











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introduction
MegalostylusSchoenherr, 1840 (Entiminae, Naupactini) is a genus of broad-nosed weevils distributed in the Mexican states of Durango, Guanajuato, Guerrero, Michoacán, Morelos, Oaxaca, Puebla and Veracruz (Champion, 1911; Muñiz-Vélez et al., 2015; Ordóñez-Reséndiz etal., 2008). It was described by Schoenherr (1840) in his monumental work Genera et species curculionidum, cum synonymia hujus familiae, specie novae aut hactenus minus cognitae based on the type species M. sturmi Boheman, 1840 and has been traditionally characterized by the presence of a short antennal scape comparatively stouter at apex than in other genera (Champion, 1911; Figs. 1-3). The most complete treatment of Megalostylus was conducted by Champion (1911), who recognized 9 species and assigned this genus to “Otiorhynchinae alatae”, group “Cyphina”. He also provided a taxonomic key based on 32 morphological characters of adults and the complete distribution records of the species until then.

Figure 1 Habitus photographs of Megalostylus species, dorsal view. A) Megalostylus albicans, female; B) M. brevipilis, male; C) M. dilaticollis, male; D) M. fusiformis male. Arrows and numbers indicate character codification, and the character state is indicated between parentheses. Scale bars = 2 mm.

Figure 2 Habitus photographs of Megalostylus species, dorsal view. A) Megalostylus macrophthalmus, male; B) M. rhodopus, male; C) M. splendidus, male; D) M. sturmi, male. Arrows and numbers indicate character codification, the character state is indicated between parentheses. Scale bars = 2 mm.

Figure 3 Habitus photographs of Megalostylus species, dorsal view. A) Megalostylus tomentosus, male; B) Megalostylodes hirsutus, male; C) morphospecies 1, male; D) morphospecies 2, male. Arrows and numbers indicate character codification, the character state is indicated between parentheses. Scale bars = 2 mm.
The classification of Megalostylus proposed by Champion (1911) more than a century ago needs a revision of the status of the species, the infraspecific varieties, and a phylogenetic analysis, to test the monophyly, species relationships, and synapomorphies that support the genus.
Materials and methods
We examined 347 adult specimens obtained from the following entomological collections: IBUNAM, Instituto de Biología, Universidad Nacional Autónoma de México, Mexico City, Mexico; INECOL, Instituto de Ecología, A. C., Xalapa, Veracruz, Mexico; USNM, National Museum of Natural History, Washington DC, USA; NHRS, Swedish Museum of Natural History, Stockholm, Sweden; and BMNH, Natural History Museum, London, England. Moreover, we included material collected by the working team at the Laboratorio de Sistemática y Evolución de Insectos (LabSei) of the ESSJicarero, UAEM.
Taxon sampling. The outgroup comprises 3 species: Megalostylodes hirsutusChampion, 1911, Naupactus cervinus (Boheman, 1840), and Pantomorus albosignatus Boheman, 1840, the former representing the most closely related genus. The ingroup includes 9 species (Figs. 1-3): M. sturmi Boheman,1840, M. rhodopus Boheman, 1840, M. albicans (Lacordaire, 1876), M. splendidus Chevrolat, 1878, M. brevipilisChampion, 1911, M. dilaticollisChampion, 1911, M. fusiformisChampion, 1911, M. macrophthalmus Champion, 1911, and M. tomentosusChampion, 1911 (Table 1). Additionally, 4 described varieties and 5 specimens of doubtful identification at species-level were included as morphospecies 1-5.
Table 1 List of species included in the cladistic analysis of the genus Megalostylus and their geographic distributions (countries and states).
| Species names | Geographic distributions |
|---|---|
| Pantomorus albosignatus Boheman | Mexico (Aguascalientes, Chihuahua, Coahuila, Mexico City, Durango, Guanajuato, Guerrero, Hidalgo, Monterrey, Oaxaca, Puebla Querétaro, San Luis Potosí, Veracruz, Zacatecas) |
| Naupactus cervinus (Boheman) | Argentina (Buenos Aires, Catamarca, Córdoba, Corrientes, Entre Ríos, Jujuy, Mendoza, Misiones, Salta, Santa Fe, Tucumán), Brasil (Paraná, Río Grande do Sul, Santa Catarina, São Paulo), México (Distrito federal), Uruguay (Artigas, Canelones, Colina, Durazno, Maldonado, Montevideo, Rio Negro, Treinta y Tres) |
| Megalostylodes hirsutus Champion | Mexico (Oaxaca) |
| Megalostylus albicans (Lacordaire) | Mexico (Colima, Estado de México, Guanajuato, Guerrero, Jalisco, Michoacán, Morelos, Nayarit, Oaxaca, Puebla) |
| Megalostylus brevipilis Champion | Mexico (Colima, Guerrero, Oaxaca) |
| Megalostylus dilaticollis Champion | Mexico (Guerrero, Michoacán, Morelos, Jalisco) |
| Megalostylus fusiformis Champion | Mexico (Morelos, Guerrero) |
| Megalostylus macrophthalmus Champion | Mexico (Oaxaca) |
| Megalostylus rhodopus Boheman | Mexico (Oaxaca) |
| Megalostylus splendidus Chevrolat | Mexico (Durango, Guerrero, Morelos, Puebla) |
| Megalostylus sturmi Boheman | Mexico (Guerrero, Michoacán) |
| Megalostylus tomentosus Champion | Mexico (Oaxaca) |
A list of 37 discrete characters (29 binary and 8 multistate) was recorded from adults, including 33 characters from the external morphology, 2 of the female genitalia, and 2 of the male genitalia (Table 2). The selection of characters was based on previous analyses of the tribe Naupactini (del Río, 2009; Lanteri & del Río, 2017; Rosas et al., 2011).
Table 2 List of 37 morphological characters, character states and codes.
| Character | Character states |
|---|---|
| 0. Shape, size, density and color of epistome scales compared to those on the rest of the rostrum | Similar in size, density and color (0); smaller, scarcer and generally different in color (1) |
| 1. Dorsal surface of rostrum | Flat (0); slightly to strongly depressed (1) |
| 2. Pair of dorsolateral carinae | Absent (0); present (1) |
| 3. Sexual dimorphism in antennae | Scape width subequal in males and females (0); scape very wide in males and thin in females (1) |
| 4. Width of scape at apex with respect to width of club, in males | Less than club width (0); greater than club width (1) |
| 5. Shape of scape | Capitate (0); strongly clavate (1) |
| 6. Vestiture of scape | Absent to almost absent (0); scales dispersed leaving integument exposed (1); dense, space between scales reduced or absent, not leaving integument exposed (2) |
| 7. Length of funicular antennomeres 1 and 2 | Antennomere 2 slightly shorter than 1 or both sub-equal (0); antennomere 2 longer than 1 (1) |
| 8. Antennae, scales on funicular antennomere 2 | Absent (0); present (1) |
| 9. Color of verticillate setae of funicle | Whitish or golden (0); brown to black (1) |
| 10. Convexity of eyes | Convex, pronounced curvature, hemispherical shape (0); slightly convex, less pronounced curvature, small elevations (1) |
| 11. Shape of pronotum | Subcylindrical, curved sides and maximum width about half (0); subconical, with sides moderately to strongly curved and strongly divergent from apex to base (1); subconical, with sides slightly curved to straight and strongly divergent from apex to base (2); bell-shaped (3); subtrapezoidal, with sides slightly concave at anterior half and progressively expanding towards lateral sides at posterior half (4) |
| 12. Glabrous areas at midline of pronotum | Absent (0); present (1) |
| 13.Width of pronotum with respect to elytral base | Narrower (0); subequal (1); wider (2) |
| 14. Lateral projection of pronotum | Absent or indistinct (0); slightly projected (1); strongly projected (2) |
| 15. Shape of lateral projections of posterior margin of pronotum | Extended towards sides (0); strongly extended backwards, towards elytral base (1) |
| 16. Sides at basal third of pronotum | Not depressed (0); strongly depressed (1) |
| 17. Shape of posterior margin of pronotum | Straight to slightly bisinuate (0); strongly bisinuate (1) |
| 18. Bulging surface beneath intercoxal granule, at anterior margin of prothorax | Indistinct or absent (0); present (1) |
| 19. Color of legs integument | Light tones, reddish or orange (0); dark tones, black or brown (1) |
| 20. Row of denticles on inner margin of protibia | Absent (0); present (1) |
| 21. Prominence opposite to mucro in protibia | Absent (0); present (1) |
| 22. Elytral anterior margin | Straight (0); bisinuate (1) |
| 23. Shape of elytra in lateral view | Moderately convex (0); almost flat (1) |
| 24. Width of base of scutellum with respect to width of interstria 2 | Reduced to indistinct (0); subequal to width of interstria 2 (1); wider than interstria 2 (2) |
| 25. Scutellum vestiture | Scales similar in color to those covering whole surface of elytra (0); Scales similar, consisted of white scales or seta-like scales (1) |
| 26. Disposition of elytral setae | Recumbent to suberect (0); erect (1) |
| 27. Size of elytral setae | Short (0); long, almost as long as width of interstria 2 at middle (1); very long, longer than width of elytral interestriae 2 at middle (2) |
| 28. Arrangement of elytral setae | Uniform throughout elytral disc (0); only on lateral edges of elytral disc, dorsally without setae (1) |
| 29. Convexity of interstriae | Flat (0); slightly convex (1) |
| 30. Vestiture on legs | Absent or almost absent (0); inconspicuous, scales are scattered leaving integument exposed (1); conspicuous, space between scales is reduced or absent, and covered the integument (2) |
| 31. Deep excavation on metafemur, near apex | Absent (0); present (1) |
| 32. Medial longitudinal depression of ventrites1 and 2, on males | Indistinct or absent (0); present (1) |
| 33. Spermatheca, body shape | Subcylindrical, slender (almost as wide as base of cornu) (0); subspherical (1) |
| 34. Shape of plate of sternite VIII | Subrhomboidal very elongated, with basal part much longer than apical part (0); subrhomboidal slightly elongated, with basal part as long as apical part (1) |
| 35. Aedeagus, sculpture of median lobe | Granulose (0); smooth (1) |
| 36. Aedeagus, shape of apex of median lobe | Acute to slightly acute (0); truncated (1); arrow shape (2) |
For the preparation of genital structures, we followed the methodology described in Rosas et al. (2011) and Lanteri and del Río (2017). A Carl Zeiss Stemi 2000 stereomicroscope, equipped with a reticle eyepiece was used for observations and measurements of the external and internal morphology. Photographs were taken with a Nikon D750 camera equipped with a SIGMA 150 mm 1:2.8 (macro) lens, and drawings were made with Corel Draw (2020 version 22.0.0412). Most characters were illustrated by photographs and drawings, to facilitate the recognition of character states (Figs. 4, 5). They were highlighted with arrows, with indication of character numbers and character states between parentheses.
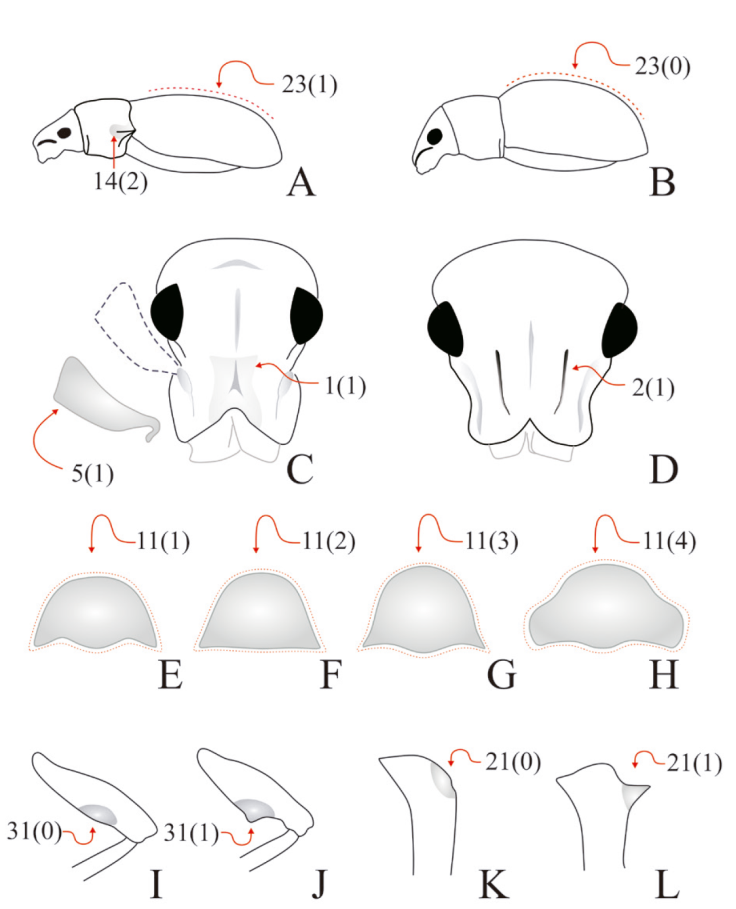
Figure 4 Diagrammatic illustrations with arrows and numbers indicating character codification of head, pronotum, legs, and elytra in Megalostylus and outgroup species. A) Megalostylus dilaticollis, lateral view; B) Pantomorus albosignatus, lateral view; C) M. rhodopus, head; D) Naupactus cervinus, head; E) M. sturmi, pronotum; F) M. tomentosus, pronotum; G) M. brevipilis, pronotum; H) M. dilaticollis, pronotum; I) M. albicans, metafemora; J) Megalostylodes hirsutus, metafemora; K) M. hirsutus, protibiae; L) M. albicans, protibiae.
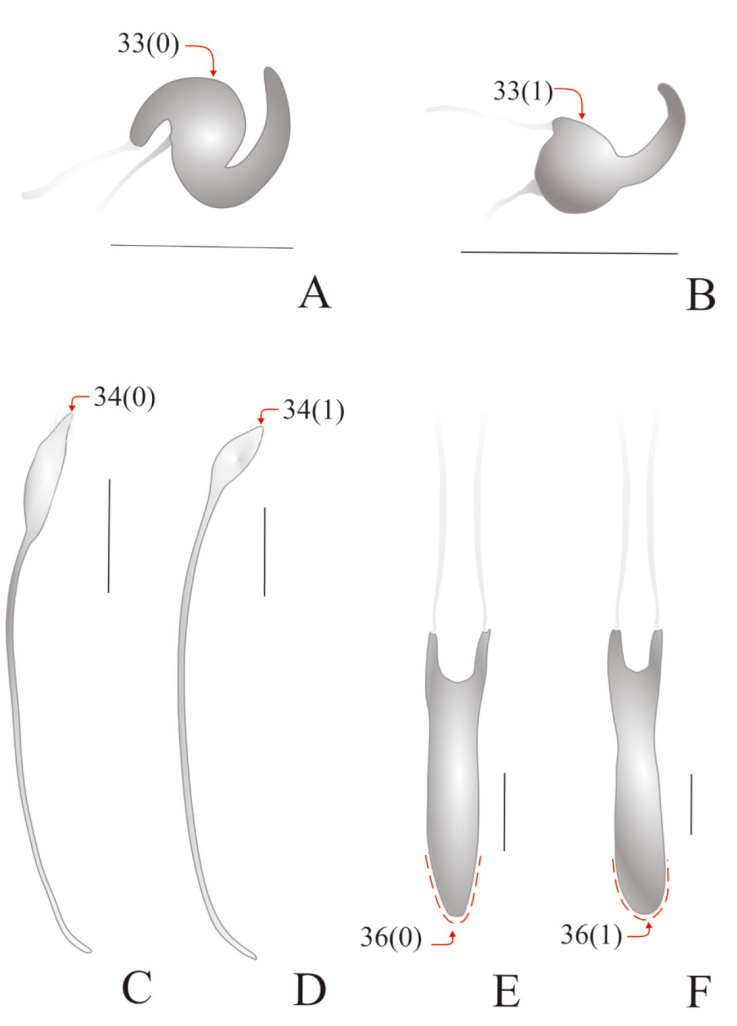
Figure 5 Diagrammatic illustrations with arrows and numbers indicating character codification of female and male genitalia in Megalostylus and outgroup species. A) Pantomorus albosignatus, spermatheca; B) M. albicans, spermatheca; C) M. splendidus, sternite VIII; D) Megalostylodes hirsutus, sternite VIII; E) M. rhodopus, aedeagus; F) M. sturmi, aedeagus. Scale bars = 0.5 mm.
A data matrix of 21 terminal groups and 37 morphological characters was compiled (Table 3). Character states that could not be examined (due to insufficient material) were scored with a “?” and character states with inapplicable entries on various terminals were scored with a “-”. Species or varieties showing 2 or more characters were coded as polymorphic characters. To facilitate the coding, Mesquite software version 3.70 was used (Maddison & Maddison, 2021). An implicit enumeration search was performed in the program TNT version 1.5 (Goloboff & Catalano, 2016). The characters were treated as unordered or non-additive, and under equal weights. The species Pantomorus albosignatus was used to root the trees. To evaluate branch support (BS), a standard Bootstrap with 1000 replicates was calculated in TNT. Branch support values greater than 50 were mapped in the cladogram. The resulting cladogram and character state transformations were examined in WINCLADA under fast optimization. All records were georeferenced, and maps created in ArcMap version 10. 4.1 (ESRI, 2015).
Table 3 Data matrix of 21 taxa and 37 morphological characters of Megalostylus and outgroups used for the cladistic analysis. Inapplicable characters are indicated with a dash (-), and missing characters with a question mark (?).
| Taxa | Characters | ||||||
|---|---|---|---|---|---|---|---|
| 0-4 | 5-9 | 10-14 | 15-19 | 20-24 | 25-29 | 30-36 | |
| Pantomorus albosignatus | 101?0 | 00000 | 00010 | 00000 | 10000 | -0100 | 00?01?? |
| Naupactus cervinus | 10100 | 00100 | 00010 | 00000 | 10100 | 00000 | 00001?2 |
| Megalostylodes hirsutus | 01011 | 10001 | 01101 | 00000 | 00102 | 11201 | 0111100 |
| Megalostylus albicans | 01011 | 12010 | 02012 | 00101 | 01111 | 00000 | 1011010 |
| Megalostylus albicans var. expansus | 01011 | 12010 | 02022 | 00101 | 01111 | 00000 | 101??10 |
| Megalostylus albicans var. farinosus | 01011 | 120?0 | 02012 | 00101 | 01111 | 00000 | 101???? |
| Megalostylus brevipilis | 010?1 | 120?0 | 03011 | 10101 | 01111 | 00000 | 101??10 |
| Megalostylus dilaticollis | 01011 | 12000 | 04022 | 01001 | 01111 | 00000 | 10110?? |
| Megalostylus fusiformis | 010?1 | 120?0 | 03011 | 10101 | 01111 | 00000 | 10????? |
| Megalostylus macrophthalmus | 01011 | 100? | 12011 | 00000 | 01112 | 10000 | 00?10?? |
| Megalostylus splendidus | 01011 | 12010 | 01011 | 10101 | 01112 | 00000 | 1011010 |
| Megalostylus splendidus var. | 01011 | 12010 | 01011 | 10101 | 01112 | 00000 | 1011010 |
| Megalostylus sturmi | 01011 | 12010 | 0100&11 | 10111 | 01112 | 01100 | 1011?11 |
| Megalostylus sturmi var. villosus | 01011 | 12010 | 0100&11 | 10111 | 01112 | 01100 | 1011011 |
| Megalostylus rhodopus | 01011 | 12001 | 01111 | 00000 | 01112 | 10010 | 0011010 |
| Megalostylus tomentosus | 01011 | 110?0 | 02011 | 00000 | 01112 | 01100 | 00110?? |
| Morphospecies 1 | 010?1 | 12001 | 01111 | 10000 | 0111? | 10010 | 001??10 |
| Morphospecies 2 | 010?1 | 11000 | 01011 | 10100 | 01112 | 10000 | 001??10 |
| Morphospecies 3 | 010?1 | 10001 | 02011 | 00000 | 01112 | 10011 | 001??10 |
| Morphospecies 4 | 010?? | 11000 | 02011 | 00000 | 01112 | 01100 | 00?10?? |
| Morphospecies 5 | 010?1 | 12000 | 04011 | 00001 | 01111 | 00000 | 101??10 |
Results
The search for the most parsimonious trees under equal weights yielded one most parsimonious tree (Fig. 6) (L = 63, IC = 0.73, IR = 0.83), showing the following phylogenetic sequence: (Pantomorus albosignatus (Naupactus cervinus (Megalostylodes hirsutus (Megalostylus)))).
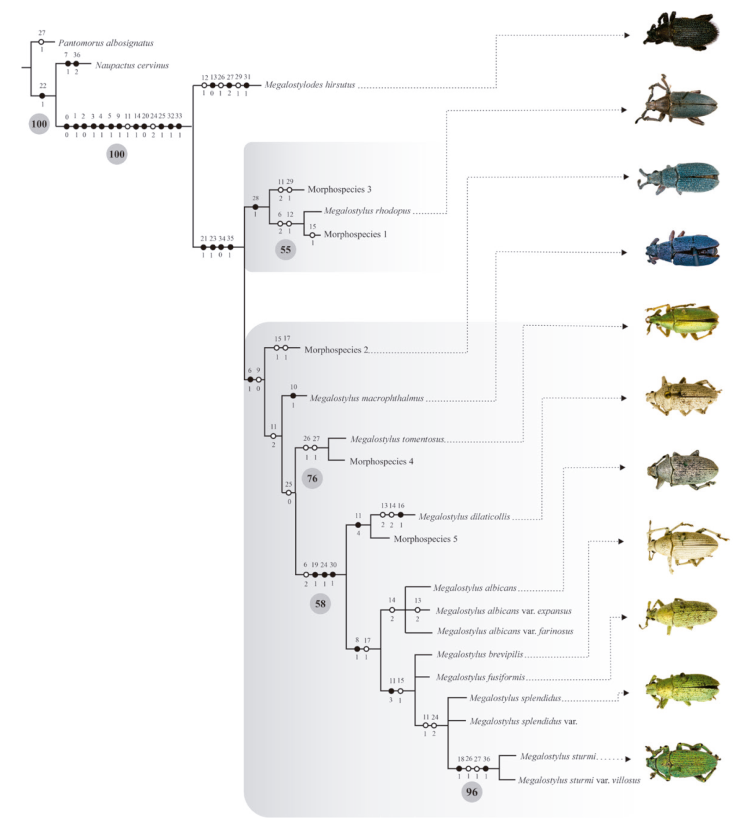
Figure 6 Most parsimonious cladogram of Megalostylus plus 3 outgroups under equal weights, showing character fast optimization. Black circles indicate synapomorphies, whereas white circles indicate non-exclusive synapomorphies. Numbers on and below each circle correspond to character and state codes, respectively. Bootstrap values are indicated below the branches.
Megalostylodes was found to be sister to the genus Megalostylus based on 12 synapomorphies: epistome scales similar in size, density, and color to those on the rest of the rostrum (0:0), dorsal surface of rostrum slightly to strongly depressed (1:1), absence of pair of dorsolateral carinae (2:0), scape very wide in males and thin in females (3:1), width of scape at apex greater than club width in males (4:1), shape of scape strongly clavate (5:1), color brown to black of the verticillate setae of funicle (9:1), slight lateral projection of pronotum (14:1), absence of a row of denticles on inner margin of protibia (20:0), vestiture of scutellum present, consisting of white scales or seta-like scales (25:1), presence of medial longitudinal depression of ventrites 1 and 2 on males (32:1), and subspherical body shape of spermatheca (33:1). There are also 2 homoplastic characters that support this relationship: pronotum subconical, with sides curved and strongly divergent from apex to base (11:1) and width of base of scutellum wider than interestria 2 (24:2).
Megalostylus was recovered as monophyletic based on 4 synapomorphies: presence of prominence opposite to mucro in protibia (21:1), elytra in lateral view almost flat (23:1), plate of sternite VIII, subrhomboidal very elongated (34:0), and sculpture of aedeagus smooth (35:1).
The sister genus Megalostylodes is characterized by 3 autapomorphies: pronotum narrower with respect to elytral base (13:0), very long elytral setae, longer than width of elytral interstriae 2 at middle (27:2), and presence of deeply excavated metafemur near apex (31:1), and 3 non-exclusive synapomorphies: presence of glabrous areas in midline of pronotum (12:1), erect disposition of elytral setae (26:1), and slightly convex interestriae (29:1).
Two main clades were found within Megalostylus: clade I, including Megalostylus rhodopus and morphospecies 1 and 3 (considered belonging to M. rhodopus), is supported by 1 exclusive synapomorphy: elytral setae only present on the lateral edges of elytral disc (28:1). Clade II is supported by 1 exclusive synapomorphy and 1 non-exclusive synapomorphy: scales of scape separate, leaving integument exposed (6:1) and whitish or golden color of the verticillate setae of funicle (9:0), respectively. Within clade II, morphospecies 2 is sister to the remaining species, which form a clade supported by 1 non-exclusive synapomorphy: pronotum subconical, sides slightly curved to straight and strongly divergent from apex to base (11:2). Megalostylus macrophthalmus is the sister species of M. tomentosus-M. sturmi var. villosus based on 1 non-exclusive synapomorphy: vestiture of scutellum present, scales similar in color to those covering the whole surface of elytra (25:0). Megalostylus tomentosus (along with morphospecies 4) is the sister of a supported clade (BT 58) M. dilaticollis- M. sturmi var. villosus clade, which is supported by 3 exclusive synapomorphies and 1 non-exclusive synapomorphy: integument of legs with dark tones, black or brown (19:1), and width of base of scutellum reduced to indistinct (24:1), vestiture on legs inconspicuous, scales are scattered leaving integument exposed (30:1), and scales separated leaving integument of scape exposed (6:2). Within this clade M. dilaticollis (along with morphospecies 5) is sister to a subclade supported by 1 exclusive synapomorphy and 1 non-exclusive synapomorphy: presence of scales on the second funicular antennomere (8:1) and posterior margin of pronotum strongly bisinuate (17:1) that includes M. albicans and its varieties, M. albicans var. expansus, and var. farinosus sister to M. brevipilis to M. sturmi var. villosus clade supported by 1 exclusive synapomorphy and 1 non-exclusive synapomorphy: pronotum flared or bell- shaped (11:3) and lateral projections of posterior margin of pronotum strongly extended backward, towards elytral base (15:1). Within this clade, M. brevipilis and M. fusiformis are sisters to the M. splendidus-M. sturmi var. villosus clade which is supported by 2 non-exclusive synapomorphies: pronotum subconical, sides curved and strongly divergent from apex to base (11:1), and width of base of scutellum wider than interestria 2 (24:2). The clade M. splendidus-M. sturmi var. villosus also has unresolved relationships between M. splendidus and his variety, both sister to the M. sturmi + M. sturmi var. villosus clade.
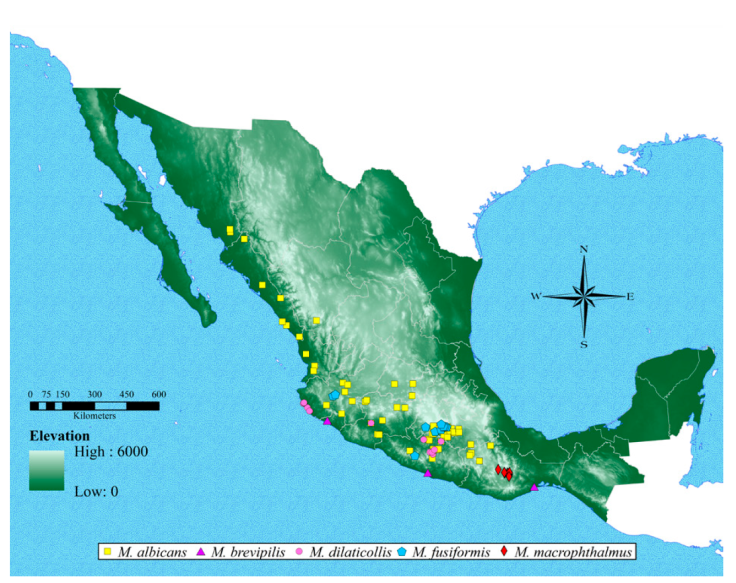
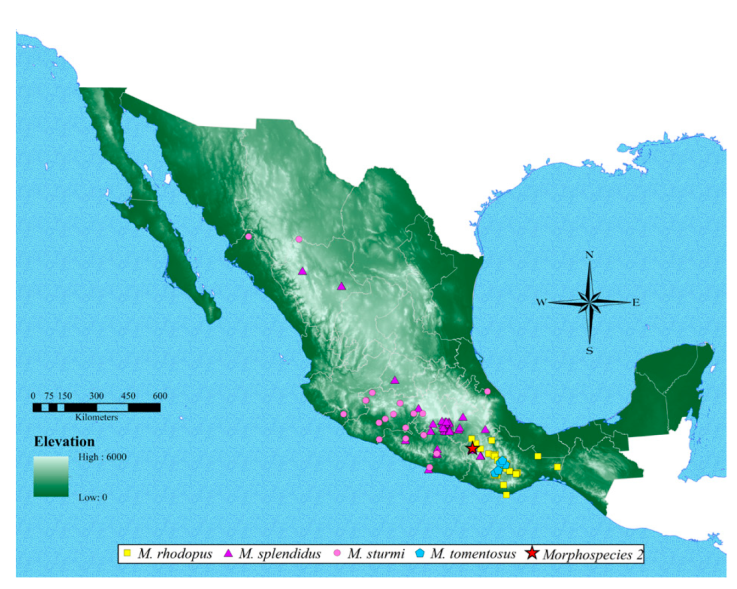
Geographic distribution. The maps (Figs. 7, 8) indicate that the species of Megalostylus are mostly distributed along the Pacific coast, with the greatest richness in the central and southern parts of the country. Megalostylus albicans shows the largest distribution, ranging from Oaxaca to Sinaloa. Six species are distributed in the central states of the country (Guanajuato, Guerrero, Michoacán, Morelos, and Puebla): M. albicans, M. brevipilis, M. dilaticollis, M. fusiformis, M. splendidus, and M. sturmi. The species M. rhodopus, M. macrophthalmus, and M. tomentosus are restricted to Oaxaca, and M. rhodopus reaches the southernmost distribution extending to the Isthmus of Tehuantepec.
Map by A. Lizama-Hernández.
Figure 7 Occurrence map of 5 Megalostylus species, ordered alphabetically. Megalostylus albicans (yellow square); M. brevipilis (purple triangle); M. dilaticollis (pink circle); M. fusiformis (blue pentagon) and M. macrophthalmus (red diamond).
Map by A. Lizama-Hernández.
Figure 8 Occurrence map of remaining 4 Megalostylus species, ordered alphabetically plus a new Megalostylus species (indicated here as morphospecies 2). Megalostylus rhodopus (yellow square); M. splendidus (purple triangle); M. sturmi (pink circle); M. tomentosus (blue pentagon) and morphospecies 2 sp. nov. (red star).
Discussion
The sister group relationship between Megalostylodes and Megalostylus is well supported by 12 exclusive synapomorphies, and the monophyly of Megalostylus is supported by a combination of 4 exclusive synapomorphies of the protibiae, elytra, and female and male genitalia. Champion (1911) distinguished Megalostylus based on the presence of a prominence opposite to mucro (or toothed tibiae at the external apical angle), and Lanteri and del Río (2017) recovered this character state (car 69[1]) as an apomorphy of Megalostylus.
Within Megalostylus most of the relationships are not well supported nevertheless, it is clear the position of M. rhodopus (including morphospecies 1 and 3) as sister species of the remaining, and it was recovered a well-supported group including M. dilaticollis (= morphospecies 5), M. albicans, M. brevipilis, M. fusiformis, M. splendidus, and M. sturmi. Megalostylus macrophthalmus and M. tomentosus (= morphospecies 4) would be basal regarding this clade. Morphospecies 2 is considered a probable new species, however, this status needs further studies.
The species of Megalostylus are very variable in scale color, size and pronotum shape, and due to that intraspecific variation, some nominal species have been synonymized: M. farinosus Chevrolat, 1878 [junior synonym of M. albicans (Lacordaire, 1876)], M. expansus Pascoe, 1881 [junior synonym of M. albicans (Lacordaire, 1876)] and M. villosus Chevrolat, 1878 [junior synonym of M. sturmi Boheman, 1840]. Our analysis explored the relationships among intraspecific varieties of the genus Megalostylus, and the results support the previously established synonymies. In the case of M. albicans with its varieties (M. albicans var. expansus and M. albicans var. farinosus) there is no reason to consider them as different species. Characters with the greatest contribution to the species relationships are those referring mainly to the vestiture of antennae, legs, elytra, shape, and characteristics of the pronotum, scutellum, and apex of aedeagus.

