











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La mastitis bovina es la enfermedad infecciosa más prevalente a nivel mundial, la cual es asociada con severas pérdidas económicas debido a la reducción en la producción y calidad de la leche, así como el costo de los tratamientos y la salud de los animales, incrementando el riesgo de sacrificio y los costos de remplazo1-3. Staphylococcus aureus es reconocido como el principal patógeno a nivel mundial causante de mastitis bovina, y se considera a este microorganismo como contagioso y difícil de tratar debido a su alta tasa de diseminación entre animales infectados y la capacidad de ocasionar infecciones crónicas o recurrentes1,4,5.
La terapia con antibióticos es una importante medida para controlar la mastitis bovina. Sin embargo, se ha incrementado el número de estudios que reportan que S. aureus presenta resistencia a múltiples clases de antibióticos como respuesta a la presión selectiva por el uso continuo de los mismos, debido al sobreuso de estas sustancias activas en la medicina veterinaria y la agricultura6,7.
Además del uso de antibióticos para el control de la mastitis, es importante identificar los patrones epidemiológicos e identificar las fuentes de contaminación. Varias fuentes potenciales de contaminación se han identificado: desde el aire de los establos, el equipo de ordeña hasta la piel de la ubre8. Sin embargo, resultados acerca del papel de la piel de la ubre bovina como una fuente de S. aureus en las infecciones intramamarias aún no son concluyentes. Por un lado, se ha demostrado que las cepas de S. aureus de la piel y del canal de la ubre de la vaca pueden ser una fuente potencial para el desarrollo de la mastitis bovina, debido a que genéticamente son iguales a las cepas obtenidas en leche y que éstas pueden promover la cronicidad de la infección4,9. Por otro lado, en otro estudio se observó que la mayoría de los S. aureus asociados con casos de mastitis pertenecen a cepas altamente adaptadas a la glándula mamaria pero diferentes a las cepas que provienen de piel8. Aun cuando no es claro el papel de las cepas aisladas de la piel de la ubre de la vaca, estas cepas pueden fácilmente transferirse a la leche cruda durante la ordeña. La leche bronca contaminada con S. aureus puede convertirse en un problema de salud pública, debido al riesgo de una intoxicación alimentaria por este microorganismo; además de la diseminación de cepas resistentes a antibióticos4,10.
En un estudio previo, se aislaron cepas de S. aureus de la piel de la ubre de vacas de tres establos lecheros del suroeste de México, estas cepas presentaron los genes de la enterotoxina A, D y E. Además, dentro de estas cepas se identificaron grupos clonales que se diseminaron entre los establos y permanecían en temporadas de lluvias y estiaje11. Por el potencial toxigénico de las cepas, es importante continuar con la determinación de otras características de importancia epidemiológica como es la resistencia a antibióticos, considerando que su persistencia en los establos puede estar relacionada a la elevada resistencia a antibióticos. Por lo tanto, el objetivo de este estudio fue determinar la resistencia a antibióticos y genes asociados en cepas de S. aureus aisladas de la piel de ubre de bovinos en granjas del suroeste de México.
Material y métodos
Cepas bacterianas
Las cepas de S. aureus provenientes del pezón de la ubre de vaca se aislaron durante el año 2019, en dos temporadas del año (lluvias y estiaje) y en tres establos lecheros ubicados en tres zonas con diferente altitud11. Las cepas están conservadas en crioviales de glicerol al 15 % (v/v) a -20 °C en el cepario del Laboratorio de Investigación de Patometabolismo Microbiano (LIPM) de la Facultad de Ciencias Químico-Biológicas de la Universidad Autónoma de Guerrero. Estas cepas fueron caracterizadas bajo el siguiente perfil fenotípico: coco Gram positivo, manitol, catalasa y coagulasa positivos e identificados molecularmente por el gen fem. La reactivación de las cepas se realizó resuspendiendo 20 µl de cada criovial en 1 ml de caldo Mueller Hinton (MH), el cual se incubó a 37 °C por 24 h. Una vez obtenido el crecimiento, se inocularon en placas de agar BHI para su conservación como cepas de trabajo.
Concentración mínima inhibitoria
La resistencia antimicrobiana se evaluó mediante el ensayo de Concentración Mínima Inhibitoria (CMI); los inóculos bacterianos se prepararon a partir de cultivos de 24 h de cada una de las cepas en caldo MH y se ajustaron al 0.5 McFarland. En una microplaca de 96 pocillos se colocó cada antibiótico en diferentes rangos de concentraciones; penicilina (PEN 0.25-2 µg/ml), ceftriaxona (CRO 0.5-32 µg/ml), ciprofloxacino (CIP 4-32 µg/ml), clindamicina (DA 4-256 µg/ml), eritromicina (E 8-256 µg/ml), kanamicina (KAN 16-256 µg/ml), gentamicina (GEN 16-256 µg/ml), tetraciclina (TET 16-256 µg/ml), vancomicina (VAN 16-256 µg/ml), cloranfenicol (CLF 16-256 µg/ml) y trimetroprima (TMP 16-256 µg/ml) indicadas por el CLSI (Clinical and Laboratory Standards Institute)12. A continuación, se agregaron los inóculos bacterianos y se incubaron a 37°C durante 24 ± 1 h. Como control positivo se inocularon las mismas cepas sin antibiótico y como control negativo se utilizó un pocillo con caldo Mueller Hinton y antibiótico. Posteriormente, se realizó la lectura del crecimiento por absorbancia a una longitud de onda de 492/630 nm en un espectrofotómetro (Thermo Scientific®, GENESYS 200, USA). Después se calculó el CMI50 y se catalogaron las cepas de manera paramétrica y de acuerdo con los valores de CMI reportados por el manual del CLSI como Sensibles (S) o Resistentes (R).
Resistencia a betalactámicos
Cada una de las cepas se incubó 18 h en caldo MH a 37 °C en condiciones estáticas, el cultivo se ajustó al 0.5 McFarland y se inoculó en agar MH mediante el método de difusión en disco con discos de cefoxitina (FOX 30μg) y penicilina (PEN 10μg) y se incubó a 35 ± 2 °C durante 24 h12; una vez transcurrido el tiempo se midieron los halos de inhibición y se categorizaron como sensibles (S) (PEN ≥ 29 mm o FOX ≥25 mm) o resistentes (R) (PEN ≤28 mm o FOX ≤24 mm)12. Después se colocaron 10 μl de nitrocefina (Oxoid Thermo Scientific™) para determinar la producción de betalactamasas mediante la observación en el cambio de coloración del halo: un halo color rojizo permitió definirlas como productoras y cuando no se observaba un cambio en la coloración del halo como no productoras.
Determinación de fenotipos de resistencia a MLSB
Los fenotipos de resistencia a MLSB (Macrólidos-Lincosamidas-Streptogramina B) se determinaron mediante los resultados obtenidos de la resistencia a la clindamicina (C-R) o eritromicina (E-R), clasificando las resistencias obtenidas como constitutiva (C-R, E-R o ambas) o inducible (E-R, pero sensible a clindamicina)12.
Genes de resistencia a antibióticos
Extracción de DNA
El DNA genómico de todas las cepas de S. aureus se obtuvo a partir de 1 ml de un cultivo estático en caldo infusión cerebro corazón (BHI, por sus siglas en inglés: “brain heart infusion) de 18 h incubado a 37 °C. Las células se centrifugaron a 10,000 rpm por 10 min, resuspendiendo las pastillas en 300 μl de buffer de lisis (10 mM Tris HCl, 1 mM EDTA pH 8.0, lisozima 0.02 mg/ml) e incubando a 37 °C por 10 min. Posteriormente, se agregaron 200 μl de solución de fenol/cloroformo/alcohol isoamílico (25:24:1 v/v/v), se mezcló por inversión 10 a 15 veces, se centrifugó nuevamente a 10,000 rpm por 10 min, se recuperaron 200 μl de la fase acuosa y se transfirieron a 1 ml de etanol frío (96%). Se incubó a -20 °C durante 24 h, después se centrifugaron a 10,000 rpm por 10 min, se desechó el sobrenadante, las pastillas celulares se dejaron secar a temperatura ambiente y se resuspendieron en 20 μl de agua estéril13.
Genes que confieren resistencia a antibióticos
La identificación de los genes que confieren resistencia a antibióticos se determinó por reacción en cadena de la polimerasa (PCR) en punto final. Para los genes mecA, blaZ, ermA-C, msrA y tetM se utilizó la siguiente mezcla de reacción: 1X Taq 2x Máster Mix RED, 1.5 MgCl2 (Ampliqon, Denmark), 40 nM de cada oligonucleótido y 100 ng de DNA. En el caso de la ermB se utilizó la siguiente mezcla de reacción 1X de Taq 2x Máster Mix RED, 1.5 MgCl2 (Ampliqon, Denmark), 160 nM de cada oligonucleótido y 100 ng de DNA. Se utilizó la cepa ATCC 43300 S. aureus subsp. aureus rosenbach para ermA, mecA, blaZ; para el gen ermB se utilizó un aislado clínico de S. aureus caracterizado en el LIPM; la cepa ATCC 25923 S. aureus subsp. aureus rosenbach y la cepa 1084 de S. hominis caracterizada por el Laboratorio de Investigación en Microbiología de la Facultad de Ciencias Químico Biológicas de la Universidad Autónoma de Guerrero, se utilizaron como controles de los genes tetM y msrA respectivamente. Los oligonucleótidos y las condiciones se describen en el Cuadro 1.
Cuadro 1 Oligonucleótidos utilizados en este estudio
| Gen | Secuencia | Ciclaje | Tamaño esperado (pb) | Referencia |
|---|---|---|---|---|
| blaZ | F-ATTTTGAAAAAGTTAATATTTTTAATTG R-CATTACACTCTTGGCGGTTTC |
1 ciclo 95°C por 5 min; 35 ciclos a 95°C por 30 s, 52°C por 50 s, y 72°C por 50 s; y 1 ciclo a 72°C por 10 min |
833 | 14 |
| mecA | F- AAAATCGATGGTAAAGGTTGGC R- AGTTCTGCAGTACCGGATTTGC |
1 ciclo 94°C por 5 min; 32 ciclos 94°C por 30 s, 57°C por 45 s, y 72°C por 1 min; y 1 ciclo 72°C por 5 min |
583 | 15 |
| ermA | F-GTTCAAGAACAATCAATACAGAG R-GGATCAGGAAAAGGACATTTTAC |
1 ciclo 94°C por 2 min; 32 ciclos 94°C por 30 s, 52°C por 1 min, y 72°C por 30s; y 1 ciclo 72°C por 10 min |
421 | 16 |
| ermB | F-CCGTTTACGAAATTGGAACAGGTAAAGGGC R- GAATCGAGACTTGAGTGTGC |
1 ciclo 95°C por 5 min; 32 ciclos 95°C por 20 s, 52°C por 45 s, y 72°C por 40 s; y 1 ciclo 72°C por 7 min |
359 | |
| ermC | F- RGCTAATATTGTTTAAATCGTCAATTCC R-GGATCAGGAAAAGGACATTTTAC |
1 ciclo 94°C por 2 min; 32 ciclos 94°C por 30 s, 52°C por 1 min, y 72°C por 30s; y 1 ciclo 72°C por 10 min |
572 | |
| msrA | F-GGCACAATAAGAGTGTTTAAAGG R-AAGTTATATCATGAATAGATTGTCCTGTT |
1 ciclo 95°C por 15 min; 32 ciclo 95°C por 30 s, 52°C por 45 s, y 72°C por 1 min; y 1 ciclo 72°C por 7 min |
940 | 17 |
| tetM | F- GTGGACAAAGGTACAACGAG R-CGGTAAAGTTCGTCACACAC |
1 ciclo 95°C por 5 min; 32 ciclo 95°C por 20 s, 52°C por 45 s, y 72°C por 40 s; y 1 ciclo 72°C por 7 min |
406 | 16 |
Resultados
Resistencia a antibióticos
Se determinó que el 95.1 % (58/61) de las cepas son resistentes a penicilina y el 100 % (61/61) son resistentes a ceftriaxona. Solo se encontró el 1.6 % de las cepas (1/61) con resistencia a ciprofloxacino, gentamicina y cloranfenicol, 98.4 % (60/61) a clindamicina, 42.6 % (26/61) a eritromicina, 62.3 % (38/61) a kanamicina, 22.9 % (14/61) a tetraciclina, 13.1 % (8/61) a trimetoprima. Por último, no se encontraron cepas resistentes a vancomicina.
En cuanto a los resultados de las concentraciones de antibiótico a las cuales son inhibidas cada una de las cepas analizadas en este trabajo. Las 58 cepas con resistencia a penicilina se distribuyeron en CMI de 0.25 a 2.0 µg/ml; en el caso de la ceftriaxona, en las 61 cepas resistentes, se determinaron CMI de 0.25 µg/ml hasta mayores a 32 µg/ml, de las cuales 40 cepas presentaron la CMI más alta (32 µg/ml).
En el grupo de antibióticos MLSB, las cepas resistentes para clindamicina (60/61) presentaron CMI de 8 a 256 µg/ml, siendo 44 cepas las que presentaron CMI de 256 µg/ml, para eritromicina las cepas resistentes tienen una CMI de 16 a 256 µg/ml, de las cuales cuatro cepas tenian CMI de 256 µg/ml; del grupo de fluoroquinolonas solo una cepa con CMI de 4 µg/ml de ciprofloxacino, en aminoglucósidos se determinaron cepas con CMI de 32 a 256 µg/ml de concentración de kanamicina y una cepa con CMI de 64 µg/ml de gentamicina.
Respecto a tetraciclina, se observaron cepas con CMI de 32 a 256 µg/ml, de las cuales 11 cepas tenian CMI de 256 µg/ml; en anfenicoles solo se obtuvo una cepa con CMI de 128 µg/ml de cloranfenicol; por último, en el grupo de las diaminopirimidinas se determinaron valores de 32 a 256 µg/ml en cepas con resistencia a trimetoprima, observando dos cepas con CMI de 256 µg/ml (Cuadro 2).
Cuadro 2 Concentración mínima inhibitoria de las cepas de S. aureus resistentes a los diferentes grupos de antibióticos
| Antibióticos | Concentraciones utilizadas (µg/ml) | Cepas resistentes % (n), N= 61 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | 256 | ||
| Betalactámicos | ||||||||||||
| Penicilina | 9 | 5 | 17 | 27 | 95.1 (58) | |||||||
| Cefalosporinas | ||||||||||||
| Ceftriaxona | 1 | 5 | 1 | 10 | 4 | 0 | 40 | 100 (61) | ||||
| Fluoroquinolonas | ||||||||||||
| Ciprofloxacino | 1 | 1.6 (1) | ||||||||||
| Lincosamidas | ||||||||||||
| Clindamicina Macrólidos | 10 | 0 | 0 | 4 | 2 | 44 | 98.4 (60) | |||||
| Eritromicina | 11 | 9 | 0 | 2 | 4 | 42.6 (26) | ||||||
| Aminoglucosidos | ||||||||||||
| Kanamicina | 10 | 9 | 7 | 12 | 62.3 (38) | |||||||
| Gentamicina | 1 | 1.6 (1) | ||||||||||
| Tetraciclinas | ||||||||||||
| Tetraciclina | 1 | 0 | 2 | 11 | 22.9 (14) | |||||||
| Fenicoles | ||||||||||||
| Cloranfenicol | 1 | 1.6 (1) | ||||||||||
| Diaminopirimidinas | ||||||||||||
| Trimetoprima | 4 | 0 | 2 | 2 | 13.1 (8) | |||||||
Los parámetros de la CMI (µg/ml) se determinaron de acuerdo con los valores establecidos en la 31ed M100 Performance Standards for Antimicrobial Susceptibility Testing12.
Fenotipos de resistencia a antimicrobianos
El 100 % de las cepas fueron resistentes a cefoxitina y solo una fue productora de betalactamasas (1.6 %). Para eritromicina y clindamicina se obtuvieron frecuencias de resistencia del 42.6 y 98.7 % respectivamente, las cuales se categorizaron dentro del grupo de fenotipos de resistencia constitutiva a MLSB, destacando que el mismo 42.6 % de cepas fue resistente a ambos antibióticos (eritromicina y clindamicina). No se determinaron cepas con resistencia a eritromicina, pero sensibilidad a clindamicina, por lo que no existe una resistencia inducible de un antibiótico; en cambio se encontró un resultado atípico de 55.8 % de cepas con resistencia a clindamicina (C-R), sin ser resistentes a eritromicina (S). El 80.2 % de las cepas presentan resistencia al menos a tres grupos de antibióticos (Cuadro 3).
Cuadro 3 Fenotipos de resistencia antibióticos en las cepas de S. aureus
| Fenotipo de resistencia a betalactámicos | Número de cepas n (%) N=61 |
|---|---|
| MRSA | 61 (100.0) |
| MSSA | 0 (0.0) |
| Producción de betalactamasas | 1 (1.6) |
| Fenotipos MLSB | (%) |
| cMLSB | |
| E- R | 26 (42.6) |
| C- R | 60 (98.7) |
| E-R, C-R | 26 (42.6) |
| iMLSB | |
| Fenotipos de resistencia a antibióticos | (%) |
| PEN, CRO | 1 (1.6) |
| PEN, CRO, DAN | 11 (18.2) |
| PEN, CRO, DAN, E | 3 (4.9) |
| PEN, CRO, DAN, E, KAN | 13 (21.3) |
| PEN, CRO, DAN, E, KAN, TET | 4 (6.7) |
| PEN, CRO, DAN, E, KAN, CIP | 1 (1.6) |
| PEN, CRO, DAN, E, KAN, TMP | 1 (1.6) |
| PEN, CRO, DAN, E, TET | 1 (1.6) |
| PEN, CRO, DAN, E, TMP | 1 (1.6) |
| PEN, CRO, DAN, E, TMP, CLF, TET | 1 (1.6) |
| PEN, CRO, DAN, KAN | 14 (23.1) |
| PEN, CRO, DAN, KAN, GEN, TET | 1 (1.6) |
| PEN, CRO, DAN, KAN, TET | 3 (4.9) |
| PEN, CRO, DAN, TMP | 3 (4.9) |
| PEN, CRO, DAN, TMP, TET | 1 (1.6) |
| CRO, DAN, E | 1 (1.6) |
| CRO, DAN, E, KAN, TET | 1 (1.6) |
PEN (penicilina), CRO (ceftriaxona), DA (clindamicina), E (eritromicina), KAN (kanamicina), GEN (gentamicina), TMP (trimetoprima), TET (tetraciclina), CLF (cloranfenicol), CIP (ciprofloxacino), MRSA (methicilin resistance Staphylococcus aureus), MSSA (methicilin sensitive Staphylococcus aureus), MLSB (Macrólidos- Lincosamidas- Streptogramina B), cMLSB (resistencia constitutiva a Macrólidos- Lincosamidas- Streptogramina B), E-R (resistencia a eritromicina), C- R (resistencia a clindamicina), iMLSB (resistencia inducible a Macrólidos- Lincosamidas- Streptogramina B).
Genes de resistencia a antibióticos
Se determinaron frecuencias absolutas de los genes que confieren resistencia a cada uno de los grupos de antibióticos. De las cepas resistentes a betalactámicos solo se encontraron tres cepas positivas a la presencia del gen blaZ (4.91 %), una cepa resultó positiva la prueba de nitrocefina destacando que ésta no fue positiva a la presencia del gen blaZ. Para mecA, no se encontraron cepas positivas a la presencia de este gen (Cuadro 4) (Figura 1).
Cuadro 4 Distribución de genes de resistencia a antibióticos en cepas de S. aureus
| Cepas resistentes | Beta lactámicos | MLS | TETRA | |||||
|---|---|---|---|---|---|---|---|---|
| Grupo de antibióticos | Antibiótico | blaZ | mecA | ermA | ermB | ermC | msrA | tetM |
| Betalactámicos | ||||||||
| Penicilina (N=58) | 3 | |||||||
| Cefalosporinas | ||||||||
| Ceftriaxona (N=61) | 3 | |||||||
| Cefoxitina (N=61) | 3 | |||||||
| ML | ||||||||
| Clindamicina/ Eritromicina (N=26) | 9 | 1 | 23 | |||||
| Clindamicina (N=60) | 1 | 1 | 29 | |||||
| TETRA | ||||||||
| Tetraciclina (N=14) | 9 | |||||||
TETRA= tetraciclina.
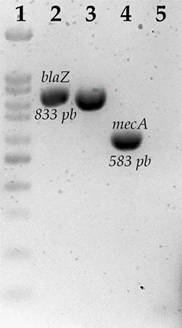
Electroforesis en geles de agarosa de la amplificación de blaZ y mecA. 1) Marcador de peso molecular de 100 pb; 2) blaZ, S. aureus ATCC43300; 3) blaZ, S. aureus S680; 4) mecA, S. aureus ATCC 43300; 5) mecA, S. aureus.
Figura 1 Genes asociados a resistencia a beta- lactámicos en cepas de S. aureus aisladas de piel de ubre bovina
Del grupo de antibióticos MLSB: se determinaron 52 cepas del total de cepas resistentes a MLSB con la presencia del gen msrA (86.6 %); de estos, 29 cepas pertenecen al perfil C-R (55.7 %) y 23 correspondían al perfil ER-CR (44.23 %). Además, se encontraron 10 cepas positivas al gen ermB (16.6 %), de las cuales sólo nueve corresponden al fenotipo ER-CR (90 %) y la cepa restante es del fenotipo C-R (10 %). También, se determinaron dos cepas con la presencia del gen ermC (3.3 %), de las cuales una cepa corresponde al fenotipo ER-CR y una cepa al fenotipo C-R y no hubo cepas positivas a ermA (Figura 2).
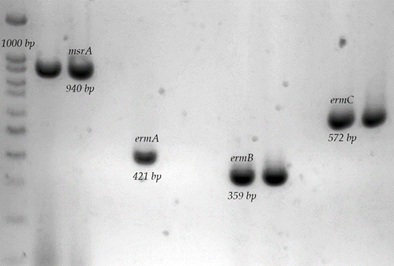
Electroforesis en geles de agarosa de la amplificación de los genes msrA, ermA- C. 1) Marcador de peso molecular de 100 pb; 2) S. haemolyticus 1035, msrA+; 3) S. aureus S697; 4) Control negativo; 5) S. aureus ATCC 43300, ermA+; 6) S. aureus S661; 7) Control negativo; 8) S. aureus S697, ermB+; 9) S. aureus S694, ermB+; 10) Control negativo; 11) S. aureus S662, ermC+; 12) S. aureus S661, ermC+.
Figura 2 Genes asociados a la resistencia a macrólidos en cepas de S. aureus aisladas de piel de ubre bovina
En la determinación del gen tetM de 14 cepas resistentes a tetraciclina, se determinaron nueve con la presencia del gen tetM (64.28 %) (Figura 3); de éstas, seis cepas presentaron CMI de 256 µg/ml, una cepa tuvo un CMI de 128 µg/ml y dos cepas con CMI de 32 µg/ml.
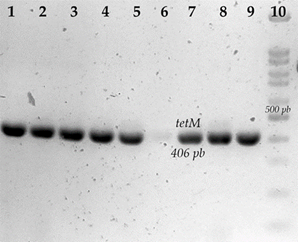
Electroforesis en geles de agarosa de la amplificación de tetM. 1) S. aureus S68; 2) S. aureus S689; 3) S. aureus S692; 4) S. aureus S693; 5) S. aureus S696; 6) S. aureus S697; 7) S. aureus S654; 8) S. aureus S662; 9) S. aureus D1; 10) marcador de peso molecular de 100 pb.
Figura 3 Genes asociados a resistencia a tetraciclina en cepas de S. aureus aisladas de piel de ubre bovina
Discusión
S. aureus es el principal patógeno a nivel mundial que causa mastitis bovina, caracterizándose por causar infecciones crónicas por la baja respuesta a antibióticos, facilitando la diseminación entre el ganado. La resistencia antibacteriana de este microorganismo surge por el extensivo uso de antibióticos para el control de la mastitis, convirtiéndose en un problema de salud pública debido a la aparición y diseminación de cepas resistentes a antibióticos a los humanos a partir de los alimentos18.
En este estudio, se encontró una alta resistencia a antibióticos betalactámicos en comparación a otros grupos de antibióticos en las cepas de S. aureus aisladas de la piel de la ubre bovina; la resistencia a estos antibióticos está mediada por la síntesis de proteínas alternas de unión a penicilina (PBP2a, PBP2´), síntesis de betalactamasas y mutaciones en los genes de las proteínas de unión a penicilina (PBP). La resistencia a cefoxitina infiere que el mecanismo podría estar mediado por la síntesis de proteínas alternas codificadas en el casete cromosomal SCCmec; sin embargo, en este estudio las cepas fueron negativas para la amplificación del gen mecA. Las cepas negativas para mecA pero con MIC elevados para oxacilina han sido reportadas como mecA -BORSA (S. aureus resistentes a oxacilina en el límite, mecA negativos) en distintos alimentos como pollo, puerco, leche, e incluso en la sociedad19 por lo que no es de extrañarse su presencia en la piel de ubre bovina. Una característica clave de las cepas BORSA, es la sobreproducción de betalactamasas20; en ese sentido, para evaluar la presencia de genes relacionados a betalactamasas se amplificó el gen blaZ, obteniendo una frecuencia baja para este gen (3/61, 4.91 %), la baja frecuencia de cepas productoras de betalactamasas codificadas por blaZ y negativas a mecA podría estar relacionada a la presencia de nuevas betalactamasas no codificadas por blaZ si no por plásmidos21. Por otro lado, para evaluar la producción de betalactamasas se valoró por la hidrolisis de nitrocefina, obteniendo una frecuencia baja (1/61, 1.64 %), remarcando que la cepa positiva a prueba de nitrocefina es negativa para el gen blaZ. En este punto, se han descrito hasta cuatro diferentes betalactamasas en S. aureus, denominadas de la A hasta la D, sin embargo, no todas se pueden determinar por la prueba de suspensión en nitrocefina, cefazolina o cefaloridina22. Aun cuando el principal mecanismo de resistencia en las cepas BORSA es la sobreproducción de betalactamasas, éste no es el único mecanismo; la presencia de nuevas beta-lactamasas, mutaciones en los genes de PBP o la presencia de bombas de eflujo podría explicar el comportamiento de estas cepas y podría estudiarse a futuro20,21,23.
Los macrólidos, lincosamidas y estreptograminas del grupo B denominados como MLSB se han convertido en una estrategia para el control de S. aureus resistentes a meticilina, así como en pacientes con alergia al grupo de betalactámicos. Del grupo de los MLSB, la clindamicina es de importancia debido a su aplicación para el tratamiento de infecciones de la piel, tejidos blandos y huesos, debido a su alta permeabilidad24-26. Con respecto a lo anterior, los granjeros que participaron en este estudio refieren el uso de este antibiótico como tratamiento preventivo de infecciones de la ubre bovina, lo cual se ve reflejado en la elevada resistencia a clindamicina (98.3 %, 60/61) en un fenotipo constitutivo con macrólidos (cMLSB) en el 42.6 % de las cepas. La resistencia a MLSB puede estar conferida por diferentes mecanismos como la modificación del sitio blanco, bombas de eflujo e inactivación enzimática del antibiótico27. La modificación del sitio blanco es mediada por la presencia de genes de metilasas de resistencia a eritromicina (erm). Los principales genes erm comúnmente encontrados incluyen ermA y ermC17. Los genes erm codifican metilasas que causan modificaciones conformacionales de la subunidad 23S RNAr, lo cual ocasiona una disminución en la unión de los antibióticos del grupo MLSB a su sitio blanco en la subunidad 50S ribosomal24. Uno de los datos relevantes de este estudio es la alta prevalencia del gen ermB en las cepas de S. aureus aisladas de ubre bovina, lo cual también se ha encontrado en cepas de Staphylococcus coagulasa positivos y negativos aislados de leche de búfalo28; a diferencia de cepas de origen clínico en donde es común encontrar los genes ermA y ermC29-31. Como se describe, la modificación del sitio blanco no es el único mecanismo que explica la resistencia constitutiva a MLSB; otro mecanismo es la presencia de bombas de eflujo como MsrA que explica la resistencia constitutiva a macrólidos32. La presencia de ambos mecanismos podría explicar en cierta medida el fenotipo constitutivo a MLSB (E-R, C-R) encontrado en este estudio, aun dejando a consideración la circulación de otros genes erm. En aquellas cepas de S. aureus que sólo son resistentes a clindamicina (55.8 %), aun cuando es un resultado poco común, éste podría explicarse por la presencia de genes linA y linB que inactivan solamente lincosamidas33 en las cepas circulantes y que podría ser una de las perspectivas a futuro de este estudio.
La tetraciclina es un antibiótico que en los últimos 40 años se sigue utilizando en la ganadería, humanos, animales pequeños, agricultura y acuacultura34. Al mismo tiempo, la tetraciclina no es solo usada para el tratamiento de infecciones en animales sino también como promotor de crecimiento o para mejorar la eficiencia de productos de engorda, práctica que en la actualidad se sigue realizando tanto en Estados Unidos de América como en otros países35. Por lo anterior, no es de sorprender la circulación de cepas de S. aureus con resistencia a tetraciclina provenientes de ganado bovino36 e incluso que S. aureus resistente a meticilina asociado a la ganadería (LA- MRSA) del complejo clonal CC9 tenga como característica distintiva la resistencia a tetraciclina37. En este sentido, se identificaron 14 cepas con resistencia a tetraciclina, remarcando que comparten otras características de resistencia bacteriana incluida la clindamicina, eritromicina y cefoxitina, por lo cual no se descarta que estas cepas también pertenezcan a este grupo clonal. Debido a esto, es necesaria en un futuro la caracterización completa de estas cepas. La resistencia a tetraciclina es mediada por genes que codifican bombas de eflujo así como proteínas de protección ribosomal35. En este último grupo, uno de los principales genes buscados es tetM. En este estudio, la frecuencia del gen tetM fue de 14.75 %, encontrándose en 9 de las 14 cepas resistentes. Este gen comúnmente se han reportado en cepas aisladas de mastitis subclínica en Brasil, China y Canadá38-40, muestras de leche de vaca en Brasil41,42, de leche de búfalo en Egipto28. En las cepas resistentes a tetraciclina pero negativas al gen tetM, la búsqueda podría enfocarse a más de 25 genes relacionados a la resistencia a tetraciclina34,35.
La determinación de los genes de resistencia en S. aureus así como en otros microorganismos es importante, debido a que muchos de estos genes se encuentran localizados en elementos genéticos móviles (EGM), los cuales, por mecanismos de transferencia horizontal pueden generar nuevas cepas resistentes a antibióticos. Incluso se han buscado estos EGM a partir de muestras ambientales debido a la importancia de estos43,44.
Conclusiones e implicaciones
Se confirmó en las unidades de producción lechera analizadas, la circulación de cepas de S. aureus provenientes de la piel de ubre bovina con resistencia a betalactámicos, macrólidos, lincosamidas y tetraciclinas con la presencia de los genes blaZ, ermABC, mrsA y tetM. Si bien, los presentes resultados no permiten generalizar, se sugiere que el adecuado seguimiento del desarrollo de casos de mastitis, la implementación de buenas prácticas de ordeño en ranchos de la región y el uso adecuado de los antimicrobianos, serían de ayuda para evitar la circulación de las cepas de S. aureus de la piel de la ubre bovina hacia la leche y en su momento hacia los humanos por el consumo de leche y sus derivados, favoreciendo la sanidad animal así como la salud humana y ambiental.

