subsequently, 200 μl of phenol/chloroform/isoamyl alcohol solution (25:24:1 v/v/v) was added, inverted 10 to 15 times, centrifuged again at 10,000 rpm for 10 min, 200 μl of the aqueous phase was recovered and transferred to 1 ml of cold ethanol (96 %). It was incubated at -20 °C for 24 h, then centrifuged at 10,000 rpm for 10 min, the supernatant was discarded, the cell pellets were left to dry at room temperature and resuspended in 20 μl of sterile water13.
Genes that confer antibiotic resistance
The identification of genes conferring antibiotic resistance was determined by endpoint polymerase chain reaction (PCR). For the mecA, blaZ, ermA-C, msrA, and tetM genes, the following reaction mixture was used: 1X Taq 2x Master Mix RED, 1.5 MgCl2 (Ampliqon, Denmark), 40 nM of each oligonucleotide, and 100 ng of DNA. In the case of ermB, the following reaction mixture was used: 1X Taq 2x Master Mix RED, 1.5 MgCl2 (Ampliqon, Denmark), 160 nM of each oligonucleotide, and 100 ng of DNA. The ATCC 43300 strain of S. aureus subsp. aureus Rosenbach for ermA, mecA, blaZ; for the ermB gene, a clinical isolate of S. aureus characterized in the LIPM was used; the ATCC 25923 strain of S. aureus subsp. aureus Rosenbach and the strain 1084 of S. hominis characterized by the Microbiology Research Laboratory of the Faculty of Chemical-Biological Sciences of the Autonomous University of Guerrero were used as controls for the tetM and msrA genes, respectively. Oligonucleotides and conditions are described in Table 1.
Table 1 Oligonucleotides used in this study
| Gene | Sequence | Cycling | Expected size (bp) | Reference |
|---|---|---|---|---|
| blaZ | F-ATTTTGAAAAAGTTAATATTTTTAATTG R-CATTACACTCTTGGCGGTTTC |
1 cycle 95°C for 5 min; 35 cycles at 95°C for 30 s, 52°C for 50 s, and 72°C for 50 s; and 1 cycle at 72°C for 10 min. |
833 | 14 |
| mecA | F- AAAATCGATGGTAAAGGTTGGC R- AGTTCTGCAGTACCGGATTTGC |
1 cycle 94°C for 5 min; 32 cycles 94°C for 30 s, 57°C for 45 s, and 72°C for 1 min; and 1 cycle 72°C for 5 min. |
583 | 15 |
| ermA | F-GTTCAAGAACAATCAATACAGAG R-GGATCAGGAAAAGGACATTTTAC |
1 cycle 94°C for 2 min; 32 cycles 94°C for 30 s, 52°C for 1 min, and 72°C for 30 s; and 1 cycle 72°C for 10 min. |
421 | 16 |
| ermB | F-CCGTTTACGAAATTGGAACAGGTAAAGGGC R- GAATCGAGACTTGAGTGTGC |
1 cycle 95°C for 5 min; 32 cycles 95°C for 20 s, 52°C for 45 s, and 72°C for 40 s; and 1 cycle 72°C for 7 min. |
359 | |
| ermC | F- RGCTAATATTGTTTAAATCGTCAATTCC R-GGATCAGGAAAAGGACATTTTAC |
1 cycle 94°C for 2 min; 32 cycles 94°C for 30 s, 52°C for 1 min, and 72°C for 30s; and 1 cycle 72°C for 10 min. |
572 | |
| msrA | F-GGCACAATAAGAGTGTTTAAAGG R-AAGTTATATCATGAATAGATTGTCCTGTT |
1 cycle 95°C for 15 min; 32 cycles 95°C for 30 s, 52°C for 45 s, and 72°C for 1 min; and 1 cycle 72°C for 7 min. |
940 | 17 |
| tetM | F- GTGGACAAAGGTACAACGAG R-CGGTAAAGTTCGTCACACAC |
1 cycle 95°C for 5 min; 32 cycles 95°C for 20 s, 52°C for 45 s, and 72°C for 40 s; and 1 cycle 72°C for 7 min |
406 | 16 |
Results
Antibiotic resistance
It was determined that 95.1 % (58/61) of the strains are resistant to penicillin and 100 % (61/61) are resistant to ceftriaxone. It was found that only 1.6 % of the strains (1/61) were resistant to ciprofloxacin, gentamicin, and chloramphenicol, 98.4 % (60/61) to clindamycin, 42.6 % (26/61) to erythromycin, 62.3 % (38/61) to kanamycin, 22.9 % (14/61) to tetracycline, 13.1 % (8/61) to trimethoprim. Finally, no vancomycin-resistant strains were found.
Regarding the results of the antibiotic concentrations to which each of the strains analyzed in this work are inhibited. The 58 strains with penicillin resistance were distributed in MICs from 0.25 to 2.0 μg/ml; in the case of ceftriaxone, in the 61 resistant strains, MICs from 0.25 μg/ml to greater than 32 μg/ml were determined, of which 40 strains had the highest MIC (32 μg/ml).
In the MLSB antibiotic group, clindamycin-resistant strains (60/61) had MICs from 8 to 256 μg/ml, with 44 strains having a MIC of 256 μg/ml; for erythromycin, the resistant strains had MICs from 16 to 256 μg/ml, of which four strains had a MIC of 256 μg/ml; of the fluoroquinolone group, only one strain with a MIC of 4 μg/ml of ciprofloxacin; in aminoglycosides, strains were determined with MICs from 32 to 256 μg/ml concentration of kanamycin and a strain with a MIC of 64 μg/ml of gentamicin.
Regarding tetracycline, there were strains with MICs from 32 to 256 μg/ml, of which 11 strains had a MIC of 256 μg/ml; in amphenicols, there was only one strain with a MIC of 128 μg/ml of chloramphenicol; finally, in the diaminopyrimidine group, values of 32 to 256 μg/ml were determined in strains with resistance to trimethoprim, observing two strains with a MIC of 256 μg/ml (Table 2).
Table 2 Minimum inhibitory concentration of S. aureus strains resistant to the different groups of antibiotics
| Antibiotics | Concentrations used (µg/ml) | Resistant strains % (n), N= 61 |
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | 256 | ||
| Beta-lactams | ||||||||||||
| Penicillin | 9 | 5 | 17 | 27 | 95.1 (58) | |||||||
| Cephalosporins | ||||||||||||
| Ceftriaxone | 1 | 5 | 1 | 10 | 4 | 0 | 40 | 100 (61) | ||||
| Fluoroquinolones | ||||||||||||
| Ciprofloxacin | 1 | 1.6 (1) | ||||||||||
| Lincosamides | ||||||||||||
| Clindamycin Macrolides | 10 | 0 | 0 | 4 | 2 | 44 | 98.4 (60) | |||||
| Erythromycin | 11 | 9 | 0 | 2 | 4 | 42.6 (26) | ||||||
| Aminoglycosides | ||||||||||||
| Kanamycin | 10 | 9 | 7 | 12 | 62.3 (38) | |||||||
| Gentamicin | 1 | 1.6 (1) | ||||||||||
| Tetracyclines | ||||||||||||
| Tetracycline | 1 | 0 | 2 | 11 | 22.9 (14) | |||||||
| Phenicols | ||||||||||||
| Chloramphenicol | 1 | 1.6 (1) | ||||||||||
| Diaminopyrimidines | ||||||||||||
| Trimethoprim | 4 | 0 | 2 | 2 | 13.1 (8) | |||||||
The parameters of the MIC (μg/ml) were determined according to the values established in the 31ed M100 Performance Standards for Antimicrobial Susceptibility Testing12.
Antimicrobial resistance phenotypes
One hundred percent of the strains were resistant to cefoxitin and only one was beta-lactamase-producing (1.6 %). For erythromycin and clindamycin, resistance frequencies of 42.6 and 98.7 % were obtained, respectively, which were categorized within the group of phenotypes of constitutive resistance to MLSB, highlighting that the same 42.6 % of strains were resistant to both antibiotics (erythromycin and clindamycin). No strains with resistance to erythromycin but sensitivity to clindamycin were determined, so there is no inducible resistance of an antibiotic; on the other hand, an atypical result of 55.8 % of strains with resistance to clindamycin (C-R), without being resistant to erythromycin (S), was found. Eighty point two percent of the strains are resistant to at least three groups of antibiotics (Table 3)
Table 3 Antibiotic resistance phenotypes in S. aureus strains
| Beta-lactam resistance phenotype | Number of strains (%) N=61 |
|---|---|
| MRSA | 61 (100.0) |
| MSSA | 0 (0.0) |
| Production of beta-lactamases | 1 (1.6) |
| Phenotypes MLSB | (%) |
| cMLSB | |
| E-R | 26 (42.6) |
| C-R | 60 (98.7) |
| E-R, C-R | 26 (42.6) |
| iMLSB | |
| Antibiotic resistance phenotypes | (%) |
| PEN, CRO | 1 (1.6) |
| PEN, CRO, DAN | 11 (18.2) |
| PEN, CRO, DAN, E | 3 (4.9) |
| PEN, CRO, DAN, E, KAN | 13 (21.3) |
| PEN, CRO, DAN, E, KAN, TET | 4 (6.7) |
| PEN, CRO, DAN, E, KAN, CIP | 1 (1.6) |
| PEN, CRO, DAN, E, KAN, TMP | 1 (1.6) |
| PEN, CRO, DAN, E, TET | 1 (1.6) |
| PEN, CRO, DAN, E, TMP | 1 (1.6) |
| PEN, CRO, DAN, E, TMP, CLF, TET | 1 (1.6) |
| PEN, CRO, DAN, KAN | 14 (23.1) |
| PEN, CRO, DAN, KAN, GEN, TET | 1 (1.6) |
| PEN, CRO, DAN, KAN, TET | 3 (4.9) |
| PEN, CRO, DAN, TMP | 3 (4.9) |
| PEN, CRO, DAN, TMP, TET | 1 (1.6) |
| CRO, DAN, E | 1 (1.6) |
| CRO, DAN, E, KAN, TET | 1 (1.6) |
PEN (penicillin), CRO (ceftriaxone), DA (clindamycin), E (erythromycin), KAN (kanamycin), GEN (gentamicin), TMP (trimethoprim), TET (tetracycline), CLF (chloramphenicol), CIP (ciprofloxacin), MRSA (methicillin-resistant Staphylococcus aureus), MSSA (methicillin-sensitive Staphylococcus aureus), MLSB (Macrolides- Lincosamides- Streptogramin B), cMLSB (constitutive resistance to Macrolides- Lincosamides- Streptogramin B), E-R (resistance to erythromycin), C-R (resistance to clindamycin), iMLSB (inducible resistance to Macrolides-Lincosamides-Streptogramin B).
Antibiotic resistance genes
Absolute frequencies of the genes that confer resistance to each of the groups of antibiotics were determined. Of the beta-lactam-resistant strains, only three strains were found positive for the presence of the blaZ gene (4.91 %), one strain was positive for the nitrocefin test, highlighting that it was not positive for the presence of the blaZ gene. For mecA, no positive strains were found for the presence of this gene (Table 4) (Figure 1).
Table 4 Distribution of antibiotic resistance genes in S. aureus strains
| Resistant strains | Beta-lactams | MLS | TETRA | |||||
|---|---|---|---|---|---|---|---|---|
| Group of antibiotics | Antibiotic | blaZ | mecA | ermA | ermB | ermC | msrA | tetM |
| Beta-lactams | ||||||||
| Penicillin (N=58) | 3 | |||||||
| Cephalosporins | ||||||||
| Ceftriaxone (N=61) | 3 | |||||||
| Cefoxitin (N=61) | 3 | |||||||
| ML | ||||||||
| Clindamycin/ Erythromycin (N=26) | 9 | 1 | 23 | |||||
| Clindamycin (N=60) | 1 | 1 | 29 | |||||
| TETRA | ||||||||
| Tetracycline (N=14) | 9 | |||||||
TETRA= tetracycline.
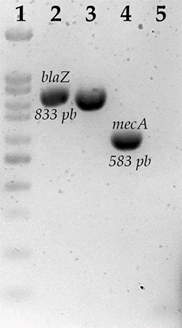
Agarose gel electrophoresis of blaZ and mecA amplification. 1) 100 bp molecular weight marker; 2) blaZ, S. aureus ATCC 43300; 3) blaZ, S. aureus S680; 4) mecA, S. aureus ATCC 43300; 5) mecA, S. aureus.
Figure 1 Beta-lactam resistance-associated genes in S. aureus strains isolated from bovine udder skin
From the MLSB antibiotic group: 52 strains of the total MLSB-resistant strains were determined with the presence of the msrA gene (86.6 %); of these, 29 strains belong to the C-R profile (55.7 %) and 23 corresponded to the ER-CR profile (44.23 %). In addition, there were 10 strains positive for the ermB gene (16.6 %), of which only nine correspond to the ER-CR phenotype (90 %) and the remaining strain is of the C-R phenotype (10 %). Likewise, two strains were determined with the presence of the ermC gene (3.3 %), of which one strain corresponds to the ER-CR phenotype and one strain to the C-R phenotype, and there were no ermA-positive strains (Figure 2).
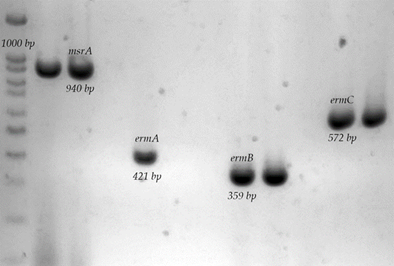
Agarose gel electrophoresis of the amplification of msrA, ermA-C genes. 1) 100 bp molecular weight marker; 2) S. haemolyticus 1035, msrA+; 3) S. aureus S697; 4) Negative control; 5) S. aureus ATCC 43300, ermA+; 6) S. aureus S661; 7) Negative control; 8) S. aureus S697, ermB+; 9) S. aureus S694, ermB+; 10) Negative control; 11) S. aureus S662, ermC+; 12) S. aureus S661, ermC+.
Figure 2 Macrolide resistance-associated genes in S. aureus strains isolated from bovine udder skin
In the determination of the tetM gene of 14 tetracycline-resistant strains, nine were determined with the presence of the tetM gene (64.28 %) (Figure 3); of these, six strains had a MIC of 256 μg/ml, one strain had a MIC of 128 μg/ml, and two strains had a MIC of 32 μg/ml.
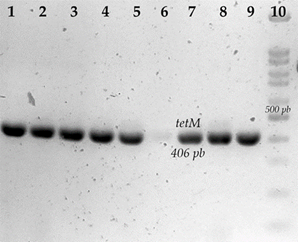
Agarose gel electrophoresis of tetM amplification. 1) S. aureus S68; 2) S. aureus S689; 3) S. aureus S692; 4) S. aureus S693; 5) S. aureus S696; 6) S. aureus S697; 7) S. aureus S654; 8) S. aureus S662; 9) S. aureus D1; 10) 100 bp molecular weight marker.
Figure 3 Tetracycline resistance-associated genes in S. aureus strains isolated from bovine udder skin
Discussion
S. aureus is the main pathogen worldwide that causes bovine mastitis, characterized by causing chronic infections due to the low response to antibiotics, facilitating spreading among cattle. The antibacterial resistance of this microorganism originates from the extensive use of antibiotics to control mastitis, becoming a public health problem due to the emergence and dissemination of antibiotic-resistant strains to humans from food18.
This study found a high resistance to beta-lactam antibiotics compared to other groups of antibiotics in S. aureus strains isolated from the skin of the bovine udder; resistance to these antibiotics is mediated by the synthesis of alternative penicillin-binding proteins (PBP2a, PBP2 ́), synthesis of beta-lactamases, and mutations in penicillin-binding protein (PBP) genes. Resistance to cefoxitin infers that the mechanism could be mediated by the synthesis of alternate proteins encoded in the SCCmec chromosomal cassette; nevertheless, in this study, the strains were negative for the amplification of the mecA gene. Strains negative for mecA but with a high oxacillin MIC have been reported as mecA-BORSA (borderline oxacillin-resistant S. aureus, mecA negative) in different foods, such as chicken, pork, milk, and even in the community19, so its presence in bovine udder skin is not surprising. A key characteristic of the BORSA strains is the overproduction of beta-lactamases20; in this sense, to evaluate the presence of genes related to beta-lactamases, the blaZ gene was amplified, obtaining a low frequency for this gene (3/61, 4.91 %), the low frequency of beta-lactamase-producing strains encoded by blaZ and negative for mecA could be related to the presence of new beta-lactamases not encoded by blaZ but by plasmids21. On the other hand, to evaluate the production of beta-lactamases, nitrocefin hydrolysis was assessed, obtaining a low frequency (1/61, 1.64 %), emphasizing that the nitrocefin-test positive strain is negative for the blaZ gene. Regarding this point, up to four different beta-lactamases have been described in S. aureus, called A to D; however, not all of them can be determined by the nitrocefin, cefazolin or cephaloridine suspension test22. Although the main mechanism of resistance in BORSA strains is the overproduction of beta-lactamases, this is not the only mechanism; the presence of new beta-lactamases, mutations in PBP genes or the presence of efflux pumps could explain the behavior of these strains and could be studied in the future20,21,23.
Macrolides, lincosamides, and streptogramins of group B, called MLSB, have become a strategy for the control of methicillin-resistant S. aureus, as well as in patients with allergy to the beta-lactam group. Of the MLSB group, clindamycin is important due to its application for the treatment of skin, soft tissue and bone infections due to its high permeability24-26. Regarding the above, the farmers who participated in this study report the use of this antibiotic as a preventive treatment for bovine udder infections, which is reflected in the high resistance to clindamycin (98.3 %, 60/61) in a constitutive phenotype with macrolides (cMLSB) in 42.6 % of the strains. MLSB resistance may be conferred by different mechanisms, such as target site modification, efflux pumps, and enzymatic inactivation of the antibiotic27. The modification of the target site is mediated by the presence of erythromycin resistance methylase (erm) genes. The main commonly found erm genes include ermA and ermC17. The erm genes encode methylases that cause conformational modifications of the 23S rRNA subunit, resulting in a decrease in the binding of MLSB antibiotics to their target site in the ribosomal 50S subunit24. One of the relevant data of this study is the high prevalence of the ermB gene in S. aureus strains isolated from bovine udder, which has also been found in coagulase-positive and negative Staphylococcus strains isolated from buffalo milk28; unlike strains of clinical origin where it is common to find the ermA and ermC genes29-31. As described, target site modification is not the only mechanism that explains constitutive resistance to MLSB; another mechanism is the presence of efflux pumps, such as MsrA, which explains the constitutive resistance to macrolides32. The presence of both mechanisms could explain to some extent the constitutive phenotype of MLSB (E-R, C-R) found in this study, even leaving the circulation of other erm genes into consideration. In those strains of S. aureus that are only resistant to clindamycin (55.8 %), although it is an uncommon result, this could be explained by the presence of linA and linB genes that inactivate only lincosamides33 in the circulating strains and that could be one of the future perspectives of this study.
Tetracycline is an antibiotic that has been used in livestock, humans, small animals, agriculture, and aquaculture for the last 40 yr34. At the same time, tetracycline is not only used for the treatment of infections in animals but also as a growth promoter or to improve the efficiency of fattening products, a practice that is still carried out today both in the United States of America and in other countries35. Therefore, it is not surprising that tetracycline-resistant strains of S. aureus from cattle36 circulate and even that livestock-associated methicillin-resistant S. aureus (LA-MRSA) of the CC9 clonal complex has tetracycline resistance as a distinctive characteristic37. In this sense, 14 strains with resistance to tetracycline were identified, noting that they share other characteristics of bacterial resistance, including clindamycin, erythromycin and cefoxitin, so it is not ruled out that these strains also belong to this clonal group. Because of this, complete characterization of these strains is necessary in the future. Tetracycline resistance is mediated by genes encoding efflux pumps and ribosomal protection proteins35. In the latter group, one of the main genes sought is tetM. In this study, the frequency of the tetM gene was 14.75 %, being found in 9 of the 14 resistant strains. This gene has commonly been reported in isolated strains of subclinical mastitis in Brazil, China, and Canada38-40, samples of cow’s milk in Brazil41,42, and buffalo milk samples in Egypt28. In strains that are resistant to tetracycline but negative for the tetM gene, the search could focus on more than 25 genes related to tetracycline resistance34,35.
The determination of resistance genes in S. aureus as well as in other microorganisms is important because many of these genes are located in mobile genetic elements (MGEs), which, by horizontal transfer mechanisms, can generate new antibiotic-resistant strains. These MGEs have even been sought from environmental samples due to their importance43,44.
Conclusions and implications
The circulation of strains of S. aureus from the skin of bovine udders with resistance to beta-lactams, macrolides, lincosamides, and tetracyclines with the presence of the genes blaZ, ermABC, mrsA and tetM genes was confirmed in the milk production units analyzed. Although the present results do not allow generalization, it is suggested that adequate monitoring of the development of mastitis cases, the implementation of good milking practices on ranches in the region, and the appropriate use of antimicrobials would help to prevent the circulation of S. aureus strains from the skin of the bovine udder to milk and eventually to humans by the consumption of milk and their derivatives, favoring animal health as well as human and environmental health.
Literatura citada
1 . Da Costa LB, Rajala-Schultz PJ, Hoet A, Seo KS, Fogt K, Moon BS. Genetic relatedness and virulence factors of bovine Staphylococcus aureus isolated from teat skin and milk. J Dairy Sci 2014;97(11):6907-6916. [ Links ]
2. Hogeveen H, Huijps K, Lam T. Economic aspects of mastitis: New developments. N Z Vet J 2011;59(1):16-23. [ Links ]
3. Rasool Z, Noreen H, Anjum A, Rizvi A, Rabaan AA, Halwani MA, et al. Genotypic and phenotypic characterization of erythromycin-resistant Staphylococcus aureus isolated from bovine mastitis and humans in close contact. Trop Med Infect Dis 2022;8(1):26. [ Links ]
4. Piccinini R, Cesaris L, Daprà V, Borromeo V, Picozzi C, Secchi C, et al. The role of teat skin contamination in the epidemiology of Staphylococcus aureus intramammary infections. J Dairy Res 2009;76(1):36-41. [ Links ]
5. Sakwinska O, Giddey M, Moreillon M, Morisset D, Waldvogel A, Moreillon P. Staphylococcus aureus host range and human-bovine host shift. Appl Environ Microbiol 2011;77(17):5908-5915. [ Links ]
6. Artursson K, Söderlund R, Liu L, Monecke S, Schelin J. Genotyping of Staphylococcus aureus in bovine mastitis and correlation to phenotypic characteristics. Vet Microbiol 2016;193:156-161. [ Links ]
7. Gomes F, Henriques M. Control of bovine mastitis: Old and recent therapeutic approaches. Curr Microbiol 2016;72(4):377-382. [ Links ]
8. Zadoks RN, Van Leeuwen WB, Kreft D, Fox LK, Barkema HW, Schukken YH, et al. Comparison of Staphylococcus aureus isolates from bovine and human skin, milking equipment, and bovine milk by phage typing, pulsed-field gel electrophoresis, and binary typing. J Clin Microbiol 2002;40(11):3894-3902. [ Links ]
9. Haveri M, Hovinen M, Roslöf A, Pyörälä S. Molecular types and genetic profiles of Staphylococcus aureus strains isolated from bovine intramammary infections and extramammary sites. J Clin Microbiol 2008;46(11):3728-3735. [ Links ]
10. Ren Q, Liao G, Wu Z, Lv J, Chen W. Prevalence and characterization of Staphylococcus aureus isolates from subclinical bovine mastitis in southern Xinjiang, China. J Dairy Sci 2020;103(4):3368-3380. [ Links ]
11. Adame-Gómez R, Toribio-Jimenez J, Castro-Alarcón N, Talavera-Alarcón K, Flores-Gavilan J, Pineda-Rodríguez SA, et al. Diversidad genética y factores de virulencia de cepas de Staphylococcus aureus aisladas de la piel de ubre bovina. Rev Mex Cienc Pecu 2021;12(3):665-680. [ Links ]
12. CLSI. Performance Standards for Antimicrobial Susceptibility Testing. 2021. [ Links ]
13. Adame-Gómez R, Muñoz-Barrios S, Castro-Alarcón N, Leyva-Vázquez MA, Toribio-Jiménez J, Ramírez-Peralta A. Prevalence of the strains of Bacillus cereus group in artisanal Mexican cheese. Foodborne Pathog Dis 2020;17(1):8-14. [ Links ]
14. El-Maghraby AS, Aziz S, Mwafy A. Efficacy of Staphylococcus aureus vaccine in chicken. J Worlds Poult Res 2020;10(2s):203-213. [ Links ]
15. Al-Amery K, Elhariri M, Elsayed A, El-Moghazy G, Elhelw R, El-Mahallawy H, et al. Vancomycin-resistant Staphylococcus aureus isolated from camel meat and slaughterhouse workers in Egypt. Antimicrob Resist Infect Control 2019;8(1):129. [ Links ]
16. Chai M, Sukiman MZ, Kamarun Baharin AH, Ramlan I, Lai LZ, Liew Y, et al. Methicillin-resistant Staphylococcus aureus from peninsular Malaysian animal handlers: molecular profile, antimicrobial resistance, immune evasion cluster and genotypic categorization. Antibiotics 2022;11(1):103. [ Links ]
17. Lina G, Quaglia A, Reverdy ME, Leclercq R, Vandenesch F, Etienne J. Distribution of genes encoding resistance to macrolides, lincosamides, and streptogramins among staphylococci. Antimicrob Agents Chemother 1999;43(5):1062-1066. [ Links ]
18. Molineri AI, Camussone C, Zbrun MV, Suárez Archilla G, Cristiani M, Neder V, et al. Antimicrobial resistance of Staphylococcus aureus isolated from bovine mastitis: Systematic review and meta-analysis. Prev Vet Med 2021;188:105261. [ Links ]
19. Zehra A, Gulzar M, Singh R, Kaur S, Gill JPS. Comparative analysis of methicillin-resistant Staphylococcus aureus (MRSA) and borderline oxacillin resistant Staphylococcus aureus (BORSA) in community and food of animal origin. FEMS Microbiol Lett 2020;367(23):fnaa201. [ Links ]
20. Hryniewicz MM, Garbacz K. Borderline oxacillin-resistant Staphylococcus aureus (BORSA) - a more common problem than expected? J Med Microbiol 2017;66(10):1367-1373. [ Links ]
21. Massidda O. Evidence for a methicillin-hydrolysing β-lactamase in Staphylococcus aureus strains with borderline susceptibility to this drug. FEMS Microbiol Lett 1992;92(3):223-227. [ Links ]
22. Zygmunt DJ, Stratton CW, Kernodle DS. Characterization of four beta-lactamases produced by Staphylococcus aureus. Antimicrob Agents Chemother 1992;36(2):440-445. [ Links ]
23. Jo A, Ahn J. Phenotypic and genotypic characterisation of multiple antibiotic-resistant Staphylococcus aureus exposed to subinhibitory levels of oxacillin and levofloxacin. BMC Microbiol 2016;16(1):170. [ Links ]
24. Cetin ES, Gunes H, Kaya S, Aridogan BC, Demirci M. Macrolide-lincosamide-streptogramin B resistance phenotypes in clinical staphylococcal isolates. Int J Antimicrob Agents 2008;31(4):364-368. [ Links ]
25. Daurel C, Huet C, Dhalluin A, Bes M, Etienne J, Leclercq R. Differences in potential for selection of clindamycin-resistant mutants between inducible erm (a) and erm (c) Staphylococcus aureus genes. J Clin Microbiol 2008;46(2):546-550. [ Links ]
26. Prabhu K, Rao S, Rao V. Inducible clindamycin resistance in Staphylococcus aureus isolated from clinical samples. J Lab Physicians 2011;3(01):025-027. [ Links ]
27. Leclercq R. Mechanisms of resistance to macrolides and lincosamides: Nature of the resistance elements and their clinical implications. Clin Infect Dis 2002;34(4):482-492. [ Links ]
28. Osman KM, Pires ÁDS, Franco OL, Orabi A, Hanafy MH, Marzouk E, et al. Enterotoxigenicity and antibiotic resistance of coagulase-negative staphylococci isolated from raw buffalo and cow milk. Microb Drug Resist 2020;26(5):520-530. [ Links ]
29. Aydeniz Ozansoy F, Cevahir N, Kaleli İ. Investigation of macrolide, lincosamide and streptogramin B resistance in Staphylococcus aureus strains isolated from clinical samples by phenotypical and genotypical methods. Mikrobiyol Bul 2015;49(1):1-14. [ Links ]
30. Malla Thakuri DR, Pokhrel A, Amatya R, Sharma Bashyal N, Neupane M, Kc S, et al. Distribution of meca and erm genes among methicillin-resistant Staphylococcus aureus with inducible resistance to clindamycin. J Nepal Health Res Counc 2023;21(1):29-33. [ Links ]
31. Nagarkoti D, Prajapati K, Sharma AN, Gyawali A, Manandhar S. Distribution of macrolide-lincosamide-streptogramin B antibiotics resistance genes in clinical isolates of staphylococci. J Nepal Health Res Counc 2021;18(4):734-740. [ Links ]
32. Schmitz FJ, Sadurski R, Kray A, Boos M, Geisel R, Kohrer K, et al. Prevalence of macrolide-resistance genes in Staphylococcus aureus and Enterococcus faecium isolates from 24 European university hospitals. J Antimicrob Chemother 2000;45(6):891-894. [ Links ]
33. Roberts MC, Sutcliffe J, Courvalin P, Jensen LB, Rood J, Seppala H. Nomenclature for macrolide and macrolide-lincosamide-streptogramin B resistance determinants. Antimicrob Agents Chemother 1999;43(12):2823-2830. [ Links ]
34. Roberts MC, Schwarz S. Tetracycline and phenicol resistance genes and mechanisms: importance for agriculture, the environment, and humans. J Environ Qual 2016;45(2):576-592. [ Links ]
35. Chopra I, Roberts M. Tetracycline antibiotics: mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol Mol Biol Rev MMBR 2001;65(2):232-260. [ Links ]
36. Nemati G, Romanó A, Wahl F, Berger T, Rojo LV, Graber HU. Bovine Staphylococcus aureus: a European study of contagiousness and antimicrobial resistance. Front Vet Sci 2023;10:1154550. [ Links ]
37. Ye X, Wang X, Fan Y, Peng Y, Li L, Li S, et al. Genotypic and phenotypic markers of livestock-associated methicillin-resistant Staphylococcus aureus CC9 in humans. Appl Environ Microbiol 2016;82(13):3892-3899. [ Links ]
38. De Oliveira RP, Da Silva JG, Aragão BB, De Carvalho RG, Juliano MA, Frazzon J, et al. Diversity and emergence of multi-resistant Staphylococcus spp. isolated from subclinical mastitis in cows in of the state of Piauí, Brazil. Braz J Microbiol 2022;53(4):2215-2222. [ Links ]
39. Majumder S, Sackey T, Viau C, Park S, Xia J, Ronholm J, et al. Genomic and phenotypic profiling of Staphylococcus aureus isolates from bovine mastitis for antibiotic resistance and intestinal infectivity. BMC Microbiol 2023;23(1):43. [ Links ]
40. Yang F, Shi W, Meng N, Zhao Y, Ding X, Li Q. Antimicrobial resistance and virulence profiles of staphylococci isolated from clinical bovine mastitis. Front Microbiol 2023;14:1190790. [ Links ]
41. Martini CL, Lange CC, Brito MA, Ribeiro JB, Mendonça LC, Vaz EK. Characterisation of penicillin and tetracycline resistance in Staphylococcus aureus isolated from bovine milk samples in Minas Gerais, Brazil. J Dairy Res 2017;84(2):202-205. [ Links ]
42. Pérez VKC, Custódio DAC, Silva EMM, De Oliveira J, Guimarães AS, Brito MAVP, et al. Virulence factors and antimicrobial resistance in Staphylococcus aureus isolated from bovine mastitis in Brazil. Braz J Microbiol 2020;51(4):2111-2122. [ Links ]
43. Wang H, Zhuang H, Ji S, Sun L, Zhao F, Wu D, et al. Distribution of erm genes among MRSA isolates with resistance to clindamycin in a Chinese teaching hospital. Infect Genet Evol 2021;96:105127. [ Links ]
44. Anand T, Bera BC, Vaid RK, Barua S, Riyesh T, Virmani N, et al. Abundance of antibiotic resistance genes in environmental bacteriophages. J Gen Virol 2016;97(12):3458-3466. [ Links ]
Received: August 26, 2024; Accepted: May 12, 2025
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
