











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
http://zoobank.org/urn:lsid:zoobank.org:pub:E20553D6-CDA6-423F-869A-56DFF6008D0A
Introduction
Tardigrades (tardus = slow, gradus = step or “slow-stepper”) have commonly been called “water bears” due to their bear-like appearance, legs with claws, and slow lumbering gait (Nelson et al., 2015, 2018). These micrometazoans are found in a wide variety of marine, freshwater and terrestrial habitats, such as tropical forests and polar and arid deserts, from mountains to the depths of the oceans, and inhabit mainly mosses and lichens (Nelson & Marley, 2000; Nelson et al., 2015; Ramazzotti & Maucci, 1983). Being hydrophilic organisms, they need an aqueous medium to move and carry out their life cycle. Particularly terrestrial tardigrades depend on water films that adhere to the substrates where they live (Glime, 2017; Nelson et al., 2015).
To date, more than 1,400 species of tardigrades have been described in 3 classes (Eutardigrada, Heterotardigrada, and Mesotardigrada) (Bertolani et al., 2014; Degma & Guidetti, 2024; Guidetti & Bertolani, 2005). As for Mexico, 83 species have been recorded in 16 of the 32 states: Baja California, Chiapas, Chihuahua, Coahuila, Mexico City, Michoacán, Morelos, Nuevo León, Oaxaca, Quintana Roo, San Luis Potosí, Sinaloa, Sonora, Estado de México, Tamaulipas and Yucatán (Anguas-Escalante et al., 2020; Beasley, 1972; Beasley et al., 2008; Dueñas-Cedillo et al., 2020, 2024; García-Román et al., 2022; Heinis, 1911; Kaczmarek et al., 2011; León-Espinosa et al., 2017, 2019; Moreno-Talamantes & León-Espinosa, 2019; Moreno-Talamantes et al., 2015, 2019, 2020; May, 1948; Núñez et al., 2021; Pérez-Pech et al., 2017, 2018, 2020; Pilato, 2006; Pilato & Lisi, 2006; Ramazzotti & Maucci, 1983; Schuster, 1971). The presence of tardigrades in the rest of the Mexican territory is unknown. In this article, we present new records of tardigrades found in samples of mosses and lichens collected in the Cantera Oriente, buffer zone that belongs to the Pedregal de San Angel Ecological Reserve (REPSA), southern Mexico City. We describe a new species, Paramacrobiotus puma sp. nov., by using an integrative approach, including morphological analysis with phase-contrast microscopy (PCM), scanning electron microscopy (SEM) as well as DNA sequencing.
Materials and methods
Five samples of mosses and 2 samples of lichens were collected within the Cantera Oriente (19°19’00.5” N, 99°10’21.5” W; 2,260 m asl) on October 2, 2015, and August 30, 2016, respectively. The area is composed mainly of high elevation xerophilic scrub and is located in southern Mexico City and corresponds to the Buffer Zone (A3) of the Pedregal de San Ángel Ecological Reserve (REPSA) (Fig. 1), produced by the Xitle Volcano’s eruption approximately 1,670 years ago. This ecological reserve is a conservation area created in 1983 within the campus of the Universidad Nacional Autónoma de México (UNAM). The objective of the reserve is to maintain an area of biological and cultural diversity containing the last remnants of natural ecosystems in the southern Mexico Basin (Palacio & Guilbaud, 2015). The samples covered an area of approximately 5 cm 2 and were deposited in brown paper bags and taken to the laboratory for examination. The specimens were collected under the Scientific Collector Permits FAUT-0027 and FAUT-209 granted by Semarnat and with project 361 issued by the Executive Secretariat of REPSA. Tardigrade specimens were deposited in the Colección de Tardígrados associated with the Colección Nacional de Ácaros (CNAC), Instituto de Biología, UNAM, Mexico City.
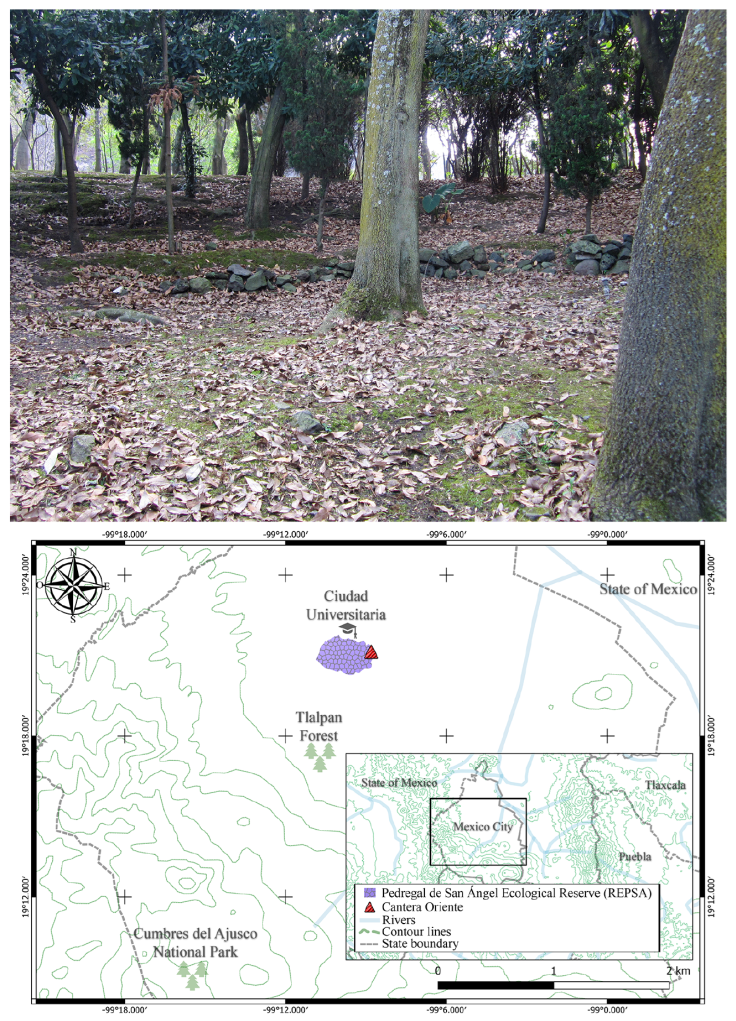
Map by D. López-Sandoval.
Figure 1 View of collection site within the buffer zone (A3), and map indicating location of the Cantera Oriente and Pedregal de San Ángel Ecological Reserve (REPSA), CDMX, Mexico.
Tardigrades and eggs were extracted from the samples using the technique described by Dastych (1985). Specimens for light microscopy were mounted on slides with Hoyer’s liquid and were observed under the microscope Nikon Optiphot-2 using phase contrast microscopy (PCM). Some specimens were separated for Scanning Electron Microscopy (SEM) and were prepared according to the technique described by Stec et al. (2015). Images were obtained in a SEM Hitachi, model SU 1510 at the Laboratorio Nacional de la Biodiversidad (LANABIO), Instituto de Biología, Universidad Nacional Autónoma de México.
The sample size for morphometrics was chosen following recommendations by Stec et al. (2016). All measurements are in micrometers (µm). For the measurements, terminology of the structures and claws of the buccopharyngeal apparatus follow Pilato and Binda (2010) and Michalczyk and Kaczmarek (2003). The claw measurements were according to Beasley et al. (2008) and Kaczmarek and Michalczyk (2017). The macroplacoid length sequence was determined according to Kaczmarek, Cytan et al. (2014) and Kaczmarek and Michalczyk (2017). The terminology and measurements for Apochela follow Tumanov (2006), Michalczyk et al. (2012a, b), and Pilato et al. (2016). The pt ratio is the relation between the length of a given structure and the length of the buccal tube, expressed as a percentage (Pilato, 1981). Morphometric data was managed using templates “Parachela” and “Apochela”. Versions 1.2 for both templates are available in the Tardigrada Register (Michalczyk & Kaczmarek, 2013). Tardigrade taxonomy follows Bertolani et al. (2014) and Stec et al. (2021). The morphometric data is given in supplementary material (SM1).
The specimens examined were identified with taxonomic keys and compared with original species descriptions and other useful literature: Tumanov (2006, 2020), Pilato and Lisi (2006), Claxton (1998), Guidetti et al. (2009, 2022), Pilato and Binda (2010), Michalczyk et al. (2012a, b), Pilato et al. (2016), Kaczmarek et al. (2017), Gąsiorek, Stec, Morek et al. (2018), Stec, Morek et al. (2018), Moreno-Talamantes et al. (2019, 2020), Dueñas-Cedillo et al. (2020), Morek and Michalczyk (2020), Stec, Morek et al. (2018), Stec, Roszkowska et al. (2018), Stec, Kristensen et al. (2020), Stec, Krzywański et al. (2020), Rocha et al. (2022), Kayastha, Mioduchowska et al. (2023), Kayastha, Stec et al. (2023). For comparison with the new species, we also examined the following type material deposited at the Institute of Zoology and Biomedical Research, Jagiellonian University, Gronostajowa, Kraków, Poland: Paramacrobiotus areolatus (Murray, 1907) (slides NO.385.63 - NO.385.71, NO.385.75 - NO.385.78 and NO.385.81) and P. lachowskaeStec, Roszkowska, Kaczmarek and Michalczyk, 2018 (slides CO.018.04 - CO.018.21).
Before DNA extraction, specimens were mounted in water on a glass slide and examined under a Nikon Optiphot-2 optical microscope to confirm identification. The DNA was extracted from individual animals following a modified protocol by Casquet et al. (2012), using the Chelex® 100 resin (Bio-Rad) extraction method. Each specimen was placed individually in a 1.5 ml Eppendorf microcentrifuge tube, in 50 μl of a 5% suspension of 75-150 μm wet bead size Chelex® 100 resin (Bio-Rad) in ddH2O with the addition of 3.5 μl Proteinase K (A&A Biotechnology) and incubated at 56 ºC for 1 h. Then, tubes were incubated at 95 ºC for 30 min and centrifuged at 4,500 rpm for 15 min. After that, the supernatant was transferred to new 1.5 ml tubes and stored at -20 ºC. After the extraction, the hologenophores (Pleijel et al., 2008), were mounted in Hoyer’s medium. Four DNA fragments were sequenced: the small ribosome subunit (18S rRNA, nDNA), the large ribosome subunit (28S rRNA, nDNA), the internal transcribed spacer 2 (ITS -2, nDNA), and the cytochrome oxidase subunit I (COI, mtDNA). Primers and original references for specific PCR programs for amplification of the fragments are listed in Table 1.
Table 1 PCR primers for amplification of the 4 DNA fragments sequenced in the present study.
| DNA marker | Primer name | Primer direction | Primer sequence (5’-3’) | Source | PCR programme source |
|---|---|---|---|---|---|
| 18S rRNA | SSU01_F | forward | AACCTGGTTGATCCTGCCAGT | Sands et al. (2008) | Zeller (2010) |
| SSU82_R | reverse | TGATCCTTCTGCAGGTTCACCTAC | |||
| 28S rRNA | 28S_Eutar_F | forward | ACCCGCTGAACTTAAGCATAT | Gąsiorek, Stec, Zawierucha et al. (2018) | Stec, Kristensen |
| et al. (2020) | |||||
| 28SR0990 | reverse | CCTTGGTCCGTGTTTCAAGAC | Mironov et al. (2012) | ||
| ITS-2 | ITS2_Eutar_Ff | forward | CGTAACGTGAATTGCAGGAC | Stec, Morek et al. (2018) | Stec, Kristensen et al. (2020) |
| ITS2_Eutar_Rr | reverse | TCCTCCGCTTATTGATATGC | |||
| COI | COI_Para_F | forward | GGTCAACAAATCATAAAGATATTGG | Gąsiorek et al. (2017) | Michalczyk et al. (2012a) |
| COI_Mac_Rr | reverse | TAAACTTCAGGGTGACCAAAAAATCA | Stec, Krzywański et al. (2020) |
For every PCR reaction, the solution contained 9.5 μl ddH2O, 3 μl 5x MyTaq Reaction Buffer (Bioline™), 0.2 μl 10 mM forward primer, 0.2 ml 10 mM reverse primer, 0.1 μl MyTaq™ DNA Polymerase, (Bioline™) (5U/μl), and 2 μl of genomic DNA extract. The PCR products were controlled by 1.5% agarose gel electrophoresis stained with GelRed Nucleic Acid Gel Stain, 10,000X (Biotium™) and purified with the ExoSap-IT enzyme (Applied Biosystems), following the manufacturer’s instructions. The sequence reaction was prepared with 4 μl of water, 2 μl of Buffer 5X, 2 μl of big dye Terminator v3.1 (Applied Biosystems), 1 μl of the primer, and 2.5 μl of the purified product. The reaction was placed in a PCR 2720 with the program suggested by the manufacturer. When finished, they were purified with Sephadex CentriSep™ plates (Princeton) and read in a 3730xl sequencer (Applied Biosystems), at the LANABIO.
All sequences were assembled, manually inspected, and processed in SeqTrace (Stucky, 2012) and submitted to GenBank.
The identity of the obtained sequences was verified using the Basic Local Alignment Search Tool (Altschul etal., 1990). As a support for the morphological comparisons between the new species and other Paramacrobiotus species, several sequences deposited in GenBank of the 4 sequenced markers were used to calculate the uncorrected genetic distances (p-distance), applying the program MEGA X (Kumar et al., 2018). The distance matrices are provided in the supplementary material (SM2).
In order to establish the phyletic position of the new species, a phylogenetic tree was constructed using the concatenated 18S rRNA + 28S rRNA + ITS-2 + COI sequences of the genus Paramacrobiotus with the sequences of 2 Minibiotus species as an outgroup (Table 2). Sequences of the newly barcoded species and sequences of species obtained from GenBank were aligned with the MAFFT algorithm version 7 (Katoh et al., 2002) implemented in the MAFFT online service (Katoh et al., 2019). Sequences were checked by visual inspection and translated to amino acids by using the invertebrate mitochondrial code implemented in MEGA X (Kumar et al., 2018) to check for the presence of pseudogenes. The sequences were concatenated using SequenceMatrix (Vaidya et al., 2011) and before partitioning, the concatenated alignment was divided into 6 data blocks constituting 3 separate blocks of ribosomal markers and 3 separate blocks of 3 codon positions in the COI data set. We selected the best scheme of partitioning with the program ModelFinder (Kalyaanamoorthy et al., 2017) and the best substitution model for the posterior phylogenetic analysis using ModelTest-NG (Darriba et al., 2020). The best-fit substitution model for each partition under the Bayesian Information Criterion (BIC) was: GTR+G4 for the first and the second codon positions and GTR+I+G4 for the third codon position in COI data set. As for the ribosomal markers the best-fit model was: HKY+G4 for ITS2 marker data set, HKY+I+G4 for 28S marker data set and HKY+I for 18S marker data set. Bayesian inference (BI) marginal posterior probabilities were calculated using MrBayes version 3.2 (Ronquist & Huelsenbeck, 2003). The analysis was run for 10 million generations using random starting trees and sampling the Markov chain every 1,000 generations. An average standard deviation of split frequencies of < 0.01 was used as a guide to ensure the 2 independent analyses had converged. To ensure Markov chains had reached stationarity and to determine the correct “burn-in” for the analysis (which was the first 10% of generations), the program Tracer version 1.7 (Rambaut et al., 2018) was used. The ESS values were > 200 and the consensus tree was obtained after summarizing the resulting topologies and discarding the “burn-in”. Additionally, a maximum likelihood (ML) analysis was run using Iqtree2 (Minh et al., 2020) and the branch support values of the ML tree were measured using 1,000 ultrafast bootstrap replicates (UFBoot) (Hoang et al., 2018). The final consensus trees were viewed and edited in FigTree version 1.4.4 available from http://tree.bio.ed.ac.uk/software/figtree
Table 2 Accession numbers sequences used for phylogenetic analysis downloaded from GenBank.
| Taxon | 18S rRNA | 28S rRNA | ITS-2 | COI | Source |
|---|---|---|---|---|---|
| Paramacrobiotus aff. richtersi BR.009 1 | MH664934 | MH664952 | MH666082 | MH676000 | Stec, Krzywański et al. (2020) |
| Paramacrobiotus aff. richtersi BR.009 2 | - | - | - | MH676001 | Stec, Krzywański et al. (2020) |
| Paramacrobiotus aff. richtersi BR.009 3 | - | - | - | MH676002 | Stec, Krzywański et al. (2020) |
| Paramacrobiotus aff. richtersi HU.012 1 | MH664936 | MH664954 | MH666084 | MH676005 | Stec, Krzywański et al. (2020) |
| Paramacrobiotus aff. richtersi HU.012 2 | - | - | - | MH676006 | Stec, Krzywański et al. (2020) |
| Paramacrobiotus aff. richtersi MG.002 1 | MH664938 | MH664956 | MH666086 | MH676008 | Stec, Krzywański et al. (2020) |
| Paramacrobiotus aff. richtersi MG.002 2 | - | - | MH666087 | - | Stec, Krzywański et al. (2020) |
| Paramacrobiotus aff. richtersi NO.386 | MH664939 | MH664957 | MH666088 | MH676009 | Stec, Krzywański et al. (2020) |
| Paramacrobiotus aff. richtersi NZ.001 | MH664940 | MH664958 | MH666089 | MH676010 | Stec, Krzywański et al. (2020) |
| Paramacrobiotus aff. richtersi TZ.018 | MH664933 | MH664951 | MH666095 | MH676017 | Stec, Krzywański et al. (2020) |
| Paramacrobiotus arduusGuidetti et al., 2019 | MK041032 | - | - | MK041020 | Guidetti et al. (2019) |
| Paramacrobiotus areolatus (Murray, 1907) | MH664931 | MH664948 | MH666080 | MH675998 | Stec, Krzywański et al. (2020) |
| Paramacrobiotus bengalenseisBasu et al., 2023 | ON923868 | - | - | OP531839 | Basu et al. (2023) |
| Paramacrobiotus bifrons (Pontremoli) 1 Brandoli et al., 2024 | - | - | PP240910 | PP236542 | Brandoli et al. (2024) |
| Paramacrobiotus bifrons (Pontremoli) 2 Brandoli et al., 2024 | - | - | PP240911 | PP236543 | Brandoli et al. (2024) |
| Paramacrobiotus bifrons (Pontremoli) 3 Brandoli et al., 2024 | - | - | PP240912 | PP236544 | Brandoli et al. (2024) |
| Paramacrobiotus bifrons (Pontremoli) 4 Brandoli et al., 2024 | - | - | PP240913 | PP236545 | Brandoli et al. (2024) |
| Paramacrobiotus bifrons (Gombola) 1 Brandoli et al., 2024 | - | - | - | PP236546 | Brandoli et al. (2024) |
| Paramacrobiotus bifrons (Gombola) 2 Brandoli et al., 2024 | - | - | PP240914 | PP236547 | Brandoli et al. (2024) |
| Paramacrobiotus bifrons (Gombola) 3 Brandoli et al., 2024 | - | - | PP240915 | PP236548 | Brandoli et al. (2024) |
| Paramacrobiotus bifrons (Gombola) 4 Brandoli et al., 2024 | - | - | PP240916 | PP236549 | Brandoli et al. (2024) |
| Paramacrobiotus bifrons (Gombola) 5 Brandoli et al., 2024 | - | - | PP240917 | PP236550 | Brandoli et al. (2024) |
| Paramacrobiotus bifrons (Gombola) 6 Brandoli et al., 2024 | - | - | PP240918 | PP236551 | Brandoli et al. (2024) |
| Paramacrobiotus bifrons (Sassi di Varana) | - | - | - | PP236552 | Brandoli et al. (2024) |
| Brandoli et al., 2024 | |||||
| Paramacrobiotus bifrons (Monte Sant’Angelo) 1 | - | - | PP240919 | PP236553 | Brandoli et al. (2024) |
| Brandoli et al., 2024 | |||||
| Paramacrobiotus bifrons (Monte Sant’Angelo) 2 | - | - | PP240920 | PP236554 | Brandoli et al. (2024) |
| Brandoli et al., 2024 | |||||
| Paramacrobiotus bifrons (Monte Sant’Angelo) 3 | - | - | PP240921 | PP236555 | Brandoli et al. (2024) |
| Brandoli et al., 2024 | |||||
| Paramacrobiotus celsusGuidetti et al., 2019 | MK041031 | - | - | MK041019 | Guidetti et al. (2019) |
| Paramacrobiotus cf. klymenki IT.048 | MH664937 | MH664955 | MH666085 | MH676007 | Stec, Dudziak et al. (2020) |
| Paramacrobiotus cf. klymenki PT.006 | MH664943 | MH664960 | MH666092 | MH676013 | Stec, Dudziak et al. (2020) |
| Paramacrobiotus depressusGuidetti et al., 2019 | MK041030 | - | - | MK041015 | Guidetti et al. (2019) |
| Paramacrobiotus experimentalisKaczmarek et al., 2020 | MN073468 | MN073465 | MN073464 | MN097837 | Kaczmarek et al. (2020) |
| Paramacrobiotus fairbanksi | MH664941 | MH664950 | MH666090 | MH676011 | Stec, Krzywański et al. (2020) |
| Paramacrobiotus filipi 1 Dudziak et al., 2020 | MT261913 | MT261904 | - | MT260372 | Stec, Dudziak et al. (2020) |
| Paramacrobiotus filipi 2 Dudziak et al., 2020 | - | - | - | MT260373 | Stec, Dudziak et al. (2020) |
| Paramacrobiotus gadabouti MD50.1 Kayastha et al., 2023 | OP394210 | - | - | OP394113 | Kayastha, Stec et al. (2023) |
| Paramacrobiotus gadabouti MD50.4 Kayastha et al., 2023 | OP394212 | - | - | OP394114 | Kayastha, Stec et al. (2023) |
| Paramacrobiotus gadabouti AU.044 Kayastha et al., 2023 | MH664932 | MH664949 | MH666081 | MH675999 | Stec, Krzywański et al. (2020) |
| Paramacrobiotus gadabouti FR.077 1 Kayastha et al., 2023 | MH664935 | MH664953 | MH666083 | MH676003 | Stec, Krzywański et al. (2020) |
| Paramacrobiotus gadabouti FR.077 2 Kayastha et al., 2023 | - | - | - | MH676004 | Stec, Krzywański et al. (2020) |
| Paramacrobiotus gadabouti PT.048 1 Kayastha et al., 2023 | MH664944 | MH664961 | MH666093 | MH676014 | Stec, Krzywański et al. (2020) |
| Paramacrobiotus gadabouti PT.048 2 Kayastha et al., 2023 | - | - | - | MH676015 | Stec, Krzywański et al. (2020) |
| Paramacrobiotus gadabouti TN.014 Kayastha et al., 2023 | MH664945 | MH664962 | MH666094 | MH676016 | Stec, Krzywański et al. (2020) |
| Paramacrobiotus lachowskaeStec et al., 2018 | MF568532 | MF568533 | MF568535 | MF568534 | Stec, Roszkowska et al. (2018) |
| Paramacrobiotus metropolitanusSugiura et al., 2022 | LC637243 | LC649795 | LC649794 | LC637242 | Sugiura et al. (2022) |
| Paramacrobiotus puma sp. nov. | PP416751 | PP416752 | PP416753 | PP414782 | Present study |
| Paramacrobiotus richtersi (Murray, 1911) | MK041023 | - | - | MK040994 | Guidetti et al. (2019) |
| Paramacrobiotus richtersi S38 (Murray, 1911) | OK663224 | OK663235 | OK663213 | OK662995 | Vecchi et al. (2022) |
| Paramacrobiotus spatialisGuidetti et al., 2019 | MK041024 | - | - | MK040996 | Guidetti et al. (2019) |
| Paramacrobiotus spatialis S107 Guidetti et al., 2019 | OK663225 | OK663236 | OK663214 | OK662996 | Vecchi et al. (2022) |
| Paramacrobiotus tonolli US (Ramazzotti, 1956) | MH664946 | MH664963 | MH666096 | MH676018 | Stec, Krzywański et al. (2020) |
| Minibiotus ioculatorStec et al. 2020 | MT023998 | MT024041 | MT024000 | MT023412 | Stec, Kristensen et al. (2020) |
| Minibiotus pentannulatus Londoño et al., 2017 | MT023999 | MT024042 | MT024001 | MT023413 | Stec, Kristensen et al. (2020) |
Results
We obtained 110 tardigrades and 27 eggs from the class Eutardigrada. The specimens examined belong to 2 orders (Apochela and Parachela), 4 families (Milnesiidae, Hypsibiidae, Macrobiotidae, and Ramazzottiidae), 6 genera (Milnesium, Hypsibius, Notahypsibius, Minibiotus, Paramacrobiotus, and Ramazzottius) and 9 species are recorded. We found 3 species which correspond to new records for Mexico and for Mexico City, and 1 species is new to science.
Class Eutardigrada Richters, 1926
Order Apochela Schuster, Nelson, Grigarick and Christenberry, 1980
Family Milnesiidae Ramazzotti, 1962
Genus Milnesium Doyère, 1840
Milnesium longiungueTumanov, 2006
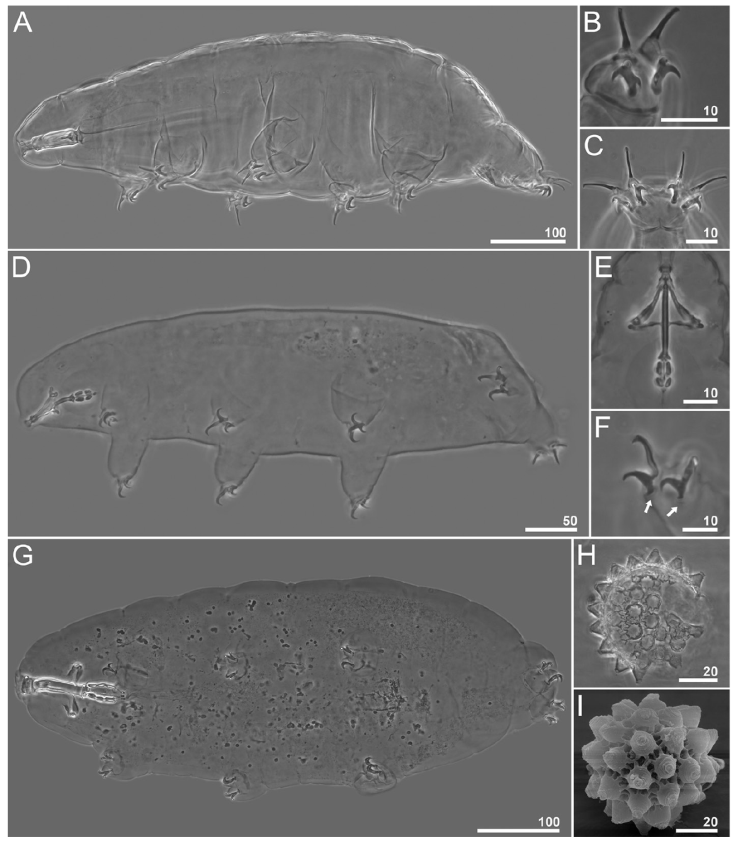
Figure 2 Tardigrade species that represent new records for Mexico. A-C, Milnesium longiungue: A, habitus; B, claws II; C, claws IV. D-F, Notahypsibius pallidoides: D, habitus; E, buccopharyngeal apparatus; F, claws III; white arrows indicate the thickened region on the lunule margin. G-I, Paramacrobiotus gadabouti: G, habitus; H, egg seen on PCM; I, egg seen on SEM. Scale bars = μm.
Taxonomic summary
Type locality: Hymalaia, India.
Material examined: Mexico: Mexico City: Cantera Oriente (19°19’00.5” N, 99°10’21.5” W; 2,260 m asl), 8 specimens (CNAC-Tar000243 - CNAC-Tar000250). Coll. D. López and G. Montiel.
Habitat: high elevation xerophilic scrub. Elevation: 2,260 m asl.
Microhabitat: mosses of the species Thuidium delicatulum (Hedw.) Schimp. collected on a rock.
Remarks. The specimens examined correspond well to the original description by Tumanov (2006). The cuticle is smooth and white colored, the eyes are present, 6 peribuccal lamellae are present, accessory points on primary branches are absent, secondary branches of external claws I-III and of posterior claws IV with 3 points (claw configuration [3-3] - [3-3]), pt of the primary branch IV length is between 82-91% and cuticular bars under the I-III are present. This species has been recorded previously in China (Beasley & Miller, 2007). This is the second record of the species outside the type locality in Hymalaia, India. New record for Mexico.
Milnesium cf. reductum
Taxonomic summary
Material examined: Mexico: Mexico City: Cantera Oriente (19°19’00.5” N, 99°10’21.5” W; 2,260 m asl), 5 specimens (CNAC-Tar000238 - CNAC-Tar000242). Coll. D. López and G. Montiel.
Habitat: high elevation xerophilic scrub. Elevation: 2,260 m asl.
Microhabitat: mosses of the species Thuidium delicatulum collected on a rock.
Remarks. The 5 specimens examined present a smooth and reddish colored cuticle, the eyes are present, 6 peribuccal lamellae are present, secondary branches of external claws I-III and of posterior claws IV with 2 points (claw configuration [2-3] - [3-2]), pt of the primary branch IV length is between 65-69% and the cuticular bars under legs I-III are present. Those characters correspond to the original description of Milnesium reductumTumanov, 2006 (Tumanov, 2006); however, we were unable to confirm the presence or absence of accessory points on primary branches in all specimens. Therefore, due to the lack of a greater number of specimens for examination, for morphometric measurements and for sequencing of genetic material, this species cannot be identified with certainty.
Order Parachela Schuster, Nelson, Grigarick and Christenberry, 1980
Superfamily Hypsibioidea Pilato, 1969
Family Hypsibiidae Pilato, 1969
Subfamily Hypsibiinae Pilato, 1969
Genus Hypsibius Ehrenberg, 1848
Hypsibius cf. dujardini
Taxonomic summary
Material examined: Mexico: Mexico City: Cantera Oriente (19°19’00.5” N, 99°10’21.5” W, 2,260 m asl), 3 specimens (CNAC-Tar000224 - CNAC-Tar000226). Coll. D. López and G. Montiel.
Habitat: high elevation xerophilic scrub. Elevation: 2,260 m asl.
Microhabitat: mosses of the species Amblystegium varium (Hedw.) Lindb. collected on a rock.
Remarks. The specimen traits correspond to the redescription of Hypsibius dujardini (Doyère, 1840) made by Gąsiorek, Stec, Morek et al. (2018). However, we cannot confirm the presence or absence of cuticular bars on legs I-III in all specimens. Therefore, due to the lack of a greater number of specimens for morphometric measurements and for sequencing of genetic material, this species cannot be identified with certainty.
Subfamily Pilatobiinae Bertolani, Guidetti, Marchioro, Altiero, Rebecchi and Cesari, 2014
Genus NotahypsibiusTumanov, 2020
Notahypsibius pallidoidesPilato, Kiosya, Lisi, Inshina and Biserov, 2011
Taxonomic summary
Type locality: Ukraine.
Material examined: Mexico: Mexico City: Cantera Oriente (19°19’00.5” N, 99°10’21.5” W; 2,260 m asl), 10 specimens (CNAC-Tar000214 - CNAC-Tar000223). Coll. D. López and G. Montiel.
Habitat: high elevation xerophilic scrub. Altitude: 2,260 m asl.
Microhabitat: mosses of the species Orthostichella rigida (Müll. Hal.) B.H. Allen & Magill collected on a rock.
Remarks. The specimens examined present Ramazzottius-like claws (Fig. 2E), a smooth cuticle, the pt of the stylet support insertion point between 56.9-57.8%, the eyes are present, a pharyngeal bulb with 2 elongated macroplacoids and a minute dot-like septulum is present (Fig. 2E). The accessory points on primary branches and the lunules under the claws are present (Fig. 2F). The cuticular bars on all legs are absent. Specimen traits correspond to the original description of Notahypsibius pallidoides by Pilato et al. (2011). Also, they were compared with the redescription by Tumanov (2020). The species represents a new record for Mexico.
Family Ramazzottiidae Sands, McInnes, Marley, Goodall-Copestake, Convey and Linse, 2008
Genus Ramazzottius Binda & Pilato, 1986
Ramazzottius cf. oberhaeuseri
Taxonomic summary
Material examined: Mexico: Mexico City: Cantera Oriente (19°19’00.5” N, 99°10’21.5” W; 2,260 m asl), 11 specimens (CNAC-Tar000227 - CNAC-Tar000237) and 3 eggs (CNAC-Tar000282 - CNAC-Tar000284). Coll. D. López, G. Montiel, L. Piña and M. Hernández.
Habitat: high elevation xerophilic scrub. Elevation: 2,260 m asl.
Microhabitat: lichens of the species Heterodermia sp. collected on a tree.
Remarks. The specimens and eggs traits correspond to the redescription of Ramazzottius oberhaeuseri (Doyère, 1840) made by Stec, Morek et al. (2018). However, due to the lack of a greater number of specimens for morphometric measurements and for sequencing of genetic material, this species cannot be identified with certainty.
Superfamily Macrobiotoidea Thulin, 1928
Family Macrobiotidae Thulin, 1928
Genus Minibiotus R.O. Schuster, 1980
Minibiotus cf. continuus
Taxonomic summary
Material examined: Mexico: Mexico City: Cantera Oriente (19°19’00.5” N, 99°10’21.5” W; 2,260 m asl), 7 specimens (CNAC-Tar000282 - CNAC-Tar000288) and 3 eggs (CNAC-Tar000289 - CNAC-Tar000291). Coll. D. López and G. Montiel.
Habitat: high elevation xerophilic scrub. Elevation: 2,260 m asl.
Microhabitat: mosses of the species Syntrichia amphidiacea (Müll. Hal.) R.H. Zander collected on a tree.
Remarks. The traits of the specimens and eggs correspond to the original description of Minibiotus continuus Pilato and Lisi, 2006 (Pilato & Lisi, 2006). However, the eggs examined differ in size from the egg in the original description, which has a diameter of 46.2 µm excluding the processes, and 52.2 µm including them. In contrast, the eggs examined in the present study the diameter is between 42.5 - 44.3 µm excluding the processes and between 49.8 - 51.9 µm including them. Therefore, due to the lack of a greater number of specimens for morphometric measurements, this species cannot be identified with certainty.
Genus ParamacrobiotusGuidetti, Schill, Bertolani, Dandekar and Wolf, 2009
Paramacrobiotus gadaboutiKayastha, Stec, Mioduchowska and Kaczmarek, 2023 (Fig. 2G-I)
Taxonomic summary
Type locality: Portugal.
Material examined: Mexico: Mexico City: Cantera Oriente (19°19’00.5” N, 99°10’21.5” W; 2,260 m asl), 7 specimens (CNAC-Tar000204 - CNAC-Tar000210), 3 eggs (CNAC-Tar000211 - CNAC -Tar000213) and 2 eggs were prepared for SEM. Coll. D. López and G. Montiel.
Habitat: high elevation xerophilic scrub. Elevation: 2,260 m asl.
Microhabitat: mosses of the species Syntrichia amphidiacea collected on a tree.
Remarks. The specimens examined lack eyes, and present a smooth cuticle, a pt of the stylet support insertion point between 77.2-80.5%, granulation on the external surface of legs I-III, smooth lunules under all claws and accessory points on primary branches. The eggs are areolated, of richtersi type with a single ring of 10-12 areolae around each process. The top endings of the processes present cap like structures. Specimens and eggs traits correspond to the original description of Paramacrobiotus gadabouti (Kayastha, Stec et al., 2023), also they were examined following the diagnostic key by Kayastha, Mioduchowska et al. (2023). In addition to the type locality in Madeira Island, Portugal, this species has also been recorded in Australia, France and Tunisia (Kayastha, Stec et al., 2023). A new record for Mexico.
Paramacrobiotus puma López-Sandoval, Montiel-Parra and Pérez sp. nov.
Table 3 Measurements (in μm), and values of the pt index, of some structures of the holotype and paratypes of Paramacrobiotus puma sp. nov.
| Character | N | Range | Mean | Sd | Holotype | ||||
|---|---|---|---|---|---|---|---|---|---|
| µm | pt | µm | pt | µm | pt | µm | pt | ||
| Body length | 25 | 364-678 | 814-1106 | 470 | 944 | 76 | 75 | 576 | 958 |
| Buccal tube | |||||||||
| Length | 25 | 36-61.3 | - | 49.7 | 7.2 | - | 60.1 | - | |
| Stylet support insertion point | 25 | 27.2-46.5 | 74.4-81.2 | 38.6 | 77.8 | 5.3 | 1.4 | 45.9 | 76.3 |
| External width | 25 | 5.6-11.8 | 15.6-25.2 | 9.6 | 19.3 | 1.6 | 1.2 | 10.5 | 17.5 |
| Internal width | 25 | 4.3-9.7 | 11.6-20 | 7.4 | 14.9 | 1.3 | 1.2 | 8.1 | 13.5 |
| Ventral lamina length | 15 | 21.7-40.3 | 54.8-75.5 | 30 | 62 | 5.6 | 3 | 34 | 56.6 |
| Placoid lengths | |||||||||
| Macroplacoid 1 | 25 | 5.5-12.7 | 13.8-20.7 | 8.6 | 17.2 | 1.9 | 1.5 | 11.7 | 19.5 |
| Macroplacoid 2 | 25 | 4.9-9.6 | 10.5-18.4 | 6.6 | 13.3 | 1.4 | 1 | 8.6 | 14.3 |
| Macroplacoid 3 | 25 | 5.7-13 | 15.8-21.5 | 9.5 | 19 | 2.2 | 1.8 | 12.8 | 21.3 |
| Macroplacoid row | 25 | 18-35.9 | 49.4-67.3 | 27.8 | 55.8 | 5 | 2.5 | 33.4 | 55.6 |
| Claw 1 lengths | |||||||||
| External primary branch | 25 | 8.8-17.6 | 16.8-31.4 | 12.8 | 25.9 | 1.8 | 1.8 | 14.4 | 24 |
| External secondary branch | 22 | 7-12.8 | 13.4-26.2 | 9.6 | 19.2 | 1.4 | 2 | 11.2 | 18.6 |
| Internal primary branch | 23 | 6.4-14.8 | 12.2-28.9 | 11.6 | 23.5 | 1.7 | 1.7 | 14.1 | 23.5 |
| Internal secondary branch | 21 | 3.2-11.7 | 6.1-24.7 | 8.2 | 16.6 | 1.3 | 2 | 10.4 | 17.3 |
| Claw 2 lengths | |||||||||
| External primary branch | 24 | 10.3-17.4 | 22-31.4 | 13.1 | 26.4 | 2.1 | 2.6 | 15.1 | 25.1 |
| External secondary branch | 25 | 6.4-12.9 | 12.2-25.3 | 9.8 | 19.7 | 1.8 | 2.1 | 12.8 | 21.3 |
| Internal primary branch | 25 | 7.2-16 | 13.8-28.9 | 12 | 24.1 | 2.1 | 1.8 | 14.8 | 24.6 |
| Internal secondary branch | 25 | 5.6-12.8 | 10.7-23.6 | 9.3 | 18.7 | 1.9 | 2.2 | 11.6 | 19.3 |
| Claw 3 lengths | |||||||||
| External primary branch | 24 | 11.2-18.4 | 24.3-31.9 | 14.1 | 28.4 | 2 | 1.4 | 16 | 26.6 |
| External secondary branch | 23 | 8-13.2 | 15.7-24.5 | 10.3 | 20.7 | 1.7 | 1.5 | 13 | 21.6 |
| Internal primary branch | 23 | 9.6-17.6 | 23.6-31.5 | 13.3 | 26.9 | 2.1 | 2.1 | 15.2 | 25.3 |
| Internal secondary branch | 22 | 7.7-12.6 | 17-22.6 | 9.6 | 19.5 | 1.5 | 1.5 | 11.2 | 18.6 |
| Claw 4 lengths | |||||||||
| Anterior primary branch | 24 | 10.2-20.8 | 19.5-33.9 | 14.7 | 29.6 | 2.3 | 1.8 | 17.6 | 29.3 |
| Anterior secondary branch | 23 | 5.6-15.2 | 10.7-24.8 | 10.6 | 21.2 | 2 | 1.7 | 13.6 | 22.6 |
| Posterior primary branch | 25 | 11.4-21.6 | 21.7-35.2 | 15.6 | 31.4 | 2.5 | 1.4 | 19.2 | 31.9 |
| Posterior secondary branch | 24 | 7.2-16 | 13.8-27.4 | 11.5 | 23.2 | 1.7 | 1.7 | 13.1 | 21.8 |
N = Number of specimens/structures measured; Range = the smallest and the largest structure among all measured specimens; Sd = standard deviation.
Table 4 Measurements (μm) of morphological structures of eggs of Paramacrobiotus puma sp. nov.
| Character | N | Range | Mean | Sd |
|---|---|---|---|---|
| Diameter of egg without processes | 22 | 72-107.1 | 89.9 | 8.1 |
| Diameter of egg with processes | 22 | 94.4-131.9 | 117.1 | 10.4 |
| Process height | 66 | 11.2-27 | 16.5 | 3.2 |
| Process base width | 66 | 9.9-21.1 | 15.7 | 2.3 |
| Process base/height ratio | 66 | 49%-144% | 97% | 18% |
| Distance between processes | 66 | 5.6-10.4 | 7.8 | 1.1 |
| Number of processes on the egg circumference | 19 | 12-15 | 13.1 | 0.8 |
N = Number of eggs/structures measured; Range = the smallest and the largest structure among all measured specimens; Sd = standard deviation.
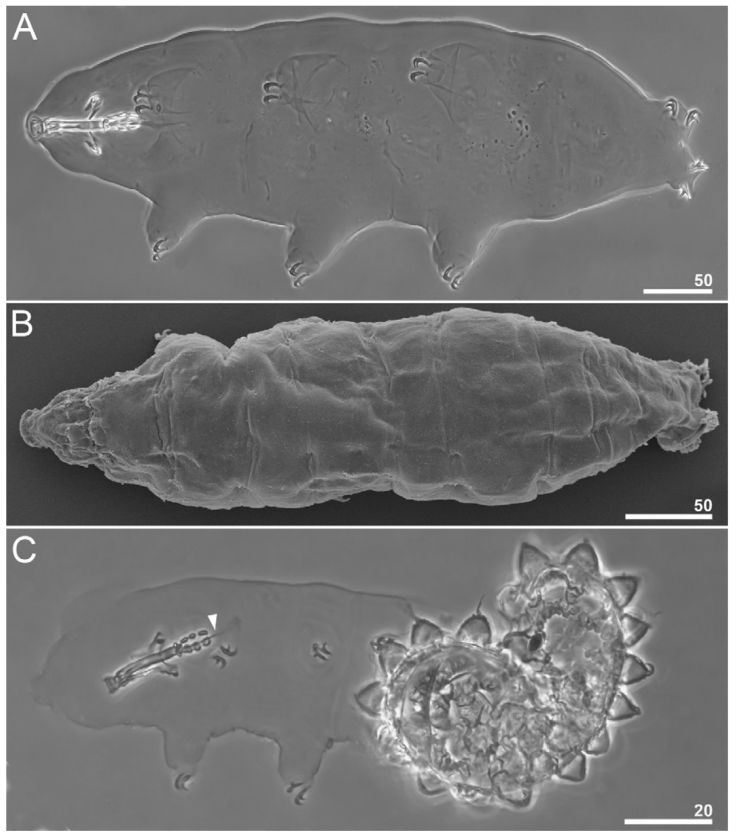
Figure 3 Paramacrobiotus puma sp. nov. Habitus: A, dorso -ventral projection of the entire animal (holotype, PCM); B, dorsal view of the entire animal (paratype, SEM); C, juvenile hatching from the egg (paratype); arrowhead indicates a sclerified line after the third macroplacoid. Scale bars = μm.
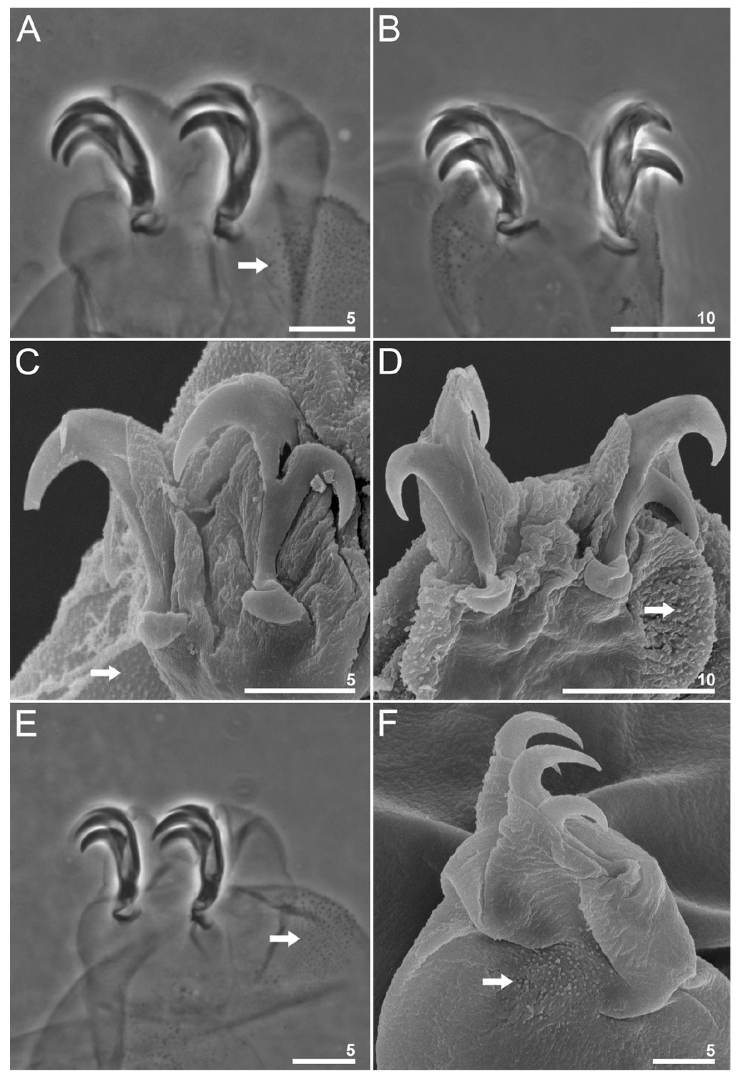
Figure 4 Paramacrobiotus puma sp. nov. Claws and leg granulation: A-B, claws II and IV, respectively (holotype PCM); C-D, claws II and IV, respectively, with smooth lunules seen in SEM (paratype); E-F, claws III seen in PCM (holotype) and SEM (paratype) respectively; arrows indicate the granulation on the external surface of the legs. Scale bars = μm.
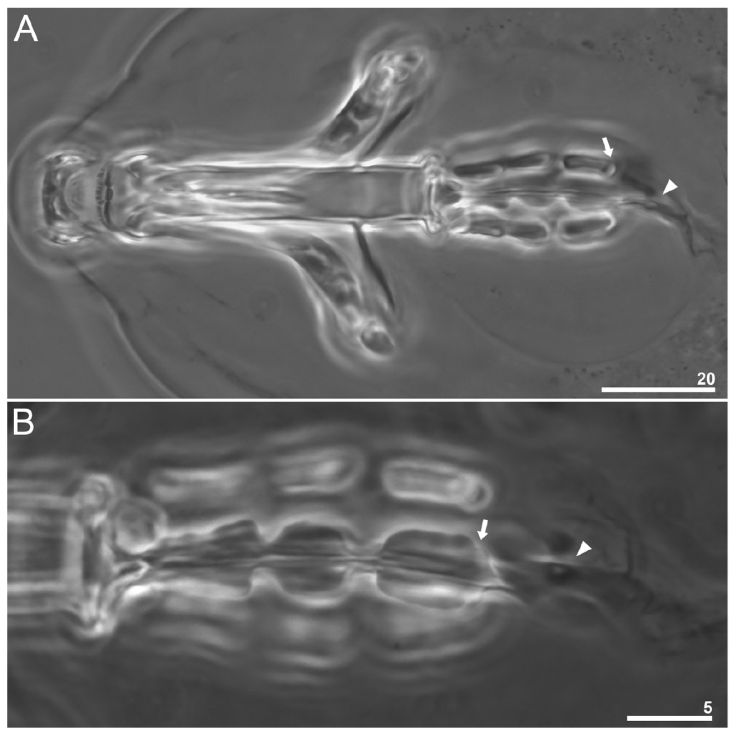
Figure 5 Paramacrobiotus puma sp. nov. Buccopharyngeal apparatus (dorso-ventral projection in PCM): A, general view (paratype); B, dorsal placoids (paratype); arrows indicate the subterminal constriction in the third macroplacoid; arrowhead indicates a sclerified line and a rudimentary microplacoid-like thickening after the third macroplacoid. Scale bars = μm.
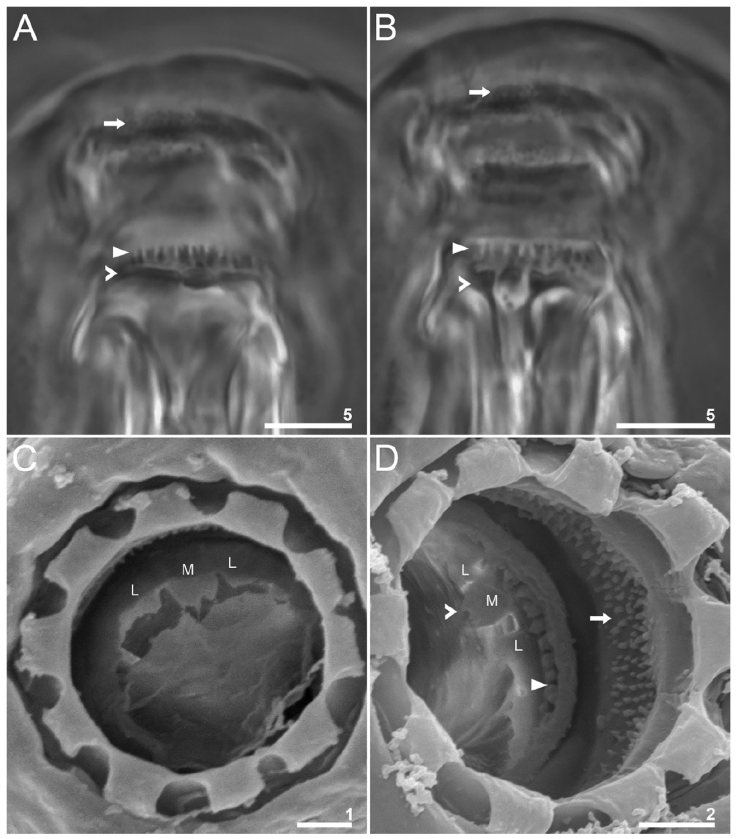
Figure 6 Paramacrobiotus puma sp. nov. Oral cavity armature: A-B, dorsal and ventral views, respectively (paratype, PCM); C-D, dorsal and ventral views, respectively (paratypes, SEM); the arrow indicates teeth of the first band; flat arrowheads indicate teeth of the second band; indented arrowheads indicate teeth of the third band; letters indicate lateral (L) and median (M) crests. Scale bars = μm.
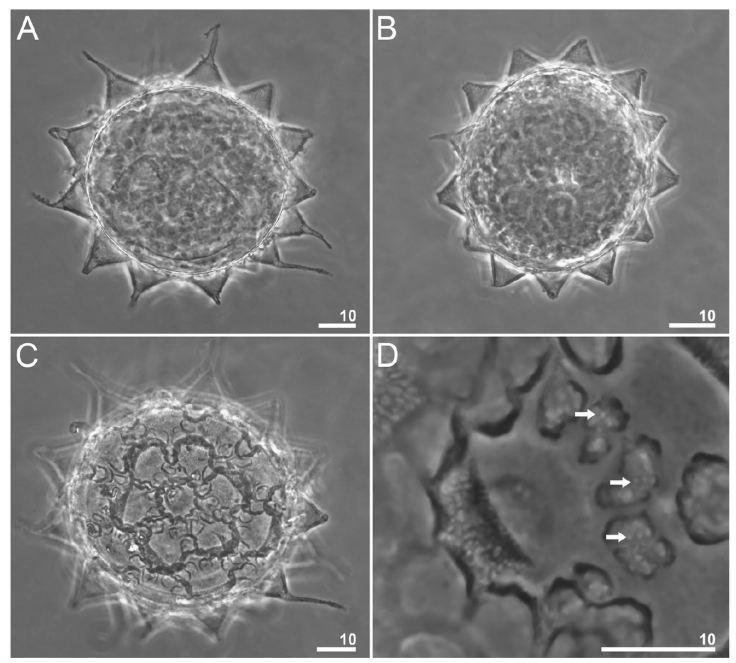
Figure 7 Paramacrobiotus puma sp. nov. Eggs seen in PCM: A-B, midsection; C, the surface of the egg seen in PCM; D, a closer look at the areolation around a process; the arrows indicate the smaller areoles. Scale bars = μm.
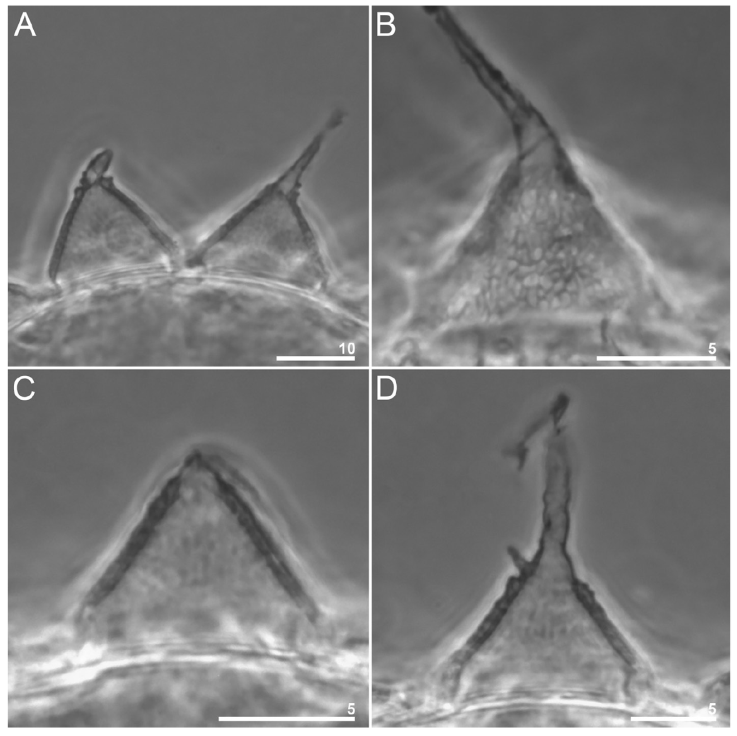
Figure 8 Paramacrobiotus puma sp. nov. Egg processes morphology seen in PCM: A, midsection of 2 processes; B, reticulum within the process walls; C-D, midsection of 2 processes with different apices. Scale bars = μm.

Figure 9 Paramacrobiotus puma sp. nov. Egg seen in SEM: A-B, chorion; C, processes; D, areolae between the processes; E-F, a closer look at the areolation between 2 processes; the arrows indicate the pores inside the areolae. Scale bars = μm.
http://zoobank.org/urn:lsid:zoobank.org:act:4C178E55-DA78-4EFD-88A5-186B7B716A72
Taxonomic summary
Type locality: Cantera Oriente (19°19’00.5” N, 99°10’21.5” W; 2,260 m asl). Mexico City, Mexico.
Material examined: Mexico: Mexico City: Cantera Oriente (19°19’00.5” N, 99°10’21.5” W; 2,260 m asl). Coll. D. López, G. Montiel, L. Piña and M. Hernández.
Microhabitat: lichens of the species Heterodermia cf. tremulans, collected from a tree trunk.
Type material. Holotype and 55 paratypes (32 specimens and 23 eggs). Additionally, 2 specimens and 2 eggs were prepared for SEM photographs, and 2 specimens were processed for DNA sequencing.
Holotype (CNAC-TTar000011). Mexico City, Mexico: Reserva Ecológica del Pedregal de San Ángel, Cantera Oriente (19°19’00.5” N, 99°10’21.5” W; 2,260 m asl).
Paratypes. (CNAC- TTar000001 - CNAC-TTar000056). The same data as for the holotype.
Type specimen’s depositories. Holotype (CNAC-TTar000011) and 51 paratypes (30 specimens and 21 eggs) (CNAC-TTar000001 - CNAC-TTar000056) are deposited at the Tardigrada Collection associated with the National Collection of Mites (CNAC) of the Instituto de Biología, UNAM, Mexico City (Mexico). Additionally, 4 paratypes (2 specimens and 2 eggs) (slides CNAC-TTar000030, CNAC-TTar000032, CNAC-TTar000050 and CNAC-TTar000053) are deposited in the Department of Animal Taxonomy and Ecology, Institute of Environmental Biology, Adam Mickiewicz University, Poznań, Poland, and 4 paratypes (2 specimens and 2 eggs) (slides CNAC-TTar000029, CNAC-TTar000031, CNAC-TTar000040 and CNAC-TTar000043) are deposited in the Institute of Zoology and Biomedical Research, Jagiellonian University, Kraków, Poland.
Etymology. The new species is named after the Puma, the official mascot which is the emblem of the National Autonomous University of Mexico (UNAM).
Description (measurements and statistics inTable 3).
White body in live specimens, transparent in specimens mounted in Hoyer’s medium (Fig. 3A-C). Eyes are present in live adult animals (present in 18 of the 25 specimens examined mounted in Hoyer’s medium) and absent in a hatching specimen examined (Fig. 3C). Cuticle smooth, without gibbosities, papillae, pores, spines, or ornaments. Small areas of granulation in the first 3 pairs of legs are present, specifically on the external surfaces near the claw bases, and as for the hind legs, the granulation is extended from the claws onto the entire dorsal surface of the legs (Fig. 4). Claws robust, of the hufelandi type (Fig. 4A, B). Smooth lunules are present under all claws (Fig. 4C, D). Accessory points on primary branches are present. Cuticular bars under the claws are absent.
Mouth antero-ventral with 10 peribuccal lamellae. Buccopharyngeal apparatus of the Macrobiotus type with ventral lamina present (Fig. 5A). The oral cavity armature is composed of 3 bands of teeth (Fig. 6). The first band of teeth consists of small cones (granules in PCM) positioned at the base of the buccal lamellae. The second band of teeth is composed of larger cones arranged in one row around the oral cavity and positioned in the rear of the oral cavity between the ring fold and the third band of teeth. The third band of teeth is positioned just before the buccal tube opening and is composed of dorsal and ventral transversal ridges, organized into 2 large lateral ridges and a smaller median ridge (Fig. 6C, D).
The pharyngeal bulb is spherical, with triangular apophyses and 3 rod-shaped macroplacoids (Fig. 5B). Macroplacoid length sequence is 2 < 1 < 3. The first macroplacoid is without constrictions but narrower anteriorly. The second macroplacoid is of uniform width and without constrictions. The third macroplacoid presents a sub-terminal constriction. Microplacoid is absent, however, a sclerified line and a rudimentary microplacoid-like thickening are present after the third macroplacoid in adult specimens (Fig. 5A, B). In the hatching specimen examined, the microplacoid-like thickening is also present (Fig. 3C).
Eggs (measurements and statistics in Table 4). Eggs of the richtersi type. Laid free, white, spherical, and with 12-15 cone-shaped processes on the circumference with a variable termination (Figs. 3C; 7A-D; 9A- F). The apices can be very short or thin and long (Fig. 8A-D). The labyrinth layer between the walls of the processes is visible under PCM as a reticular pattern (Fig. 8B). Between 12 and 14 areolae around each process. The internal surface of the areolae is sculpted with a reticular pattern with pores (Figs. 7D; 9E, F).
DNA sequences. We obtained DNA sequences for all 4 molecular markers from 1 hologenophore (voucher number: CNAC-TTar000033). The sequence length and GenBank accession number of each marker are as follows: 18s rRNA (GenBank: PP416751), 1632 -bp long; 28s rRNA (GenBank: PP416752), 737-bp long; ITS-2 (GenBank: PP416753), 419-bp long; COI (GenBank: PP414782), 631-bp long.
Phenotypic differential diagnosis. The genus Paramacrobiotus is divided into 2 morphologically distinct species groups: the richtersi group (species with a microplacoid in the pharynx) and the areolatus group (species without a microplacoid or with rudimentary structures in the place of microplacoid in the pharynx). Also, there are 7 types of eggs, being the areolatus and richtersi the most common types (Kaczmarek et al., 2017; Kayastha, Mioduchowska et al., 2023). Since the microplacoid is absent in Paramacrobiotus puma sp. nov., it belongs to the areolatus group along with 12 other species: P. areolatus (Murray, 1907), P. bifronsBrandoli, Cesari, Massa, Vecchi, Rebecchi and Guidetti, 2024, P. centesimus (Pilato, 2000), P. csotiensis (Iharos, 1966), P. derkai (Degma, Michalczyk & Kaczmarek, 2008), P. huziori (Michalczyk & Kaczmarek, 2006), P. intiiKaczmarek, Cytan, Zawierucha, Diduszko and Michalczyk, 2014, P. klymenkiPilato, Kiosya, Lisi and Sabella, 2012, P. lachowskaeStec, Roszkowska, Kaczmarek and Michalczyk, 2018, P. spinosusKaczmarek, Gawlak, Bartels, Nelson and Roszkowska, 2017, P. tonollii (Ramazzotti, 1956) and P. walteri (Biserov, 1998) (Biserov, 1998; Brandoli et al., 2024; Degma et al., 2008; Iharos, 1966; Kaczmarek, Michalczyk et al., 2014, 2017; Michalczyk & Kaczmarek, 2006; Murray, 1907; Pilato, 2000; Pilato et al., 2012; Ramazzotti, 1956; Stec, Roszkowska et al., 2018). Moreover, Paramacrobiotus puma sp. nov., presents the richtersi type of egg, and is most similar to 7 species within the areolatus group: P. areolatus, P. centesimus, P. intii, P. klymenki, P. lachowskae, P. spinosus and P. walteri. Nevertheless, the new species can be differentiated specifically from all species mentioned above by the following traits.
Paramacrobiotus puma sp. nov. differs from P. areolatus, restricted only to Svalbard and Greenland (McInnes, 1994; Stec, Krzywański et al., 2020), by the presence of smooth lunules under the IV claws (crenate in P. areolatus) and a different type of egg (richtersi type in P. puma sp. nov. vs. areolatus type in P. areolatus). From P. bifrons, recorded only in Italy (Brandoli et al., 2024), by the presence of smooth lunules under the IV claws (clearly indented larger lunules on legs IV in P. bifrons) and by a different type of egg (richtersi type in P. puma sp. nov. vs. 2 egg types in P. bifrons: areolatus type and csotiensis type). From P. centesimus, recorded only in Brazil and Ecuador (Kaczmarek et al., 2015; Pilato, 2000), by a different type of egg (richtersi type in P. puma sp. nov. vs. areolatus type in P. centesimus), longer egg processes (11.2-27.0 μ m in P. puma sp. nov. vs. 7.0-11.0 μm in P. centesimus), and by larger full egg diameter (94.4-131.9 μm in P. puma sp. nov. vs. 76.0-91.0 μm in P. centesimus). From P. csotiensis, recorded only in Hungary (Iharos, 1966), by a different type of egg (egg processes shape blunt with a transparent cover [csotiensis type] in P. csotiensis vs egg processes shape conical without a transparent cover [richtersi type] in P. puma sp. nov.), and by different egg diameters (diameters with processes of 75-80 μm and without processes of 60-65 μm in P. csotiensis vs 94.4-131.9 μm with processes and of 72.0 -107.1 μm without processes in P. puma sp. nov.). From P. derkai, recorded only in Colombia and Peru (Degma et al., 2008; Kaczmarek et al., 2016), by less protruding accessory points on primary branches, for a different type of egg (richtersi type in P. puma sp. nov. vs huziori type in P. derkai), spaces between neighbor areolae (narrower than areolae widths in P. puma sp. nov. vs. usually broader than the areolae widths in P. derkai), and by longer egg processes (11.2-27.0 μm P. puma sp. nov. vs. 8.0-17.1 μm in P. derkai). From P. huziori, recorded only in Costa Rica (Michalczyk & Kaczmarek, 2006; Kaczmarek, Michalczyk et al., 2014), by leg granulation in aggregations of small granules or cones absent (present in P. huziori), a different type of egg (richtersi type in P. puma sp. nov. vs. huziori type in P. huziori), shorter egg processes (11.2-27 μm in P. puma sp. nov. vs. 20.0-33.0 in P. huziori), and by the number of egg processes (12-15 in P. puma sp. nov. vs. 9-11 in P. huziori). From P. intii, recorded only in Peru (Kaczmarek, Cytan et al., 2014), by the oral cavity armature, showing bands I to III in PCM (only I and II in P. intii), a different type of egg (richtersi type in P. puma sp. nov. vs. areolatus type in P. intii), shorter egg processes (11.2-27 μm in P. puma sp. nov. vs. 15.4-24.4 in P. intii), and by the number of egg processes (12-15 in P. puma sp. nov. vs. 9-10 in P. intii). From P. klymenki, recorded only in Belarus (Pilato et al., 2012), by presenting eyes (absent in P. klymenki), lunules IV smooth (crenate in P. klymenki), a different type of egg (richtersi type in P. puma sp. nov. vs. areolatus type in P. klymenki), and by the number of egg processes (12 -15 in P. puma sp. nov. vs. 10 -11 in P. klymenki). From P. lachowskae, recorded only in Colombia (Stec, Roszkowska et al., 2018), by macroplacoid length sequence (2 < 1 < 3 in P. puma sp. nov. vs. 2 < 3 ≤ 1 in P. lachowskae), a different type of egg (richtersi type in P. puma sp. nov. vs. areolatus type in P. lachowskae), egg processes shape (conical processes with filaments not covered with hairs in P. puma sp. nov. vs. dome-like, wrinkled, and with long flexible spines/filaments covered by fine short hairs in P. lachowskae). From P. spinosus, recorded only in Ecuador (Kaczmarek et al., 2017), by the presence of eyes (absent in P. spinosus), egg processes shape (conical processes with transverse annulations smooth in P. puma sp. nov. vs. transverse annulations associated with short spines in the upper parts of egg processes in P. spinosus), by the number of egg processes (12-15 in P. puma sp. nov. vs. 10-11 in P. spinosus), and by the number of areolae on egg surface (between 12 and 14 areolae around each process in P. puma sp. nov. vs. 10 areolae in P. spinosus). From P. tonollii, recorded in Lapland (Finland), USA, and Canada (McInnes, 1994), by a different type of egg (richtersi type in P. puma sp. nov. vs. tonollii type in P. tonollii) and by the number of egg processes (12 -15 in P. puma sp. nov. vs. 8-10 in P. tonollii). And from P. walteri, known only from Russia (Biserov, 1998), by the presence of smooth lunules under claws IV (dentate in P. walteri), and by a different type of egg (richtersi type in P. puma sp. nov. vs. areolatus type in P. walteri).
Genotypic differential diagnosis. The ranges of uncorrected genetic p- distances from the most to the least conservative between the new species and other species of the genus Paramacrobiotus for which sequences are available from GenBank, are as follows: 18S rRNA: 0.06-5.02% (1.66% on average), with the most similar being Paramacrobiotus lachowskae from Magdalena Province, Colombia (MF568532), and the least similar being a haplotype attributed to P. danielae (Pilato, Binda, Napolitano & Moncada, 2001) from undetermined location (MZ081363). In the 28S rRNA: 0.00-8.57% (5.98% on average), being identical to Paramacrobiotus lachowskae from Magdalena Province, Colombia (MF568533), and the least similar being P. tonollii (Ramazzotti, 1956) from the East Tennessee State University campus, USA (MH664963). In the ITS-2: 10.36-35.69% (25.29% on average), with the most similar being Paramacrobiotus lachowskae from Magdalena Province, Colombia (MF568535), and the least similar P. tonollii (Ramazzotti, 1956) from Oregon, USA (GQ403679). In the COI: 11.33-26.94% (21.78% on average), with the most similar being Paramacrobiotus lachowskaeStec, Roszkowska, Kaczmarek and Michalczyk, 2018 from Magdalena Province, Colombia (MF568534), and the least similar being a haplotype attributed to P. richtersi (Murray, 1911) from China (unpublished) (GU339056).
Phylogenetic analysis. The phylogenetic reconstruction performed using BI and ML methods resulted in trees with similar topology and mostly well supported nodes, although the lowest support values corresponded to the ML tree (Fig. 10). The results show that the Paramacrobiotus richtersi morphogroup was recovered as paraphyletic, and the areolatus morphogroup as polyphyletic, as the sequences of the new species obtained in this study clustered together with P. lachowskae from Colombia within the clade of the richtersi morphogroup. These results are largely consistent with the phylogenies previously presented by Stec, Krzywański et al. (2020) and Kayastha, Stec et al. (2023). Paramacrobiotus areolatus and P. tonolli from the USA are found at the base of the areolatus morphogroup clade and interestingly, P. cf. klymenky IT.048 from Italy was clustered with P. bifrons also from Italy, being P. cf. klymenky PT.006 from Portugal a sister lineage to that cluster.
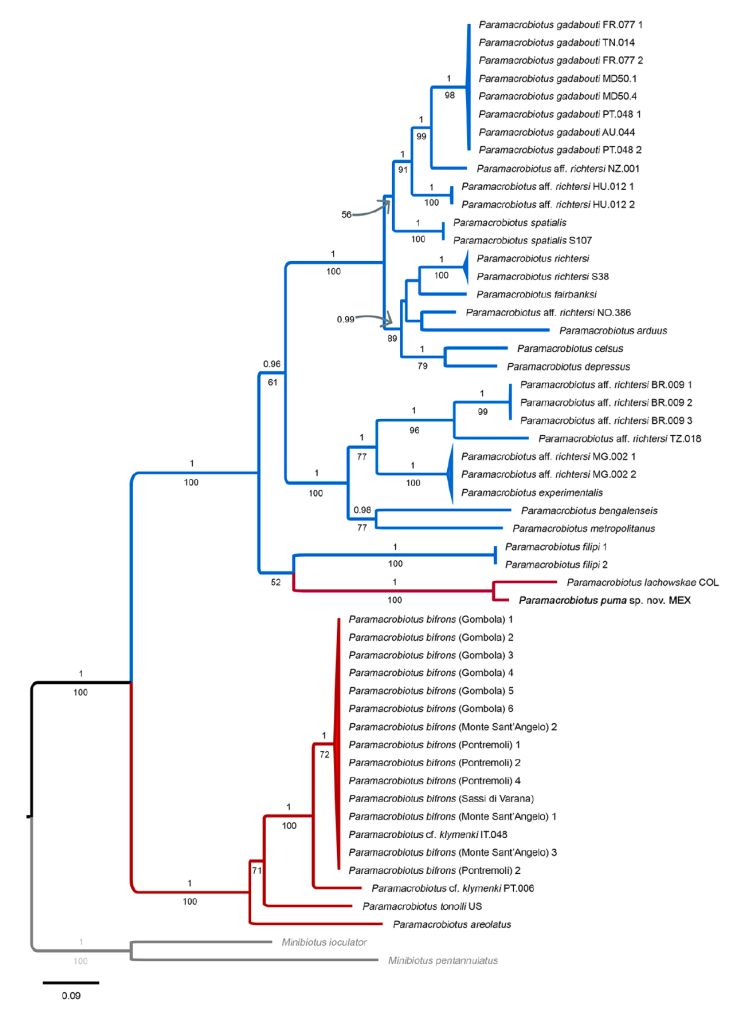
Figure 10 Phylogeny constructed from concatenated sequences of the genus Paramacrobiotus (18S rRNA + 28S rRNA + ITS-2 + COI; Table 2). Numbers above branches indicate Bayesian posterior probabilities values (≥ 0.90), while number below branches indicate bootstrap support values (≥ 50). In agreement with previous phylogenies (Basu et al., 2023; Kayastha, Stec et al., 2023; Stec, Krzywański et al., 2020), taxa of the richtersi and areolatus morphogroups are indicated by blue and red branches, respectively. The outgroup is indicated in gray font. The new species is in bolded font. The scale bar represents substitutions per position.
Discussion
To date, 83 species of tardigrades have been reported for Mexico. Particularly for the genus Milnesium, 5 species have been previously reported: Milnesium barbadosense Meyer and Hinton, 2012 (Moreno-Talamantes et al., 2019), M. cassandrae Moreno-Talamantes, Roszkowska, García-Aranda, Flores-Maldonado and Kaczmarek, 2019 (Moreno-Talamantes et al., 2019), M. fridae Moreno-Talamantes, León-Espinosa, García-Aranda, Flores-Maldonado and Kaczmarek, 2020 (Moreno-Talamantes et al., 2020), M. sp. (Moreno-Talamantes et al., 2020) and M. tardigradum tardigradum Doyère, 1840 (Schuster, 1971; Beasley, 1972; Kaczmarek et al., 2011; Moreno-Talamantes et al., 2019, 2020). However, currently, the geographical distribution of Milnesium tardigradum is restricted to Central and Western Europe (Kaczmarek et al., 2011; Michalczyk et al., 2012a, b; Morek et al., 2018), so the records provided by Schuster (1971) and Beasley (1972) are doubtful and it is necessary to inspect the specimens collected in Mexico since they are probably a different species. Milnesium longiungue and M. cf. reductum represent a new record for Mexico and the number of species of the genus Milnesium increases to 7.
Regarding the family Hypsibiidae, 4 subfamilies have been recorded: Diphasconinae, Hypsibiinae, Itaquasconinae and Pilatobiinae (Schuster, 1971; Ramazzotti & Maucci, 1983; Moreno-Talamantes et al., 2019; Dueñas-Cedillo et al., 2020). For the subfamily Hypsibiinae, 4 species belonging to the genus Hypsibius are present in the country: H. cf. convergens recorded by Schuster (1971), H. cf. microps and H. cf. pallidus recorded by Dueñas-Cedillo et al. (2020) and H. pallidus Thulin, 1911 recorded by Ramazzotti and Maucci (1983), and with the record of H. cf. dujardini, there are 5 species recorded for the genus. And as for the subfamily Pilatobiinae, Notahypsibius pallidoides represents the first species of this genus recorded for Mexico.
Specifically for the genus Paramacrobiotus, P. areolatus (Murray, 1907) and P. richtersi (Murray, 1911) were recorded by Schuster (1971), being the only 2 species of the genus that are present in Mexico. However, these records should be considered doubtful, because although both species were previously considered cosmopolitan (Kaczmarek et al., 2011, 2017), they are currently within 2 morphogroups of closely related species and their distribution is very restricted. Currently, Paramacrobiotus areolatus is restricted to Svalbard and Greenland (Stec, Krzywański et al., 2020), and P. richtersi to Ireland and Finland (Kayastha, Mioduchowska et al., 2023). Furthermore, in addition to the diagrams showing the buccopharyngeal apparatus and the egg type, Schuster (1971), did not provide more details of the chorion or the egg ornamentation. Therefore, it is necessary to inspect the specimens registered in Mexico that were attributed to P. areolatus and P. richtersi by Schuster, since they are most likely different species. Paramacrobiotus gadabouti is a new record for Mexico, and with the description of P. puma sp. nov., there are 4 species of the genus recorded for the country.
Phylogenetic analysis. The genus Paramacrobiotus has been studied on several occasions. Guidetti et al. (2009), erected the genus by separating it from Macrobiotus Schultze, 1834. Subsequently, the redescriptions of Paramacrobiotus richtersi, the nominal taxon (Guidetti et al., 2019) and P. areolatus (Stec, Krzywański et al., 2020), contributed significantly by providing detailed morphological and molecular data from both species and several additional new species described. Recently, Basu et al. (2023) and Kayastha, Stec et al. (2023) described Paramacrobiotus bengalenseis and P. gadabouti, respectively, contributing to new phylogenetic hypotheses. Basu et al. (2023) demonstrated the monophyly of the richtersi group but found the areolatus group as paraphyletic, Kayastha, Stec et al. (2023) found the representatives of the areolatus group formed a paraphyletic group caused by P. lachowskae which was clustered together with the richtersi morphogroup.
In the present study, we found a tree topology very similar to those previously published by Stec, Krzywański et al. (2020) and Kayastha, Stec et al. (2023), however, the richtersi group was recovered as paraphyletic and the areolatus group as polyphyletic (Fig. 10). As for Paramacrobiotus puma sp. nov., it was clustered together with P. lachowskae from Colombia, both species being the only neotropical representatives within the areolatus morphogroup along the phylogenies. These results indicate that Paramacrobiotus puma sp. nov. is very close to P. lachowskae, as seen in the genetic distances (p-distances), particularly for the ribosomal marker 28s rRNA, which proved to be a similar haplotype between both species (see results above). Nevertheless, morphologically, several differences are found between both species, such as the macroplacoid length sequence, the type of eggs and the egg processes shape (see differential diagnosis).
Finally, according to the phylogenetic tree obtained, we recovered Paramacrobiotus cf. klymenky IT.048 from Italy, clustered with P. bifrons, which indicates that they may represent closely related species. Also, Paramacrobiotus cf. klymenky PT.006 is recovered as a sister taxon of that cluster (Fig. 10). In our study, the p-distances between Paramacrobiotus cf. klymenky IT.048 and P. bifrons, showed to be low (between 0.53-2.72%) for the COI marker, but very variable regarding the ITS-2 marker (between 0.55-6.55%) (Supplementary material, SM2). Stec, Krzywański et al. (2020) studied Paramacrobiotus cf. klymenky IT.048 along with P. cf. klymenky PT.006, where despite being morphologically identical (suggesting they were a single species), both species presented discordant genetic distances between the ITS-2 and COI markers. Moreover, regardless of the genetic distance method used, for the COI marker they were different species and as for the ITS-2 marker, they turned out to be the same species. Nevertheless, although in the different phylogenies they are clustered together (Basu et al., 2023; Kayastha, Stec et al., 2023; Stec, Krzywański et al., 2020), whether the Italian and the Portuguese populations are different, or the same species is difficult to answer so far, since a greater number of studies with an integrative approach are still needed to gradually gain a better resolution within this genus.

