










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.10 no.3 Texcoco abr./may. 2019 Epub 30-Mar-2020
https://doi.org/10.29312/remexca.v10i3.678
Essays
Volatile organic compounds of plants induced by insects: current situation in Mexico
1Departamento de Parasitología-Universidad Autónoma Agraria Antonio Narro. Calzada Antonio Narro núm. 1923, Buena Vista, Saltillo, Coahuila, México. CP. 25315. Tel. 01(844) 4110200. (cantuayala@hotmail.com).
2Centro de Investigación en Química Aplicada. Blvd. Enrique Reyna Hermosillo núm. 140, Saltillo, Coahuila, México. CP. 25294. Tel. 01(844) 4389830.
Plants produce and emit volatile organic compounds, generally they are mixtures that are composed of terpenes, fatty acid derivatives and aromatic compounds. Volatile organic compounds serve plants for: reproduction, attracting pollinators or seed dispersers, as a defense to repel insects or stop colonization by phytopathogenic bacteria and fungi, to attract natural enemies of herbivores and as intraspecific and interspecific messengers. Plants emit some volatiles especially when the vegetative parts are exposed to abiotic or biotic stimulation. Considering the foregoing, a bibliographic review was carried out, the purpose of which is to present the classification of volatile organic compounds in plants, to describe and indicate the characteristics of the extraction and identification techniques of volatile organic compounds, to present the advances reported in the scientific literature about these compounds induced by insects and their current situation in Mexico.
Keywords: interspecific communication; intraspecific communication; semiochemicals
Las plantas producen y emiten compuestos orgánicos volátiles, generalmente son mezclas que están compuestas por terpenos, derivados de ácidos grasos y compuestos aromáticos. Los compuestos orgánicos volátiles sirven a las plantas para: reproducción, atraer polinizadores o dispersores de semillas, como defensa para repeler insectos o detener la colonización por bacterias y hongos fitopatógenos, para atraer enemigos naturales de herbívoros y como mensajeros intraespecíficos e interespecíficos. Las plantas emiten algunos volátiles especialmente cuando las partes vegetativas se exponen a estimulo abiótico o biótico. Considerando lo antes expuesto, se realizó una revisión bibliográfica, siendo el objetivo del presente trabajo dar a conocer la clasificación de los compuestos orgánicos volátiles de las plantas, describir y señalar las características de las técnicas de extracción e identificación de los compuestos orgánicos volátiles, presentar los avances reportados en la literatura científica acerca de estos compuestos inducidos por insectos y su situación actual en México.
Palabras clave: comunicación interespecífica; comunicación intraespecífica; semioquímicos
General characteristics of volatile organic compounds of plants
Plants synthesize and emit a large variety of volatile organic compounds, which have important ecological functions. In most cases, at room temperature the volatile organic compounds of plants are gases, some are emitted by plants especially vegetative parts when they are exposed to biotic or abiotic stimulation (Scala et al., 2013; Dong et al., 2016). Volatile organic compounds are important for plants, as defense mechanisms to repel insects or stop colonization by pathogenic bacteria and fungi, to promote reproduction, by attracting pollinators or seed dispersers, they are intraspecific and interspecific messengers (Marín and Céspedes, 2007; Ángeles et al., 2012; Dicke et al., 2017).
In addition, these volatile organic compounds induced by herbivores, which consist of odors released by the plants, serve as important signals for parasitoids and predators to locate their host or prey (Ponzio et al., 2013; Schettino, 2017). They also influence the dynamics of the community (Figure 1), by inducing phytohormonal signaling with the subsequent arrival of other herbivores and, therefore, causing plant interactions with other members of the community (Dicke and Baldwin, 2010; Stam et al., 2014; Mofikoya et al., 2017). Some authors mention that volatile compounds act as a deterrent to feeding and ovipositing in insect pests (Ignacimuthu et al., 2011; Hijaz et al., 2016).
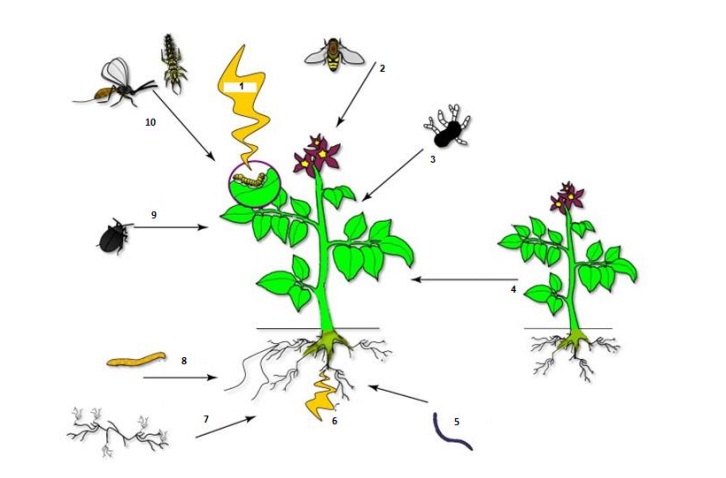
Figure 1 1) plant damaged by an herbivore emits semiochemicals, which interact with; 2) pollinator; 3) pathogen; 4) competitor or parasitic plant; 5) entomopathogenic nematode; 6) production of systemic semiochemicals; 7) mycorrhiza; 8 and 9) herbivorous; and 10) entomophages (modified from Dicke and Baldwin, 2010).
On the other hand, volatile organic compounds in ecosystems not only modulate the interactions between members of different trophic levels, but also play an important role in the chemistry of the atmosphere, herbivores induce a biochemical response at the site of damage (response local induced) and secondary metabolites induced in this site can migrate to other tissues of the plant and activate chemical resistance throughout the plant (induced systemic response) (Bautista et al., 2012a).
Volatile organic compounds are produced and emitted by leaves, fruits, flowers and roots, but they are mainly released; Throughout, the surface and storage sites in the leaves are influenced by environmental factors such as temperature, humidity and nutrient availability (Sharma et al., 2012; Kallenbach et al., 2014).
With the understanding of the role that semiochemicals play in the ecology and mechanisms of emission of volatile organic compounds from plants induced by herbivores, ways are opened to develop new strategies in biological and ethological control programs, with the aim of improving the effectiveness of natural enemies in the suppression of populations of pests in crops (Peñaflor and Bento, 2013). In addition, plants have developed perceptual skills that allow them to monitor and respond to a wide range of changing biotic and abiotic conditions, profoundly influencing their interactions with other organisms (Mescher and De Moraes, 2015).
Classification of volatile organic compounds of plants
Most volatile organic compounds in plants are products or byproducts of primary metabolic pathways. Based on their biosynthetic origin, volatile organic compounds are divided into four main metabolic pathways (Figure 2): 1) sikimate-phenylalanine; 2) mevalonic acid (MVA); 3) methyryltritol phosphate (MEP); and 4) of lipoxygenase (LOX), these pathways regulate the emission of benzenoid compounds, penilpropanoids, sesquiterpenes, monoterpenes, hemiterpenes, diterpenes, volatile carotenoid derivatives and methyl jasmonate, as well as green leaf volatiles (Dudareva et al., 2004; Dudareva et al., 2013).
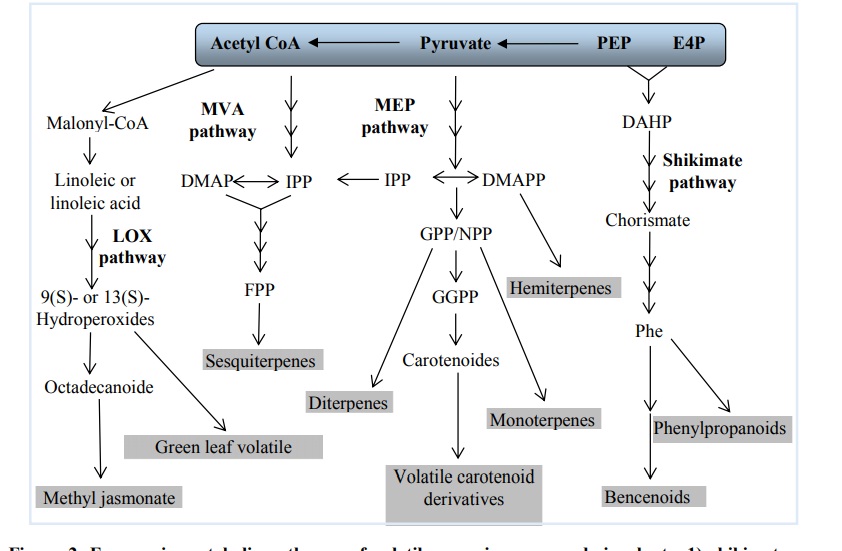
Figure 2 Four main metabolic pathways of volatile organic compounds in plants. 1) shikimate-phenylalanine; 2) mevalonic acid (MVA); 3) methylerythritol phosphate (MEP); and 4) lipoxygenase (LOX). The stacked arrows illustrate multiple enzymatic reactions. Abbreviations: DAHP, 3-deoxy-D-arabinoheptulosonate-7 phosphate; DMAPP, dimethylallyl pyrophosphate; E4P, erythrose 4-phosphate; FPP, farnesyl pyrophosphate; GGPP, geranylgeranyl pyrophosphate; GPP, geranyl pyrophosphate; IPP, isopentenyl pyrophosphate; NPP, neryl pyrophosphate; PEP, phosphoenolpyruvate; Phe, phenylalanine (modified from Dudavera et al., 2013).
Terpenes represent the most diverse group of volatile compounds, which are classified according to the number of carbons in their chain, including hemiterpenes (C5), monoterpenes (C10), sesquiterpene (C15), homoterpene (C11 and C16), some diterpenes (C20) and triterpene (C30), there are also volatile organic sulfur compounds such as dimethylsulfide and methanethiol (Heil and Karban, 2010; Scala et al., 2013; Vivaldo et al., 2017).
Kallenbach et al. (2014), conducted a classification of 38 volatile compounds for analysis in field experiments; It comprises green leaf volatiles: 3 (Z) -hexene-1-al, 2 (E) -hexene-1-al, 3 (Z) -hexene-1-ol, 2 (E) -hexene-1-ol, Hexan-1-ol, 3 (Z) -hexene-1-ol-acetate, 3 (Z) -hexenilisobutanoate, 3 (Z) -hexenylbutanoate, 3 (Z) -hexenyl-2-me-butanoate, 3 (Z) -hexenylvalerate, 3 (Z) -hexenyl- (E) -2-me-butenoate and 3 (Z) -hexenylcaproate; monoterpenes: α-pinene, β-myrcene, (Z) -β-ocimene, (E) -β-ocimene, linalool, α-terpinol and geraniol; sesquiterpenes: elemeno isomer, β-caryophyllene, α-dupreziane, (E) -α-bergamoteno, sesquiterpene 1, sesquiterpene 2, sesquiterpene 3, (E) -β-farnesene, sesquiterpene 4, 5’-epi-aristolochene, sesquiterpene 5, sesquiterpene 6, sesquiterpene 7, sesquiterpene 8, sesquiterpene 9 and sesquiterpene oxide; other compounds: benzyl alcohol, benzyl acetone and nicotine.
Some authors, such as Heil (2014), mention small molecules of C6, which participate in communication in the same plant and neighboring plants. Also, Zhang et al. (2015), classifies them in jasmonic acid, salicylic acid and ethylene signaling pathways. Martínez (2013), developed a nomenclature of semiochemicals based on two types: 1) pheromones, which mediate interactions between organisms of the same species and the interaction in general is beneficial for both the emitter and the receiver; and 2) allelochemicals: kairomonas, mediating interactions between different species, are the compounds that favor the receptor but not the emitter allomonas, are favorable to the emitter but not to the receiver and the sinomonas, which are favorable to both.
Extraction techniques and analysis of volatile organic compounds of plants
There are several methodologies for the extraction and analysis of volatile organic compounds obtained from plants, although most of the volatiles, for their quantitative and qualitative analysis, are obtained and analyzed by gas chromatography and coupled to mass spectrometry (CG-EM). (Moraes et al., 2008). Chaverri and Ciccio (2015), by means of this CG-EM technique, analyzed the chemical composition of the oils of leaves and fruits of wild pepper Pimenta guatemalensis (Lundell), obtaining β-caryophyllene, terpinolene, eugenol and sesquiterpenes.
Also, Michereff et al. (2011), by CG-EM determined high production of volatile compounds (E, E) -α-farnesene, methyl salicylate, (Z) -3-hexenyl acetate and (E) -2-octen-1-ol, in studies carried out on volatiles emitted by the damage of herbivores in resistant plants, compared to plants susceptible to soybean (Glycine max L.), also mentions that (E) -2-octen-1-ol, was not determined in susceptible plants. In this sense, Panighel and Flamini (2014), mention that the technique of CG-EM can be used to analyze volatiles such as monoterpenes and methoxypyrazines in grapes and wine, being a quick and simple technique.
Heredia et al. (2016), using this technique they identified 82 compounds in leaves of the plant species Zanthoxylum pistacifolium Griseb. Predominantly terpenoid derivatives and as major compounds α-pinene (12.35%), linalool (6.68%), 2,6-dimethyl-2, 4, 6-octatriene (6.50%), limonene (6.19%) and phytol (6.06%). A modification to the technique (CG-EM), is by soft chemical ionization mass spectrometry, this technique has been developed and used mainly by physicists and analytical scientists, who have used them in a wide range of scientific research areas for example: aroma, biomarkers of diseases, detection of dangerous compounds, atmospheric chemistry (Materic et al., 2015). Vidal et al. (2016), using this technique coupled to electroanthenographic (CG-EAG) performed the identification and synthesis of the sexual pheromone (Z) -trips-11-ene, in volatile released by male adults of the soldier beetle Chauliognathus fallax Germar.
Another modification to the CG technique is by solid phase microextraction coupled with gas chromatography and spectrometry (MEFS-CGEM), Raffo et al. (2018), by this technique identified the compounds (Z) - and (E) -3-hexenal, (Z) - 1,5-octadien-3-one, together with 4-mercaptobutyl and 4- (methylthio) butyl isothiocyanate in eruca leaves Eruca sativa L. and Diplotaxis tenuifolia L. Also Darshanee et al. (2017), through this technique they evaluated tomato plants (Solanum lycopersicum L.) to determine the chemical compounds that attract whitefly from greenhouses Trialeurodes vaporariorum (Westwood), identifying volatile (Z) -3-hexen-1-ol, a-pinene, (E) - β-caryophyllene, α-humulene and azulene. On the other hand, Souza et al. (2017), mention that this technique could open new perspectives and opportunities in agricultural and ecological studies, for the detection and identification of emissions of volatile organic compounds from plants induced by herbivores.
Application of volatile organic compounds of plants
The volatile organic compounds of plants play a preponderant role in the life cycle of plants, transmitting and providing information with the environment that surrounds them, possessing such diverse properties that generate complex interactive networks, with ecological, physiological and atmospheric implications (Marin and Cespedes, 2007). They could also be exploited as an important tool for the management of pests, to reduce the amounts of insecticides used for their control, the resistance of plants to insects, particularly the resistance induced, can also be manipulated with the use of chemical elicitors of metabolites secondary, which confer resistance to insects (Sharma et al., 2012).
They can also be exploited in the strategies of integrated pest management, such as increasing biological control in order to improve the control of pests in crops (Peñaflor and Bento, 2013). Tosh and Brogan (2015), mention that the use of volatile organic compounds of plants, could profoundly change the behavior of the insects that feed on them. In addition, the defense of plants against herbivores and pathogens involves the participation of a huge range of different metabolites, some of which act directly as defensive weapons against enemies (toxins or deterrents), for example, two well-studied groups of defensive weapons, are glucosinolates and benzoxazinoids, which trigger the accumulation of protective polysaccharides as a barrier against aphids and pathogens (Maag et al., 2014).
On the other hand, it has been seen that the accumulation of volatile secondary metabolites in the communication between plants and other organisms, represents an alternative for the sustainable protection of crops, the communication of plants has been extended to organisms antagonistic to pests and plant to plant (Pickett and Khan, 2016).
Other volatile compounds such as insect pheromones offer a potential to control crop pests, their volatility and instability are problems for deployment in agriculture, but could be solved by expressing genes that code for the biosynthesis of pheromones in crop plants, this has been achieved by genetic engineering of a hexaploid in a wheat variety Triticum aestivum L., to increase the release of (E) -β-farnesene (Eβf) to repel populations of aphids Sitobion avenae (Fabricius), Rhopalosiphum padi ( L.) and Metopolophium dirhodum (Walker) (Bruce et al., 2015).
Also salicylic acid induces resistance in plants against biotrophic pathogens and phloem-feeding insects, applications of salicylic acid could be used to repel, reduce fecundity, longevity and survival of the whitefly Bemisia tabaci (Gennadius) in crops, applications of salicylic acid also increase the amount and composition of volatiles released by plants, especially the amount of methyl salicylate and δ-limonene (Shi et al., 2016).
Bleeker et al. (2009), determined that the volatile sesquiterpene compounds zingiberene and curcumene, and the monoterpenes: p-cymene, α-terpinene and α-phellandrene, were involved in tomato and whitefly interactions; therefore, it may be feasible to alter the choice behavior of the white fly by manipulating the terpenoid composition. Du et al. (2016), evaluated seven volatile chemical repellents and their combinations to reduce the population densities of whitefly B. tabaci biotype Q, in tomato crop in greenhouses, the mixture of DLCO (D-limonene, citral and olive oil (63:7:30) had a better yield (SC50= 22.59 mg mL-1) to repel white flies and lower cost due to the lower price of olive oil than the other chemicals used and point to it as a new repellent with potential for the control of white flies.
Laredo et al. (2017), mentions jasmonic acid inducing local and systemic resistance to diseases and is expected to develop a new and effective pest control agent that will help the plant defend itself against its enemies; however, a series of laboratory and field investigations are still required to understand the biological behavior of this acid. Züst and Agrawal (2016), in recent studies, included the identification of salivary proteins of aphids involved in the manipulation of host plants and plant receptors involved in the recognition of aphids.
In studies conducted by Mofikoya et al. (2017), the results indicate that passive adsorption and re-release of a volatile monoterpene can occur in nature; however, this process depends on the distance between the emitting source and the receiving plants, as well as the concentration of air pollutants in the air. Therefore, in the field use of volatile plant compounds in modern pest control, the effects of distances and air pollution should be considered.
Study of the volatile organic compounds of plants in Mexico
The study of the volatile organic compounds of the plants in the international scope began in the decade of the eighties, in Mexico practically started in the new millennium. The literature reported works in different areas, in this review some works made in the country are mentioned below. Calyecac et al. (2006), in tests of attraction in the laboratory and greenhouse indicate that the combination of the volatile attractants released by the male weevil of the peeled tomato Trichobaris championi Barber., more those released by its host tomato shell Physalis ixocarpa Brot., provide the best attraction stimulus for the peeled tomato weevil.
Also in peeled tomato Physalis philadelphica Lam., using MEFS-CGEM, volatile methyl salicylate, β-caryophyllene, β-pinene, 3-carene, benzyl alcohol, β-elemene, decanal and germecrene D (Calyecac et al., 2007).
On the other hand, in wild creole avocado Persea americana var. drymifolia (Schltdl et Cham.) S. F. Blake, the foliar chemical profile was determined by gas chromatography coupled to mass spectrometry (CG-EM), the most abundant compounds were estragole (22 to 72%) and caryophyllene (9 to 16%), this chemical variation could be important to identify trees with relevant chemical phenotypes and as a source of resistance genes against insect pests and diseases of commercial varieties of Persea americana Mill. (Torres et al., 2009). Likewise, extraction of volatile compounds of basil (Ocimum basilicum L.) was carried out by (MEFS) and analyzed in (CG-EM), 25 volatile compounds were identified as phenylpropanoids, monoterpenes, sesquiterpenes, esters and aldehydes (González et al., 2011).
Studies conducted by Angeles et al. (2012), detected a total of 36 different volatile organic compounds, 23 terpenoids, decanal, dean and methyl salicylate; produced by tomato plants infested with whitefly from greenhouses. It is also analyzed the volatile components associated with the aroma of chili pepper (Capsicum annuum L. var. glabriusculum), by (MEFS) and analyzed in (CG-EM); the analysis identified ester compounds, sesquiterpenes, alcohols and alkanes (Vázquez et al., 2015).
Bautista and Espinosa (2013), evaluated the variation of emissions of volatile organic compounds or uniformity among tomato individuals in response to different damages; the identification of the volatiles was by (CG-EM); eleven volatile compounds were identified: 3-hexanol, α-pinene, o-cymeno, β-myrcene, (+) - 4-carene, α-phellandrene, α-terpinene, p-cymeno, β-phellandrene, nonanal and decanal. On the other hand, individual volatile organic compounds such as limonene, linalool, nonanal, methyl salicylate and methyl jasmonate, at natural concentrations directly inhibited the germination of conidia of the fungus Colletotrichum lindemuthianum (Sacc. & Magnus) Briosi & Cavara, in bean Phaseolus vulgaris L., were analyzed by gas chromatography coupled to electronic ionization mass spectrometry (CG-EMIES) (Quintana et al., 2014).
The lima bean (Phaseolus lunatus L.) has been used to study the interaction between plants, ants and pollinators and their consequences for the reproduction of the plant, two field experiments were carried out in which the presence of ants and production was manipulated of nectar by induction with jasmonic acid (JA), the plants induced by JA without ants produced a greater number of seeds in comparison with the plants treated with JA with ants (Hernandez et al., 2016). In plants of cultivated jicama Pachyrhizus erosus L. and wild P. ferrugineus (Piper) Sorenson, volatile organic compounds were identified by (MEFS) and analyzed in (CG-EM), in P. erosus 21 compounds were found, being the most abundant: cyclohexanone (32.8%), 3-hexen-1-ol (Z) (32.7%) and 3-hexenal (Z) (10.5%), in P. ferrugineus 11 compounds were found, the most abundant were: 5-hexen-1-ol acetate (51.5%), undecanal (22.4%) and 2-hepten-1-al (14.5%) (Estrella et al., 2016).
Bautista et al. (2017), isolated volatile and non-volatile compounds of mistletoe cactus Rhipsalis baccifera (JS Muell.) Stearn, by extraction in organic solvent, hydrodistilation, dynamic aeration, were identified and analyzed by MEFS, metabolites were detected as ketones, alcohols, alkanes, monoterpenes, aromatic compounds and an ester. In another study they analyzed the essential oil of the salvia aerial parts Salvia ballotiflora Benth. Using (CG-EM), it was determined that it possesses insecticidal properties against the corn leafworm Spodoptera frugiperda Smith. Being β-caryophyllene and the oxide of caryophyllene the main components (Cárdenas et al., 2015).
In addition, the composition of odors with the content of phenoloxidase (PO) in honeybees of the Euglossa imperialis Cockerell orchids and the variation in body size between individuals was analyzed, the analysis of the odor composition was by (CG-EM), 60 compounds were determined in one hundred males of E. imperialis, the most represented compounds were tetradecyl tetradecanoate (87%), β-cubebeno (76%), Z, E-2,13-octadecadien-1-ol (65%) and eicosano (61%) (Arriaga et al., 2017).
Currently in the laboratory is a study on the determination of volatile compounds by the method of MEFS, in soybean plants in response to the attack of whitefly, the analysis of the compounds will be made by means of CG-EM, as part of the Postgraduate Program in Agricultural Parasitology, of the Autonomous Agrarian University Antonio Narro. The study mentioned, is focused on the main biotic factors that limit the production of soybean in Mexico, such as insect pests and diseases; among the former the whitefly is one of the most important, in 1994 it reduced the yield from 2.1 to 1.5 t ha-1 in 200 thousand hectares in northwestern Mexico (Ochoa et al., 2011). For the control of this insect, chemical insecticides and some biorationals are currently used, such as soaps and vegetable extracts (Cortez et al., 2005).
Perspectives of volatile organic compounds of plants in Mexico
The use of volatile plant compounds technology as an additional tool in integrated pest management programs offers a new and environmentally sound approach to crop protection. This technique involves the development of baits that attract beneficial organisms and the manipulation of biochemical processes that induce and regulate plant defenses, key factors in the improvement of control programs against economically important pests (Arab and Bento, 2006). Volatile plants can also be used to protect against pathogens and insect pests.
It is sought in the future to increase the expression of genes involved in the biosynthesis of volatile and that volatiles can be stored and released at the time of damage, to be used as antimicrobial compounds or herbivore repellents, this would greatly increase the protection of plants in their natural environment and would provide an alternative natural pest control (Marín and Céspedes, 2007).
One of the key points in chemical ecology has been to include the resistance of the plant as a key factor to understand how the interactions between different trophic levels work and evolve. However, it is now necessary to include in this scheme the agronomic management, to know and manage the environment in which the interactions between plant-herbivore-entomophagous are developed (Bautista et al., 2012b). For further studies, the study of the role that volatile compounds develop in tritrophic interactions (plant-herbivore-entomophagous) raises more questions about the ecological and evolutionary processes that have modulated the interactions that are established between plants and organisms of different trophic levels (Bautista et al., 2012a).
Given that volatile organic compounds of plants are relatively difficult to identify, one of the great challenges is to isolate and characterize, he or those that modulate interactions of interest between two or more organisms (Martínez, 2013). In this sense, the direct defensive attributes of volatile organic compounds in plants provide a useful framework for the design of biological control strategies that can allow the sustainable and profitable protection of our crops (Stenberg et al., 2015).
Meiners (2015) suggests that studying multitrophic interactions under more realistic conditions will help to identify mechanisms with evolutionary potential and patterns that can be used in the practice of biological control. Likewise, Gish et al. (2015), mention that there are questions about evolutionary and ecological functions remain unanswered. An important point is the lack of information on the ways in which environmental factors, including habitat and atmospheric conditions influence the volatiles of plants, understanding these influences has significant implications for the sustainable management of natural and agricultural ecosystems and should be a priority for future research. Rodrígues et al. (2014), indicate that when genes responsible for the biosynthesis of volatiles induced by herbivores are identified, it will be possible to use genetic engineering to induce the production of specific compounds or mixtures of compounds in crops that do not produce naturally or to increase the flow of these compounds.
In Mexico, there are currently groups of researchers working on the isolation, determination and application of volatile organic compounds of plants, although there is no central research group that works on the potential exploitation of plant-herbivore-entomophagous interactions, for that in the future there are biocontrol strategies in the main agricultural crops. Studies on volatile compounds of plants, would contribute to reduce the impact of insecticides on ecosystems and reduce the costs of crop production, improving agricultural productivity and with it the socioeconomic conditions in general of agricultural producers.
As knowledge about tritrophic interactions increases, one of the biggest challenges is how to exploit the volatile organic compounds information from plants to develop new strategies for pest control. It is necessary to carry out more studies to identify the volatile organic compounds that govern the behavior directed by the smell of insect pests and their natural enemies, to formulate strategies for their control, for the development of varieties with constitutive and induced resistance to insect pests, also the manipulation of said volatiles to attract natural enemies of crop pests, to improve the efficiency of biological control agents in the management of pests.
Conclusions
Volatile organic compounds in plants are products or byproducts of primary metabolic pathways. Based on their biosynthetic origin, volatile organic compounds are divided into four main metabolic pathways: shikimate-phenylalanine, mevalonic acid (MVA), methyleritritol phosphate (MEP) and lipoxygenase (LOX). In the international context and in Mexico, the extraction and identification of the volatile organic compounds of the plants, is carried out mainly by microextraction in solid phase and gas chromatography and spectrometry (MEFS-CGEM).
In Mexico, there are currently groups of researchers in universities and research centers, working on the determination and application of volatile organic compounds of plants, some of them focused on the protection of crops against fungal diseases, although there is no group Research center working on issues related to plant-herbivore-entomophagous interactions, which represents a great potential for agriculture as a biocontrol strategy.
Literatura citada
Ángeles, L. Y. I.; Martínez, G. N. A.; Ramírez, R. R.; López, M. G.; Sánchez, H. C. and Délano, F. J. P. 2012. Cross-Kingdom effects of plant-plant signaling via volatile organic compounds emitted by tomato (Solanum lycopersicum) plants infested by the greenhouse whitefly (Trialeurodes vaporariorum). J. Chem. Ecol. 38(11):1376-1386. [ Links ]
Arab, A. and Bento, J. M. S. 2006. Plant volatiles: new perspectives for research in Brazil. Neotropical Entomol. 35(2):151-158. [ Links ]
Arriaga, O. B. J.; Contreras, G. J.; Espinosa, G. F. J.; García, R. Y. M.; Moreno, G. M.; Lanz, M. H.; Godínez, A. H. and Cueva, D. R. 2017. Are body size and volatile blends honest signals in orchid bees? Ecol. Ev. 7(9):3037-3045. [ Links ]
Bautista, L. A. and Espinosa, G. F. J. 2013. Odor Uniformity among tomato individuals in response to herbivore depends on insect species. PLOS ONE. 8(10):1-12. [ Links ]
Bautista, L. A.; Bravo, M. A. E. y Espinosa, G. F. J. 2012a. Importancia ecológica de la emisión de compuestos volátiles vegetales. 11. In: temas selectos en ecología química de insectos. Rojas, J. C. y Malo, E. A. (Eds.). El Colegio de la Frontera Sur. México. 268-286 pp. [ Links ]
Bautista, L. A.; Parra, R. F. y Espinosa, G. F. J. 2012b. Efectos de la domesticación de plantas en la diversidad fitoquímica. 10. In: temas selectos en ecología química de insectos. Rojas, J. C. y Malo, E. A. (Eds.). El Colegio de la Frontera Sur. México. 253-267 pp. [ Links ]
Bautista, S. J. A.; Cibrián, T. J.; Salomé, A. L. F.; Soto, H. R. M. y De la Cruz, D. E. 2017. Composición química del aroma de tallos y frutos de Rhipsalis baccifera (J. Miller) Stearn. Rev. Fitotec. Mex. 40(1):45-54. [ Links ]
Bleeker, P. M.; Diergaarde, P. J.; Ament, K.; Guerra, J.; Weidner, M.; Schutz, S.; de Both, M. T. J.; Haring, M. A. and Schuurink, R. C. 2009. Plant Physiol. 151(2):925-935. [ Links ]
Bruce, T. J. A.; Aradottir, G. I.; Smart, L. E.; Martin, J. L.; Caulfield, J. C.; Doherty, A.; Sparks, C. A.; Woodcock, C. M.; Birkett, M. A.; Napier, J. A.; Jones, H. D. and Pickett, J. A. 2015. The first crop plant genetically engineered to release an insect pheromone for defence. Sci. Reports. 11:1-9. [ Links ]
Calyecac, C. H. G.; Cibrián, T. J.; López, C. J. y García, V. R. 2006. Emisores de los volátiles de atracción de Trichobaris championi Barber. Agrociencia. 40(5):655-663. [ Links ]
Calyecac, C. H. G.; Cibrián, T. J.; Soto, H. M. y García, V. R. 2007. Aislamiento e identificación de volátiles de Physalis philadelphica Lam. Agrociencia. 41(3):337-346. [ Links ]
Cárdenas, O. N. C.; González, C. M. M.; Figueroa, B. R.; Flores, M. A.; Romo, A. D.; Martínez, G. D. E.; Pérez, M. V. and Ramos, L. M. A. 2015. Composition of the essential oil of Salvia ballotiflora (Lamiaceae) and its insecticidal activity. Molecules. 20(5):8048-8059. [ Links ]
Chaverri, C. and Cicció, J. F. 2015. Leaf and fruit essential oil compositions of Pimenta guatemalensis (Myrtaceae) from Costa Rica. Rev. Biol. Trop. 63(1):303-311. [ Links ]
Cortez, M. E.; Rodríguez, C. F. G.; Martínez, C. J. L. y Macías, C. J. 2005. Tecnología de producción y manejo de la mosca blanca de la hoja plateada en el cultivo de soya en el Norte de Sinaloa. INIFAP-CIRNO-CEVAF. Los Mochis, Sinaloa, México. Folleto técnico núm. 25. 52 p. [ Links ]
Darshanee, H. L. C.; Ren, H.; Ahmed, N.; Zhang, Z. F.; Liu, Y. H. and Liu, T. X. 2017. Volatile mediated attraction of greenhouse whitefly Trialeurodes vaporariorum tomato and eggplant. Frontiers Plant Sci. 8:1-13. [ Links ]
Dicke, M. and Baldwin, I. T. 2010. The evolutionary context for herbivore-induced plant volatiles: beyond the ‘cry for help’. Trends Plant Sci. 15(3):167-175. [ Links ]
Dong, F.; Fu, X.; Watanabe, N.; Su, X. and Yang, Z. 2016. Recent advances in the emission and functions of plant vegetative volatiles. Molecules. 21(2):1-10. [ Links ]
Du, W.; Han, X.; Wang, Y. and Qin, Y. 2016. A primary screening and applying of plant volatiles as repellents to control whitefly Bemisia tabaci (Gennadius) on tomato. Sci. Reports. 6:1-10. [ Links ]
Dudareva, N.; Klempien, A.; Muhlemann, J. K. and Kaplan, I. 2013. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 198(1):16-32. [ Links ]
Dudareva, N.; Pichersky, E. and Gershenzon, J. 2004. Biochemistry of plant volatiles. Plant Physiol. 135(4):1893-1902. [ Links ]
Estrella, P. J. C.; Gómez, V. J. C.; Céspedes, C. L.; Alarcón, J.; Cano, S. Z.; Reyes, G. A.; Cerbón, C. M. A. and Reyes, Ch. R. 2016. Volatile organic compounds from Pachyrhizus ferrugineus and Pachyrhizus erosus (Fabaceae) leaves. Boletín Latinoamericano y del Caribe de Plantas Medicinales y Aromáticas. 15(3):175-181. [ Links ]
Gish, M.; De Moraes, C. M. and Mescher, M. C. 2015. Herbivore-induced plant volatiles in natural and agricultural ecosystems: open questions and future prospects. Insect Sci. 9:1-6. [ Links ]
González, Z. J. A.; González, S. H. M.; González, P. S.; Rosales, R. T. y Andrade, G. I. 2011. Microextracción en fase sólida de compuestos volátiles en albahaca (Ocimum basilicum L.). Revista Acta Universitaria. Universidad de Guanajuato. 21(1):17-22. [ Links ]
Heil, M. 2014. Herbivore-induced plant volatiles: targets, perception and unanswered questions. New Phytol. 204(2):297-306. [ Links ]
Heil, M. and Karban, R. 2010. Explaining evolution of plant communication by airborne signals. Trends Ecol. Evol. 25(3):137-144. [ Links ]
Heredia, D. Y.; González, F. R.; Escalona, A. J. C.; García, D. J. y De la Vega, A. J. 2016. Influencia del medio de extracción en la composición de sustancias volátiles de las hojas de Zanthoxylum pistacifolium Griseb. Rev. Cubana Química. 28(1):490-506. [ Links ]
Hernández, C. J.; Forter, B.; Moreira, X.; Heil, M. and Benrey, B. 2016. Induced floral and extrafloral nectar production affect ant-pollinator interactions and plant fitness. Biotropica. 48(3):342-348. [ Links ]
Hijaz, F.; Nehela, Y. and Killiny, N. 2016. Possible role of plant volatiles in tolerance against huanglongbing in citrus. Plant Signaling Behavior. 11(3):1-12. [ Links ]
Ignacimuthu, S.; Ward, A. R.; Sharma, H. C. and Paulraj, M. G. 2011. Herbivore induced plant volatiles: their role in plant defense for pest management. Plant Signaling and Behavior. 6(12):1973-1978. [ Links ]
Kallenbach, M.; Oh, Y.; Eilers, E. J.; Veit, D.; Baldwin, I. T. and Schuman, M. C. 2014. A robust, simple, high-throughput technique for time-resolved plant volatile analysis in field experiments. The Plant J. 78(6):1060-1072. [ Links ]
Kroes, A.; Weldegergis, B. T.; Cappai, F.; Dicke, M. and van Loon, J. J. A. 2017. Terpenoid biosynthesis in arabidopsis attacked by caterpillars and aphids: effects of aphid density on the attraction of a caterpillar parasitoid. Oecologia. 185(4):699-712. [ Links ]
Laredo, A. E. I.; Martínez, H. J. L.; Iliná, A.; Guillen, C. L. y Hernández, C. F. D. 2017. Aplicación de ácido jasmónico como inductor de resistencia vegetal frente a patógenos. Rev. Mex. Cienc. Agríc. 8(3):673-683. [ Links ]
Maag, D.; Erb, M.; Koollner, T. G. and Gershenzon, J. 2015. Defensive weapons and defense signals in plants: some metabolites serve both roles. Bioessays. 37(2):167-174. [ Links ]
Marín, L. J. C. y Céspedes, C. L. 2007. Compuestos volatiles de plantas. Origen, emisión, efectos, análisis y aplicaciones al agro. Rev. Fitotec. Mex. 30(4):327-351. [ Links ]
Martínez, A. 2013. Introducción a la ecología química y su uso en el manejo de insectos plaga en sistemas forestales. 17. In: manejo integrado de plagas forestales. Villacide, J. y Corley, J. (Eds.). Instituto Nacional de Tecnología Agropecuaria. Argentina. 1-14 pp. [ Links ]
Materic, D.; Bruhn, D.; Turner, C.; Morgan, G.; Mason, N. and Gauci, V. 2015. Methods in plant foliar volatile organic compounds research. Appl. Plant Sci. 3(12):1-13. [ Links ]
Meiners, T. 2015. Chemical ecology and evolution of plant-insect interactions: a multitrophic perspective. Insect Sci. 8:22-28. [ Links ]
Mescher, M. C. and De Moraes, C. M. 2015. Role of plant sensory perception in plant-animal interactions. J. Exp. Bot. 66(2):425-433. [ Links ]
Michereff, M. F. F.; Laumann, R. A.; Borges, M.; Michereff, F. M.; Diniz, I. R.; Neto, A. L. F. and Moraes, M. C. B. 2011. Volatiles mediating a plant-herbivore-natural enemy interaction in resistant and susceptible soybean cultivars. J. Chem. Ecol. 37(3):273-285. [ Links ]
Mofikoya, A. O.; Kim, T. H.; El-Raheem, A. M. A.; Blande, J. D.; Kivimaenpaa, M. and Holopainen, J. K. 2017. Passive adsorption of volatile monoterpene in pest control: aided by proximity and disrupted by ozone. J. Agric. Food Chem. 65(44):9579-9586. [ Links ]
Moraes, M. C. B.; Pareja, M.; Laumann, R. A. and Borges, M. 2008. The chemical volatiles (semiochemicals) produced by neotropical stink bugs (Hemiptera: Pentatomidae). Neotropical Entomol. 37(5):489-505. [ Links ]
Ochoa, E. X. M.; Cantúa, A. J. A.; Aguilera, M. N. A. y Montoya, C. L. 2011. Guía para producir soya en el sur de Sonora. INIFAP-CIRNO)-CENEB. Ciudad Obregón, Sonora, México. Folleto para productores núm. 41. 32 p. [ Links ]
Panighel, A. and Flamini, R. 2014. Applications of solid-phase microextraction and gas chromatography/mass spectrometry (SPME-GC/MS) in the study of grape and wine volatile compounds. Molecules. 19(12):21291-21309. [ Links ]
Peñaflor, M. F. G. V. and Bento, J. M. S. 2013. Herbivore-induced plant volatiles to enhance biological control in agriculture. Neotropical Entomol. 42(4):331-343. [ Links ]
Pickett, J. A. and Khan, Z. R. 2016. Plant volatile-mediated signaling and its application in agriculture: successes and challenges. New Phytol. 212(4):856-870. [ Links ]
Ponzio, C.; Gols, R.; Pieterse, C. M. J. and Dicke, M. 2013. Ecological and phytohormonal aspects of plant volatile emission in response to single and dual infestations with herbivores and phytopathogens. Functional Ecol. 27(3):587-598. [ Links ]
Quintana, R. E.; Morales, V. A. T.; Molina, T. J.; Adame, A. R. M.; Acosta, G. J. A. and Heil, M. 2015. Plant volatiles cause direct, induced and associational resistance in common bean to the fungal pathogen Colletotrichum lindemuthianum. J. Ecol. 103(1):250-260. [ Links ]
Raffo, A.; Masci, M.; Moneta, E.; Nicoli, S.; del Pulgar, J. S. and Paoletti, F. 2018. Characterization of volatiles and identification of odor-active compounds of rocket leaves. Food Chem. 240(1):1161-1170. [ Links ]
Rodríguez, H. V.; Marques, F. A.; Navarro da Silva, M. A.; Annies, V.; Helena, B.; Sales, M. B. H. L. N. and Fernandes, S. M. F. G. 2014. Noctuidae-induced plant volatiles: current situation and prospects. Química Nova. 37(10):1663-1669. [ Links ]
Scala, A.; Allmann, S.; Mirabella, R.; Haring, M. A. and Schuurink, R. C. 2013. Green Leaf Volatiles: A Plant’s Multifunctional Weapon against Herbivores and Pathogens. Inter. J. Mol. Sci. 14(9):17781-17811. [ Links ]
Schettino, M.; Grasso, D. A.; Weldegergis, B. T.; Castracani, C.; Mori, A.; Dicke, M.; Van Lenteren, J. C. and Van Loon, J. J. A. 2017. Response of a predatory ant to volatiles emitted by aphid and caterpillar-infested cucumber and potato plants. J. Chem. Ecol. 43(10):1007-1022. [ Links ]
Sharma, H. C.; Ward, A. R.; Paulraj, M. G.; Ahmad, T.; Buhro, A. H.; Hussain, B. and Ignacimuthu, S. 2012. Mechanisms of plant defense against insect herbivores. Plant Signaling and Behavior. 7(10):1306-1320. [ Links ]
Shi, X.; Chen, G.; Tian, L.; Peng, Z.; Xie, W.; Wu, Q.; Wang, S.; Zhou, X. and Zhang, Y. 2016. The salicylic Acid-mediated release of plant volatiles affects the host choice of Bemisia tabaci. Inter. J. Mol. Sci. 17(7):1-11. [ Links ]
Souza, S. E. A.; Giovanni, S.; Jorge, N. C.; Hoffmann, C.; dos Santos, I. R. M.; Zini, C. A. and Soares, G. L. G. 2017. Development of a HS-SPME-GC/MS protocol assisted by chemometric tools to study herbivore-induced volatiles in Myrcia splendens. Talanta. 17:9-20. [ Links ]
Stam, J. M.; Kroes, A.; Li, Y.; Gols, R.; van Loon, J. J. A.; Poelman, E. H. and Dicke, M. 2014. Plant Interactions with multiple insect herbivores: from community to genes. Annual Rev. Plant Biol. 65:689-713. [ Links ]
Stenberg, J. A.; Heil, M.; Ahman, I. and Björkman, C. 2015. Optimizing Crops for Biocontrol of Pests and Disease. Trends Plant Sci. 20(11):698-712. [ Links ]
Torres, G. G.; Montes, H. S. y Espinosa, G. F. J. 2009. Patrones de variación y distribución geográfica en fenotipos químicos foliares de Persea americana var. drymifolia. Rev. Fitotec. Mex. 32(1):19-30. [ Links ]
Tosh, C. R. and Brogan, B. 2015. Control of tomato whiteflies using the confusion effect of plant odours. Agronomy Sustainable Development. 35(1):183-193. [ Links ]
Vázquez, C. C. F; Valiente, B. J. I.; Caballero, M. P.; Mújica, P. H.; Rodríguez, R. J. and Welti, C. J. 2015. Kinetic and statistical criteria for the selection of conditions of extraction of volatile compounds of piquin pepper (Capsicum annuum L. var. glabriusculum). Rev. Mex. Ing. Quím. 14(2):231-241. [ Links ]
Vidal, D. M.; Fávaro, C. F.; Guimarães, M. M. and Zarbin, P. H. G.; 2016. Identification and synthesis of the male-produced sex pheromone of the soldier beetle Chauliognathus fallax (Coleoptera: Cantharidae). Jo. Braz. Chem. Soc. 27(8):1506-1511. [ Links ]
Vivaldo, G.; Masi, E.; Taiti, C.; Caldarelli, G. and Mancuso, S. 2017. The network of plants volatile organic compounds. Scientific Reports. 7:1-18. [ Links ]
Zhang, P. J.; Huang, F.; Zhang, J. M.; Wei, J. N. and Lu, Y. B. 2015. The mealybug Phenacoccus solenopsis suppresses plant defense responses by manipulating JA-SA crosstalk. Sci. Reports. 5:1-7. [ Links ]
Züst†, T. and Agrawal, A. A. 2016. Mechanisms and evolution of plant resistance to aphids. Nature Plants. 2(1):1-9. [ Links ]
Received: April 01, 2019; Accepted: May 01, 2019
 Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
