Artículos
Population dynamics of Naegleria fowleri genotype 2 in natural aquatic environments in Sonora, Mexico, throughout the year
Dinámica poblacional del genotipo 2 de Naegleria fowleri en ambientes acuáticos naturales de Sonora, México, a lo largo del año
Lares-Jiménez, Luis Fernando1
 http://orcid.org/0000-0002-8314-7512
http://orcid.org/0000-0002-8314-7512
Rodríguez-Anaya, Libia Zulema2
http://orcid.org/0000-0003-1643-3938
Alfaro-Sifuentes, Rosalía3
http://orcid.org/0000-0002-5366-7402
Gonzalez-Galaviz, José Reyes2
http://orcid.org/0000-0001-7235-9328
Borquez-Román, Manuel Alejandro3
http://orcid.org/0009-0009-0111-0387
Otero-Ruiz, Alejandro3
http://orcid.org/0000-0002-8793-7204
González-Peraza, Fernando4
Ibarra-Gámez, José Cuauhtémoc1
http://orcid.org/0000-0001-5601-7598
Casillas-Hernández, Ramón1
http://orcid.org/0000-0003-1977-5298
Lares-Villa, Fernando13*
http://orcid.org/0009-0001-1576-6098
http://orcid.org/0000-0002-8314-7512Rodríguez-Anaya, Libia Zulema2
http://orcid.org/0000-0003-1643-3938Alfaro-Sifuentes, Rosalía3
http://orcid.org/0000-0002-5366-7402Gonzalez-Galaviz, José Reyes2
http://orcid.org/0000-0001-7235-9328Borquez-Román, Manuel Alejandro3
http://orcid.org/0009-0009-0111-0387Otero-Ruiz, Alejandro3
http://orcid.org/0000-0002-8793-7204González-Peraza, Fernando4
Ibarra-Gámez, José Cuauhtémoc1
http://orcid.org/0000-0001-5601-7598Casillas-Hernández, Ramón1
http://orcid.org/0000-0003-1977-5298Lares-Villa, Fernando13*
http://orcid.org/0009-0001-1576-6098
-
Publication dates-
May 26, 2025
Jan-Dec , 2025
- Article in PDF
- Article in XML
- Automatic translation
- Send this article by e-mail
- Share this article +
Abstract
We carried out monthly samplings to determine the concentration of Naegleria fowleri in four natural water bodies of the Yaqui Valley, Sonora, Mexico. We measured the temperature, dissolved oxygen, and pH of the water. The water samples were shaken and processed to determine the concentration using the most probable number (MPN) method and seeded on non-nutritive agar plates with Escherichia coli. Each amoeba detected was isolated in a new plate to continue with the identification. MPN tables were used for the amoeba count, and the identity of the genus Naegleria and N. fowleri was used using specific primers and end-point PCR. We identified N. fowleri in all four water bodies during the summer and fall seasons, with 70 MPN L-1 as the highest concentration. We selected seven N. fowleri strains for sequencing and genotyping; all belonged to type 2. Through the results, we can affirm that the variations in the number of N. fowleri recorded throughout the year at the different sites studied are influenced by different biotic or abiotic factors, in addition to temperature, dissolved oxygen, and pH determined in this study. This makes it difficult to predict their presence in natural aquatic environments with relatively low populations.
Keywords::
Naegleria, free-living amoebae, environmental distribution, primary amoebic meningoencephalitis, meningitis, thermophilic, N. fowleri genotype 2
Introduction
Naegleria fowleri received a new species status in 1970 due to its pathogenicity in humans (De Jonckheere, 2011). This free-living amoeba, also known as “the brain-eating amoeba” (Ruszkiewicz et al., 2019), is the causative agent of primary amoebic meningoencephalitis (PAM), an acute and usually fatal disease (Gompf and Garcia, 2019). PAM occurs primarily in children and adolescents with a history of swimming or diving in various aquatic environments. N. fowleri can reach the central nervous system (CNS) through water entering the nose (Cope et al., 2019). Using devices for washing the nose for medical or religious reasons with water contaminated with N. fowleri has been the cause of several deaths from PAM in adults (Yoder et al., 2012). This pathogen is a thermophilic ameboflagellate isolated worldwide from freshwater lakes, ponds, rivers, hot springs, thermally polluted water, warm groundwater, inadequately treated swimming pools, sewage, in biofilms of drinking water distribution systems, and soil, where it lives by feeding on bacteria and other microbes in the environment. It tolerates temperatures of up to 45 °C and thrives during warmer months when the ambient temperature increases. Naegleria is not found in salt water, like the sea (Morgan et al., 2016; Puzon et al., 2017; CDC, 2024).
-
De Jonckheere, 2011Origin and evolution of the worldwide distributed pathogenic amoeboflagellate Naegleria fowleriInfection, Genetics and Evolution, 2011
-
Ruszkiewicz et al., 2019Brain diseases in changing climateEnviron. Res, 2019
-
Gompf and Garcia, 2019Lethal encounters: The evolving spectrum of amoebic meningoencephalitisIDCases, 2019
-
Cope et al., 2019Response and remediation actions following the detection of Naegleria fowleri in two treated drinking water distribution systems, Louisiana, 2013-2014J Water Health, 2019
-
Yoder et al., 2012Primary amebic meningoencephalitis deaths associated with sinus irrigation using contaminated tap waterClin. Infect. Dis, 2012
-
Morgan et al., 2016Characterization of a drinking water distribution pipeline terminally colonized by Naegleria fowleriEnviron Sci Technol, 2016
-
Puzon et al., 2017Comparison of biofilm ecology supporting growth of individual Naegleria species in a drinking water distribution systemFEMS Microbiol. Ecol, 2017
Most of the current work is focused on N. fowleri in artificial environments, such as drinking water distribution systems rather than natural environments, reporting presence and absence data and how to reduce its presence (Miller et al., 2017; Miller et al., 2018). Several studies have also focused on improving isolation, culture, and identification procedures by molecular methods, including quantitative determinations of N. fowleri from different environments, and how biotic and abiotic factors could explain its presence and distribution (Puzon et al., 2009; Streby et al., 2015; Miller et al., 2015; Moussa et al., 2020).
-
Miller et al., 2017Elimination of Naegleria fowleri from bulk water and biofilm in an operational drinking water distribution systemWater Res, 2017
-
Miller et al., 2018Preferential feeding in Naegleria fowleri; intracellular bacteria isolated from amoebae in operational drinking water distribution systemsWater Res, 2018
-
Puzon et al., 2009Rapid detection of Naegleria fowleri in water distribution pipeline biofilms and drinking water samplesEnviron Sci Technol, 2009
-
Streby et al., 2015Comparison of real-time PCR methods for the detection of Naegleria fowleri in surface water and sedimentParasitol Res, 2015
-
Miller et al., 2015Reduced efficiency of chlorine disinfection of Naegleria fowleri in a drinking water distribution biofilmEnviron Sci Technol, 2015
-
Moussa et al., 2020An optimized most probable number (MPN) method to assess the number of thermophilic free-living amoebae (FLA) in water samplesPathogens, 2020
The current study involved a seasonal search of N. fowleri in three natural surface bodies of water and one thermal spring from Yaqui Valley, the cradle of the green revolution, in northwestern Mexico. The objective was to determine the number and distribution of amoebas, and assess whether the presence of N. fowleri correlates with seasonal factors or other environmental factors and which type of N. fowleri predominates in this region of the eight genotypes recorded worldwide (De Jonckheere, 2011). It is still unclear how often N. fowleri is present in natural surface waters and at what concentration its presence becomes a human health hazard.
-
De Jonckheere, 2011Origin and evolution of the worldwide distributed pathogenic amoeboflagellate Naegleria fowleriInfection, Genetics and Evolution, 2011
Material and methods
Sampling sites
In the study, we included four natural water bodies with recreational activities. La Isleta (27°45’21.46’’ N, 109°54’04.79’’ O), a lagoon called Las Palmas (27°43’58.42’’ N, 109°52’28.56’’ O), a thermal spring called Aguacaliente (27°43’54.87’’ N, 109°50’12.69’’ O) and, the Laguna del Náinari (27°29’42.73’’ N, 109°58’02.81’’ O). The climate in the area is sweltering, with an average annual precipitation of 410 mm (INAFED, 2020).
-
INAFED, 2020,
Sampling
Samples were taken monthly from each site from May 2017 to April 2018. The sampling point was previously established, referring to the presence of organic matter and inert matter as support. Once we selected the sampling spot, rubbed and mixed the waters, we removed any amoeba adhering to the substrate, and we collected 1 L per site in sterile containers. Likewise, the temperature of the water and the environment were measured in situ with a BRANNAN mercury thermometer, dissolved oxygen with a YSI oximeter, salinity with a VITAL SINE refractometer and, in the laboratory, the pH with a potentiometer HANNA (Pernin et al., 1998; Lares-Villa and Hernández-Peña, 2010). We performed all statistical analyses using the STATGRAPHICS Plus software (version 5.1, Statistical Graphics Corp., USA).
-
Pernin et al., 1998Comparative recoveries of Naegleria fowleri amoebae from seeded river water by filtration and centrifugationAppl Environ Microbiol, 1998
-
Lares-Villa and Hernández-Peña, 2010Concentration of Naegleria fowleri in natural waters used for recreational purposes in Sonora, Mexico (November 2007-October 2008)Exp. Parasitol, 2010
Culture on non-nutritive agar with Escherichia coli (NNE)
Bacteriological agar MCD-LAB at 5% was prepared, heated until complete dissolution, and autoclaved at 121 °C for 15 minutes. We then emptied the medium into disposable KLINICUS Petri dishes. After solidifying, we applied 200 µL of live E. coli suspension to each plate and spread it with a Drigalski spatula.
Sample processing to determine the most probable number (MPN)
After stirring to homogenize, we filtered the water samples through 1.2 µm Merck Millipore cellulose membranes using the Micro Filtration Systems filter kit, where five volumes of 100 mL and five volumes of 10 mL were filtered separately (Pernin et al., 1998; Lares-Villa and Hernández-Peña, 2010; Blodgett, 2024). We cut the filters in half and placed them inverted on the same NNE plate to allow amoebae to exit quickly after incubation. Likewise, we planted directly in NNE plates in quintuplicate, 1 and 0.1 mL per sample, leaving them at rest for 20 minutes before incubating the Petri dishes with inverted NNE and in plastic bags to prevent them from drying out. We obtained 80 plaques per month from the four sites and incubated them at 45 °C. The reason for planting four volumes of water samples, 100, 10, 1, and 0.1 mL, is that we did not know the number of amoebae that could exist in the different bodies of water (Pernin et al., 1998; Lares-Villa and Hernández-Peña, 2010).
-
Pernin et al., 1998Comparative recoveries of Naegleria fowleri amoebae from seeded river water by filtration and centrifugationAppl Environ Microbiol, 1998
-
Lares-Villa and Hernández-Peña, 2010Concentration of Naegleria fowleri in natural waters used for recreational purposes in Sonora, Mexico (November 2007-October 2008)Exp. Parasitol, 2010
-
Blodgett, 2024BAM Appendix 2: Most Probable Number from Serial Dilutions, 2024
-
Pernin et al., 1998Comparative recoveries of Naegleria fowleri amoebae from seeded river water by filtration and centrifugationAppl Environ Microbiol, 1998
-
Lares-Villa and Hernández-Peña, 2010Concentration of Naegleria fowleri in natural waters used for recreational purposes in Sonora, Mexico (November 2007-October 2008)Exp. Parasitol, 2010
Isolation and counting of thermophilic amoebae
We examined each culture daily under an inverted Axiovert 135 ZEISS microscope (Carl Zeiss, Gottingen, Germany) for five days and registered the amoebae presence or absence from each NNE plate. After we compared the results with NMP tables to determine the MPN of amoebae per liter from each sampling site, the MPN/L obtained by filtration had a correction multiplied by two, due to loss of amoebae with this concentration method. All plates that did not show growth after five days were considered negative. We collected each amoeba growth that emerged along the two halves of the filter for isolation and subculture. Similarly, each amoeba “colony” was grown separately on seeded plates with 1 and 0.1 mL of the water sample, and transferred to a new NNE plate to prevent further cloning of the amoeba. For this last part, the first observation was before 24 hours of incubation. For counting thermophilic amoebae (TA), only the positive or negative growth of each seeded plate and the corresponding dilution registered were compared with MPN tables to obtain the final count (Pernin et al., 1998; Lares-Villa and Hernández-Peña, 2010; Blodgett, 2024).
-
Pernin et al., 1998Comparative recoveries of Naegleria fowleri amoebae from seeded river water by filtration and centrifugationAppl Environ Microbiol, 1998
-
Lares-Villa and Hernández-Peña, 2010Concentration of Naegleria fowleri in natural waters used for recreational purposes in Sonora, Mexico (November 2007-October 2008)Exp. Parasitol, 2010
-
Blodgett, 2024BAM Appendix 2: Most Probable Number from Serial Dilutions, 2024
Flagellation test
All those strains that showed morphological characteristics suspected of belonging to the genus Naegleria underwent the flagellation test. This test consisted of adding 2 mL of sterile distilled water to each plate and incubating at 37 °C for four hours. Then, we examined each dish by inverted microscopy at two, three, and four hours, and in the presence of flagellated bodies, 0.1 mL of the liquid was transferred to a new plate with NNE to incubate again for 24 h at 45 °C and prepare them for DNA extraction (De Jonckheere, 1977).
-
De Jonckheere, 1977Use of an axenic medium for differentiation between pathogenic and nonpathogenic Naegleria fowleri IsolatesAppl Environ Microbiol, 1977
DNA extraction
Once the pure cultures were obtained and had sufficient growth, we collected the amoebae for DNA extraction using the commercial kit DNeasy® Blood and Tissue Kit (Qiagen), according to the manufacturer’s instructions. Subsequently, we verified its integrity by electrophoresis in 1% agarose gel, stained with ethidium bromide, at 50 V for one hour. Finally, we quantify DNA using a NanoDrop 2000c spectrophotometer (Zysset-Burri et al., 2014).
-
Zysset-Burri et al., 2014Genome-wide identification of pathogenicity factors of the free-living amoeba Naegleria fowleriBMC Genomics, 2014
PCR endpoint and counting of thermophilic Naegleria (TN) and Naegleria fowleri (Nf)
We performed PCR amplification according to the manufacturer’s instructions using the GoTaq Flexi DNA Polymerase Kit (Promega). Primers ITS1 5’-GAACCTGCGTAGGGATCATTT-3’ and ITS2 5’-TTTCTTTTCCTCCCCTTATTA-3’ were used for the identification of the genus Naegleria and for the confirmation of N. fowleri the primers used were FW1 5’-GTGAAAACCTTTTTTCCATTTACA-3‘, RV1 5’-AAATAAAAGATTGACCATTTGAAA-3’, with an expected amplicon length of 410 bp and 310 bp respectively (Zhang et al., 2018). The thermal cycler conditions were as follows: initial denaturation of 94 °C for 3 minutes, followed by 30 cycles of 94 °C for 30 s, 55 °C for 30 s, 72 °C for 30 s, and a final extension at 72 °C for 5 min. Then, we visualized the PCR products in a UV transilluminator at a wavelength of 260 nm after electrophoretic separation in a 2% agarose gel stained with ethidium bromide (Panda et al., 2015). From the thermophilic isolates positive to Naegleria genus and N. fowleri, through PCR test, the number of plates where they grew, of each dilution, was registered to obtain the MPN of thermophilic Naegleria, and the MPN of N. fowleri per liter (Lares-Villa and Hernández-Peña, 2010; Blodgett, 2024).
-
Zhang et al., 2018Identification and molecular typing of Naegleria fowleri from a patient with primary amebic meningoencephalitis in ChinaInt. J. Infect. Dis, 2018
-
Panda et al., 2015Prevalence of Naegleria fowleri in environmental samples from northern part of IndiaPLoS ONE, 2015
-
Lares-Villa and Hernández-Peña, 2010Concentration of Naegleria fowleri in natural waters used for recreational purposes in Sonora, Mexico (November 2007-October 2008)Exp. Parasitol, 2010
-
Blodgett, 2024BAM Appendix 2: Most Probable Number from Serial Dilutions, 2024
Sanger sequencing and bioinformatic analysis to identify genotypes
For strains confirmed as N. fowleri, the ITS1, 5.8S, and ITS2 regions were amplified with the ITS primers. Using the Sanger technique, we purified and sequenced the PCR products in both directions. Subsequently, we edited the sequences, aligned them, and analyzed them with the CLC Genomics Workbench v.20 programs (https://digitalinsights.qiagen.com/) for genotype characterization. We used the representative sequences for each genotype deposited in the GenBank in comparing alignments: AY376149, X96564, X96562, AJ132030, AJ132028, FR875287, X96563 and FR875288 (Zhang et al., 2018).
-
Zhang et al., 2018Identification and molecular typing of Naegleria fowleri from a patient with primary amebic meningoencephalitis in ChinaInt. J. Infect. Dis, 2018
Results and discussion
Sampling sites
Except for Aguacaliente, which is a source of thermal water, the water from the other study sites, La Isleta (where water flows continuously through a canal), Las Palmas (a natural lake), and Laguna del Nainari (an artificial lake), is supplied by water from the Alvaro Obregon dam. It is important to note that in certain areas and times of the year, this water is supplemented with groundwater to meet the specific agricultural needs of the region. While the water in the Yaqui Valley is predominantly used for irrigation, the four sites we investigated are also popular recreational spots for families, offering opportunities for picnics and swimming. However, if N. fowleri is present, the water could be a source of significant health risks. This underscores the gravity of the situation and the importance of our study’s findings for public health.
MPN of thermophilic amoebae (TA)
The data in Table 1 reveal that thermophilic amoebae were present throughout the year. The term ‘thermophilic’ is used for organisms that grow above 40 °C, according to De Jonckheere (2002). In our study, we focused on the growth of N. fowleri, selecting 45 °C as the main isolation temperature to eliminate organisms that cannot grow at this temperature. The data in Table 1 reveal the intriguing complexity of our findings, with thermophilic amoebae being present throughout the year. Still, their figures vary significantly depending on the month and sampling site, ranging from 4 NMP TA/L to 2398 NMP TA/L.
-
De Jonckheere (2002)A century of research on the amoeboflagellate genus NaegleriaActa Protozool, 2002
Table 1
The most probable number of thermophilic amoebae (TA) from La Isleta, Las Palmas, Aguacaliente, and Laguna del Nainari in 2017-2018.
The most probable number of thermophilic amoebae (TA) from La Isleta, Las Palmas, Aguacaliente, and Laguna del Nainari in 2017-2018.
Tabla 1.
Número más probable de amibas termófilas (TA) de La Isleta, Las Palmas, Aguacaliente y Laguna del Náinari en 2017-2018.
Número más probable de amibas termófilas (TA) de La Isleta, Las Palmas, Aguacaliente y Laguna del Náinari en 2017-2018.
| MPN of thermophilic amoebae | ||||
| Sampling | La Isleta | Las Palmas | Aguacaliente | Laguna del Nainari |
| MPN TA/L | MPN TA/L | MPN TA/L | MPN TA/L | |
| 01/05/2017 | 788 (± 458) | 1724 (± 860) | 2398 (± 1209) | 1299 (± 669) |
| 20/06/2017 | 1299 (± 669) | 14* (± 4) | 201 (± 200) | 10* (± 3) |
| 10/07/2017 | 788 (± 458) | 48* (± 12) | 2398 (± 1209) | 8* (± 3) |
| 14/08/2017 | 201 (± 200) | 201 (± 200) | 22* (± 6) | 10* (± 3) |
| 13/09/2017 | <200 | 70* (± 19) | 70* (± 19) | 201 (± 200) |
| 18/10/2017 | 184* (± 54) | 322* (± 89) | 184* (± 54) | 4* (± 2) |
| 15/11/2017 | 452 (± 319) | 322* (± 89) | 201 (± 200) | 108* (± 33) |
| 11/12/2017 | 452 (± 319) | <200 | 1299 (± 669) | 16* (± 5) |
| 15/01/2018 | 108* (± 33) | 10* (± 3) | 184* (± 54) | 10* (± 3) |
| 12/02/2018 | 26* (± 7) | 18* (± 5) | <200 | 4* (± 2) |
| 12/03/2018 | 26* (± 7) | 14* (± 4) | 184* (± 54) | 48* (± 12) |
| 16/04/2018 | 322* (± 89) | 10* (± 3) | 322* (± 89) | 28* (± 7) |
Table 1 presents a clear pattern: Aguacaliente consistently recorded the highest TA concentrations, a trend directly linked to favorable climatic conditions. These conditions, characterized by temperatures above 40 °C, provide an ideal environment for the growth of microorganisms. Among the thermophilic or thermotolerant amoebas reported in Aguacaliente in previous studies are the genera Naegleria, Acanthamoeba, Balamuthia, Vermamoeba, and Stenamoeba (Lares-Jiménez et al., 2018).
-
Lares-Jiménez et al., 2018Potentially pathogenic genera of free-living amoebae coexisting in a thermal springExp. Parasitol, 2018
MPN of thermophilic Naegleria spp.
Table 2 clearly shows the differences between the studied water bodies. In Aguacaliente hot springs, the genus Naegleria was present throughout the sampling year. Compared to the Pearson analysis, we did not observe a correlation between climatic parameters and the presence of Naegleria spp. The number of Naegleria spp. identified by flagellar transformation and confirmed by PCR varied between 4 NMP/L and 788 NMP/L in the different sites sampled, with La Isleta, Las Palmas, and Laguna del Nainari having the lowest densities and frequencies of monthly isolates compared with Aguacaliente. Of the 47 species of Naegleria described worldwide by different methods, including molecular ones, 20 species grow between 40 and 45 or more degrees Celsius, but mainly only seven species grow at 45 °C (Guzmán-Fierros et al., 2008). Of the few genetic studies on Naegleria spp. conducted in different bodies of water in this region, including the four sites of this study, only the presence of thermophilic N. fowleri, N. thiangensis, and N. lovaniensis has been reported. The latter is the most abundant thermophilic Naegleria species (Guzmán-Fierros et al., 2008; Lares-Villa and Hernández-Peña, 2010; Lares-Jiménez et al., 2018).
-
Guzmán-Fierros et al., 2008Identificación de especies de Naegleria en sitios recreativos en Hornos, SonoraRev. Mex. Biodiv, 2008
-
Guzmán-Fierros et al., 2008Identificación de especies de Naegleria en sitios recreativos en Hornos, SonoraRev. Mex. Biodiv, 2008
-
Lares-Villa and Hernández-Peña, 2010Concentration of Naegleria fowleri in natural waters used for recreational purposes in Sonora, Mexico (November 2007-October 2008)Exp. Parasitol, 2010
-
Lares-Jiménez et al., 2018Potentially pathogenic genera of free-living amoebae coexisting in a thermal springExp. Parasitol, 2018
Table 2
The most probable number of thermophilic Naegleria (TN) from La Isleta, Las Palmas, Aguacaliente, and Laguna del Nainari in 2017-2018.
The most probable number of thermophilic Naegleria (TN) from La Isleta, Las Palmas, Aguacaliente, and Laguna del Nainari in 2017-2018.
Tabla 2.
Número más probable de Naegleria termófilas (TN) de La Isleta, Las Palmas, Aguacaliente y Laguna del Náinari en 2017-2018.
Número más probable de Naegleria termófilas (TN) de La Isleta, Las Palmas, Aguacaliente y Laguna del Náinari en 2017-2018.
| MPN of thermophilic Naegleria | ||||
| Sampling | La Isleta | Las Palmas | Aguacaliente | Laguna del Nainari |
| MPN TN/L | MPN TN/L | MPN TN/L | MPN TN/L | |
| 01/05/2017 | - | 4* (± 2) | 322* (± 89) | - |
| 20/06/2017 | - | - | 201 (± 200) | - |
| 10/07/2017 | 22* (± 6) | 4* (± 2) | 788 (± 458) | - |
| 14/08/2017 | 4* (± 2) | 70* (± 19) | 10* (± 3) | - |
| 13/09/2017 | 4* (± 2) | 16* (± 5) | 34* (± 9) | 70* (± 19) |
| 18/10/2017 | - | 10* (± 3) | 184* (± 54) | - |
| 15/11/2017 | 4* (± 2) | 4* (± 2) | 34* (± 9) | 4* (± 2) |
| 11/12/2017 | - | 16* (± 5) | 201 (± 200) | - |
| 15/01/2018 | - | - | 184* (± 54) | - |
| 12/02/2018 | - | - | 8* (± 3) | - |
| 12/03/2018 | - | - | 34* (± 9) | - |
| 16/04/2018 | - | - | 22* (± 6) | - |
MPN of Naegleria fowleri
The results shown in Table 3 confirm the presence of N. fowleri in all four study sites and establish a connection with the findings of Lares-Villa and Hernández Peña (2010). The levels of N. fowleri varied depending on the month and location of sampling, ranging between 4 NMP Nf/L and 70 NMP Nf/L. Notably, N. fowleri was detected during August, September, and October, which aligns with the months reported in the study mentioned above and with data ranging from 4 to 18 NMP Nf/L. This correlation enhances the reliability of our research and contributes to the existing knowledge about N. fowleri in natural water bodies.
-
Lares-Villa and Hernández Peña (2010)Concentration of Naegleria fowleri in natural waters used for recreational purposes in Sonora, Mexico (November 2007-October 2008)Exp. Parasitol, 2010
Table 3
The most probable number of N. fowleri (Nf) from La Isleta, Las Palmas, Aguacaliente, and Laguna del Nainari in 2017-2018.
The most probable number of N. fowleri (Nf) from La Isleta, Las Palmas, Aguacaliente, and Laguna del Nainari in 2017-2018.
Tabla 3.
Número más probable de N. fowleri (Nf) de La Isleta, Las Palmas, Aguacaliente y Laguna del Náinari en 2017-2018.
Número más probable de N. fowleri (Nf) de La Isleta, Las Palmas, Aguacaliente y Laguna del Náinari en 2017-2018.
| MPN of Naegleria fowleri | ||||
| Sampling | La Isleta | Las Palmas | Aguacaliente | Laguna del Nainari |
| MPN Nf/L | MPN Nf/L | MPN Nf/L | MPN Nf/L | |
| 01/05/2017 | - | - | - | - |
| 20/06/2017 | - | - | - | - |
| 10/07/2017 | - | - | - | - |
| 14/08/2017 | - | 22* (± 6) | - | - |
| 13/09/2017 | 4* (± 2) | 16* (± 5) | - | 70* (± 19) |
| 18/10/2017 | - | - | 18* (± 5) | - |
| 15/11/2017 | - | - | - | - |
| 11/12/2017 | - | - | - | - |
| 15/01/2018 | - | - | - | - |
| 12/02/2018 | - | - | - | - |
| 12/03/2018 | - | - | - | - |
| 16/04/2018 | - | - | - | - |
This report marks the first documented presence of N. fowleri in Laguna del Nainari and Aguacaliente. Our findings suggest that the lagoon’s connection to La Isleta, which supplies its water, may have facilitated the amoeba’s presence in September. Similarly, the onset of the rainy season in October could have introduced the pathogenic amoeba into the thermal water source in Aguacaliente, challenging the notion that temperatures above 40 °C alone are responsible (Stahl and Olson, 2021).
-
Stahl and Olson, 2021Environmental abiotic and biotic factors affecting the distribution and abundance of Naegleria fowleriFEMS Microbiology Ecology, 2021
Table 4 presents the results of identifying free-living amoebae, which were isolated and grown at the precise temperature of 45 °C at the four specific sampling points under study. Our research involved a meticulously designed and executed process of isolation and identification of a total of 655 thermophilic AVL strains. Among these, 260 showed flagellated bodies. Through PCR analysis, we identified 238 as Naegleria spp. and 22 as N. fowleri, providing a comprehensive and reliable understanding of the presence of these strains in the studied areas. It was found that 91.5% of amoeboid organisms with flagella tested positive in the endpoint PCR for Naegleria species. This suggests that another genus with flagellated organisms in its life cycle was identified in the study, potentially including different amoebae belonging to the Heterolobosea class (Page, 1988; Robinson et al., 1989). The isolation of N. fowleri, representing approximately 10% of the isolates, is a significant finding. However, when we compare it with the total thermophilic AVL, the number of N. fowleri would barely represent approximately 3.4%. This means that N. fowleri, the only strain of Naegleria pathogenic for humans reported so far, is found in meager proportions in natural environments.
-
Page, 1988A new key to freshwater and soil gymnamoebae: with instructions for culture, 1988
-
Robinson et al., 1989A temporary flagellate (mastigote) stage in the vahlkampfiid amoeba Willaertia magna and its possible evolutionary significanceBioSystems, 1989
Table 4
Free-living amoebae isolated and grown at 45 °C, from La Isleta, Las Palmas, Aguacaliente, and Laguna del Nainari, Sonora, from May 2017 to April 2018.
Free-living amoebae isolated and grown at 45 °C, from La Isleta, Las Palmas, Aguacaliente, and Laguna del Nainari, Sonora, from May 2017 to April 2018.
Tabla 4.
Amibas de vida libre aisladas y cultivadas a 45 °C, de La Isleta, Las Palmas, Aguacaliente y Laguna del Náinari, Sonora, de mayo de 2017 a abril de 2018..
Amibas de vida libre aisladas y cultivadas a 45 °C, de La Isleta, Las Palmas, Aguacaliente y Laguna del Náinari, Sonora, de mayo de 2017 a abril de 2018..
| Free living amoebae | La Isleta | Las Palmas | Aguacaliente | Laguna del Náinari | Total |
| Thermophilic amoebae | 273 | 104 | 225 | 53 | 655 |
| Positive flagellation test | 16 | 36 | 198 | 10 | 260 |
| Thermophilic Naegleria spp. | 8 | 29 | 192 | 9 | 238 |
| Naegleria fowleri | 1 | 10 | 4 | 7 | 22 |
Regarding the strains positive for N. fowleri, they represent only 9.2% of Naegleria spp. It has been found that there is a more significant proliferation of other species, such as N. lovaniensis, which has been previously demonstrated (Guzmán-Fierros et al., 2008; Lares-Jiménez et al., 2018), in addition to the fact that the pathogen represents only 3.4% of the total, results that agree with those obtained by Lares-Villa and Hernández-Peña (2010), where N. fowleri only represented 1.8% of the total isolated thermophilic amoebas. This reiteration of the agreement of our results with previous studies should instill confidence in the scientific community. Although none of the sites reported levels of N. fowleri greater than 100 MPN L-1, a value established by Cabanes et al. (2001), where they mention that values higher than this number mean a risk of infection for the population, or if the standard applied by the Australian government were taken as a reference, which implies two thermophilic Naegleria per liter (De Jonckheere, 2014), it is must prevent the population from accessing all the sampling locations studied, particularly during August, September and October.
-
Guzmán-Fierros et al., 2008Identificación de especies de Naegleria en sitios recreativos en Hornos, SonoraRev. Mex. Biodiv, 2008
-
Lares-Jiménez et al., 2018Potentially pathogenic genera of free-living amoebae coexisting in a thermal springExp. Parasitol, 2018
-
Lares-Villa and Hernández-Peña (2010)Concentration of Naegleria fowleri in natural waters used for recreational purposes in Sonora, Mexico (November 2007-October 2008)Exp. Parasitol, 2010
-
Cabanes et al. (2001)Assessing the risk of primary amoebic meningoencephalitis from swimming in the presence of environmental Naegleria fowleriAppl. Environ. Microbiol, 2001
-
De Jonckheere, 2014What do we know by now about the genus Naegleria? ExpParasitol, 2014
Genotyping of Naegleria fowleri
Once the nucleotide sequences were obtained, assembled, edited, aligned, and analyzed with the CLC Genomics program for genotype characterization, we found that the seven N. fowleri strains belong to type 2, which have a length of 42 bp in their ITS1 (Figure 1) and nucleotide T at position 31 in its 5.8S rDNA (Figure 2). As described by De Jonckheere (2011), there are eight genotypes distributed around the world that have been characterized based on the number of repetitions in the ITS1 region, which can vary in size depending on the characteristics of each genotype, having a range of 42-142 bp and a C / T transition at position 31 in the 5.8S region. The ITS2 region is identical among all N. fowleri strains and, therefore, is not analyzed in these characterizations (De Jonckheere, 2004). Based on this and the analysis carried out in this work, it was observed that the seven N. fowleri strains belong to type 2, which have a length of 42 bp in their ITS1 (Figure 1) and nucleotide T at position 31 in its 5.8S rDNA (Figure 2). The discovery of type 2 in the Yaqui Valley aligns with the findings of De Jonckheere (2011), who reported that type 1, 2, and 3 genotypes have been found in America. Additionally, Vargas-Zepeda et al. (2005) conducted a molecular analysis of N. fowleri isolated from a CSF sample in Sonora and identified type 2. This suggests the need to expand the analysis in Mexico to confirm the prevalence of genotype 2 in both the region and the country. Genotype 2 has been found in environmental samples as well as in clinical cases (Pelandakis et al., 2000; Zhou et al., 2003; Cogo et al., 2004; Nicolas et al., 2010). In Taiwan, the first case of PAM was reported in November 2011, and subsequent sampling of recreational hot springs visited by the patient revealed the presence of N. fowleri type 2, which matched the type found in the patient. This molecular characterization method was used as an epidemiological tracing tool (Tung et al., 2013). More recently, Zhang et al. (2018) conducted a molecular typing study of this amoeba from a patient’s sample in the Zhejiang province of China and once again found type 2.
-
De Jonckheere (2011)Origin and evolution of the worldwide distributed pathogenic amoeboflagellate Naegleria fowleriInfection, Genetics and Evolution, 2011
-
De Jonckheere, 2004Molecular definition and the ubiquity of species in the genus NaegleriaProtist, 2004
-
De Jonckheere (2011)Molecular definition and the ubiquity of species in the genus NaegleriaProtist, 2004
-
Vargas-Zepeda et al. (2005)Successful treatment of Naegleria fowleri meningoencephalitis by using intravenous amphotericin B, fluconazole and rifampicin. Case reportArch. Med. Res, 2005
-
Pelandakis et al., 2000Analysis of the 5.8S rRNA gene and the internal transcribed spacers in Naegleria spp. and in N. fowleriJ. Eukar. Microbiol, 2000
-
Zhou et al., 2003Genetic variations in the internal transcribed spacer and mitochondrial small subunit rRNA gene of Naegleria sppJ. Eukaryot. Microbiol, 2003
-
Cogo et al., 2004Fatal Naegleria fowleri meningoencephalitis. ItalyEmerg. Infect. Dis, 2004
-
Nicolas et al., 2010Diagnostic moléculaire d’une méningoencephalite amibienne primitive à l’occasion d’un cas fatal en GuadeloupeBull. Soc. Pat. Exo, 2010
-
Tung et al., 2013Identification and significance of Naegleria fowleri isolated from the hot spring which related to the first primary amebic meningoencephalitis (PAM) patient in TaiwanInt. J. Parasitol, 2013
-
Zhang et al. (2018)Identification and molecular typing of Naegleria fowleri from a patient with primary amebic meningoencephalitis in ChinaInt. J. Infect. Dis, 2018
Thumbnail
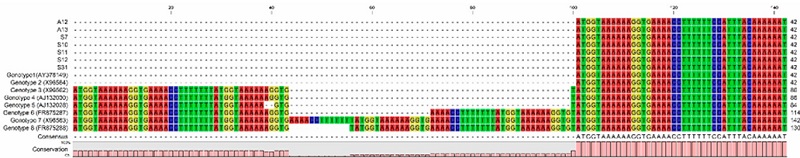
Figure 1
Alignment of the ITS1 regions of Naegleria fowleri. DNA sequence alignments were performed using QIAGEN CLC Genomics Workbench 21.0. The length of the ITS1 region and the base at position 31 of the 5.8S rRNA sequences, were compared using genotypes 1-8 deposited in the National Center for Biotechnology Information (NCBI) database.
Alignment of the ITS1 regions of Naegleria fowleri. DNA sequence alignments were performed using QIAGEN CLC Genomics Workbench 21.0. The length of the ITS1 region and the base at position 31 of the 5.8S rRNA sequences, were compared using genotypes 1-8 deposited in the National Center for Biotechnology Information (NCBI) database.
Thumbnail
Figura 1.
Alineación de las regiones ITS1 de Naegleria fowleri. Las alineaciones de secuencias de ADN se realizaron utilizando QIAGEN CLC Genomics Workbench 21.0. La longitud de la región ITS1 y la base en la posición 31 de las secuencias de ARNr 5.8S se compararon utilizando los genotipos 1 a 8 depositados en la base de datos del Centro Nacional de Información Biotecnológica (NCBI).
Alineación de las regiones ITS1 de Naegleria fowleri. Las alineaciones de secuencias de ADN se realizaron utilizando QIAGEN CLC Genomics Workbench 21.0. La longitud de la región ITS1 y la base en la posición 31 de las secuencias de ARNr 5.8S se compararon utilizando los genotipos 1 a 8 depositados en la base de datos del Centro Nacional de Información Biotecnológica (NCBI).
Thumbnail
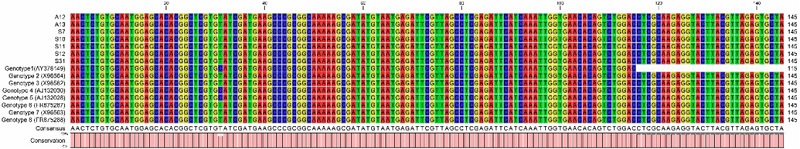
Figure 2
Alignment of the 5.8S region (145 bp) showing the C / T transition at position 31.
Alignment of the 5.8S region (145 bp) showing the C / T transition at position 31.
Thumbnail
Figura 2.
Alineación de la región 5.8S (145 pb) que muestra la transición C/T en la posición 31.
Alineación de la región 5.8S (145 pb) que muestra la transición C/T en la posición 31.
Water temperature, environmental temperature, pH, and dissolved oxygen at the sampling sites
Figures 3-6 show the mean and standard deviation of the monthly measured parameters (water temperature, ambient temperature, pH, and dissolved oxygen) grouped by seasons of the year and sampling sites. In general, samples collected during the winter (December to February) had the lowest average water and ambient temperatures, while the highest temperatures were obtained in the summer (June to August), showing significant differences in both seasons. Therefore, values with the same letter are statistically equal (p≤ 0.05) (Figures 3 and 4).
Thumbnail
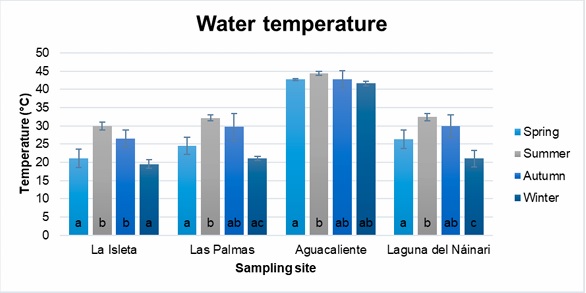
Figure 3
Annual behavior of water temperature at sampling sites.
Annual behavior of water temperature at sampling sites.
Thumbnail
Figura 3.
Comportamiento anual de la temperatura del agua en los sitios de muestreo.
Comportamiento anual de la temperatura del agua en los sitios de muestreo.
Thumbnail
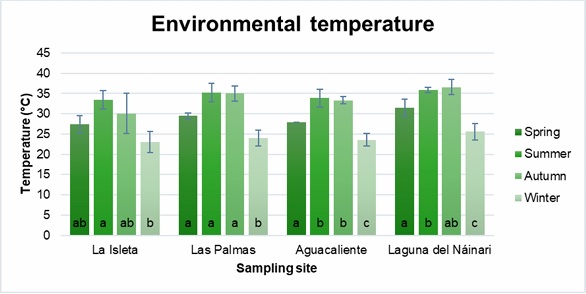
Figure 4
Annual behavior of environmental temperature at sampling sites.
Annual behavior of environmental temperature at sampling sites.
Thumbnail
Figura 4.
Comportamiento anual de la temperatura ambiental en los sitios de muestreo.
Comportamiento anual de la temperatura ambiental en los sitios de muestreo.
Thumbnail
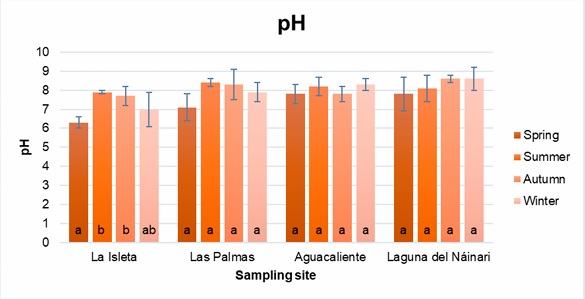
Figure 5
Annual water pH behavior at sampling sites.
Annual water pH behavior at sampling sites.
Thumbnail
Figura 5.
Comportamiento anual del pH del agua en los sitios de muestreo.
Comportamiento anual del pH del agua en los sitios de muestreo.
Thumbnail
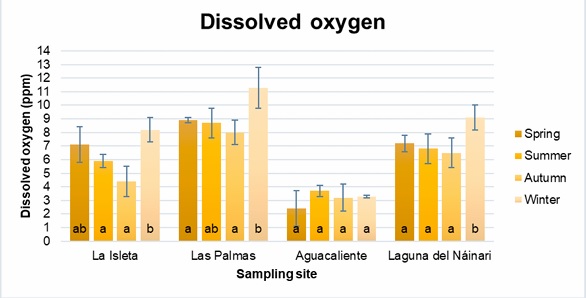
Figure 6
Annual behavior of dissolved oxygen at sampling sites.
Annual behavior of dissolved oxygen at sampling sites.
Thumbnail
Figura 6.
Comportamiento anual del oxígeno disuelto en los sitios de muestreo.
Comportamiento anual del oxígeno disuelto en los sitios de muestreo.
In the case of water temperatures, the effect of the different seasons of the year is observed in the three reservoirs of natural water coming from the dam, and this is not the case in Aguacaliente, which is a source of natural thermal water (Figure 3). At this site, the water temperature was above 40 °C all year round. Although slight variations in environmental temperature are observed in the four sampling sites, we can practically observe two climates, summer and autumn, with the highest temperatures, and winter and spring, with the lowest temperatures (Figure 4). As seen in Table 1, the thermal body of Aguacaliente presented the highest concentrations of thermophilic amoebas throughout the year, a situation attributed to the primary favorable climatic condition for the growth of this type of microorganisms, which is water temperatures above 40 °C throughout the year. When performing the Pearson analysis, we did not observe a correlation between the behaviors of the number of thermophilic amoebae and the presented climatic conditions.
The pH averages obtained were similar at the four sampling sites throughout the year, except for La Isleta, which showed a statistically significant decrease in the spring with a pH R-value of -0.9020. This decrease in pH was particular because the waters of La Isleta, Las Palmas, and the Nainari Lagoon are interconnected, and the pH of the water did not vary significantly. The decrease in pH was probably due to the discharge of some contaminant into the water flow from upstream on the day of sampling. Regarding dissolved oxygen, except in Aguacaliente, the other sites showed significant differences, mainly between the autumn and winter seasons. In the Pearson analysis carried out to observe correlations between the variables, it was shown that there is a statistically significant relationship between the water temperature and the ambient temperature for La Isleta, Las Palmas and Laguna del Náinari, with an R value of 0.9563, 0.9711 and 0.9647, respectively. In addition, in the case of the Laguna del Náinari, a significant relationship was found between dissolved oxygen and water temperature, as well as statistically significant for environmental temperature, with an R-value of -0.9170 and -0.9717, respectively. On the other hand, for Las Palmas, only a meaningful relationship was presented, with an R-value of -0.9226 between dissolved oxygen and environmental temperature.
According to Marín-Galvín (2019), dissolved oxygen is a very relevant gas in the dynamics of water, where its solubility is a function of several factors: temperature, pressure, coefficient of solubility, vapor pressure, salinity, and physicochemical composition of water. In this way, we can see that the amount of dissolved oxygen varies inversely proportional to temperature, results that have already been observed in various studies. This is associated with the fact that at higher temperatures, aquatic organisms require more oxygen, which tends to decrease (Rosenfeld, 2017). Although the effect of some abiotic factors remains inconclusive for the presence of N. fowleri in natural surface waters, including the increase in water temperature due to climate change (Navarro-Estupiñan et al., 2018), it is necessary to continue studying not only abiotic factors, but also biotic and nutritional factors to know whether or not these determine the increase or control of the population of this pathogenic amoeba (Stahl and Olson, 2021).
-
Marín-Galvín (2019)Fisicoquímica y microbiología de los medios acuáticos tratamiento y control de calidad de aguas, 2019
-
Rosenfeld, 2017Developing flow-ecology relationships: implications of nonlinear biological responses for water managementFreshwater Biol, 2017
-
Navarro-Estupiñan et al., 2018Observed trends and future projections of extreme heat events in Sonora, MexicoInt J Climatol, 2018
-
Stahl and Olson, 2021Environmental abiotic and biotic factors affecting the distribution and abundance of Naegleria fowleriFEMS Microbiology Ecology, 2021
Conclusions
Having determined the presence of this pathogen in the four sites studied demonstrates its wide ecological diversity because each of them shows different characteristics, and they also have great distances between them, so it would be interesting to focus future studies on the transport routes that can influence the movement of this pathogen, together with implementing greater parameters that allow a better understanding of the behavior of the species. Likewise, it is suggested that the four sites continue to be monitored because, with the current climate change and environmental contamination, the population densities of this pathogen may increase, posing a great risk of infection for those who visit these sites. Specifically, constant monitoring of the Laguna del Náinari is suggested since it was the site that showed the highest concentration and is located within the city, in addition to its supply being made through the low channel to which the population has access and is used to irrigate crops, generating contamination risks in nearby towns.
Conflicts of interest
The authors declare no conflict of interest.
Acknowledgments
Thanks to Bernardo Baldenea-Vega, Jesús-Andrea Campas-Aguirre, Rebeca Estefanya Payán-Sánchez and Leobardo-Daniel González-Zúñiga for their technical support. This work was funded by Coordinación de gestión y apoyo a cuerpos académicos del Instituto Tecnológico de Sonora (Project PROFAPI 2023_CA_019 RCH).
References
- Blodgett, R. 2024. BAM Appendix 2: Most Probable Number from Serial Dilutions | FDA. Available from: Available from: https://www.fda.gov/food/laboratory-methods-food/bam-appendix-2-most-probable-number-serial-dilutions [Accessed 11/03/24]. Links
- Cabanes, P.A., Wallet, F., Pringuez, E. and Pernin, P. 2001. Assessing the risk of primary amoebic meningoencephalitis from swimming in the presence of environmental Naegleria fowleri. Appl. Environ. Microbiol. 67(7), 2927-2931. DOI:10.1128/AEM.67.7.2927-2931.2001 Links
- Centers for Disease Control and Prevention. 2022. Naegleria fowleri: primary amebic meningoencephalitis (PAM)-amebic encephalitis. https://www.cdc.gov/parasites/naegleria/infection-sources.html [Accessed 13/03/ 2024]. Links
- Cogo, P.E., Scaglia, M., Gatti, S., Rossetti, F., Alaggio, R., Laverda, A.M., Zhou, L., Xiao, L. and Visvesvara, G. 2004. Fatal Naegleria fowleri meningoencephalitis. Italy. Emerg. Infect. Dis. 10, 1835-1837. DOI: 10.3201/eid1010.040273 Links
- Cope, J.R., Kahler, A.M., Causey, J., Williams, J.G., Kihlken, J., Benjamin, C., Ames, A.P., Forsman, J., Zhu, Y., Yoder, J.S., Seidel, C.J. and Hill, V.R. 2019. Response and remediation actions following the detection of Naegleria fowleri in two treated drinking water distribution systems, Louisiana, 2013-2014. J Water Health. 17(5):777-787. DOI: 10.2166/wh.2019.239 Links
- De Jonckheere, J.F. 2014. What do we know by now about the genus Naegleria? Exp. Parasitol. 145, S2-S9. DOI: 10.1016/j.exppara.2014.07.011 Links
- De Jonckheere, J.F. 2011. Origin and evolution of the worldwide distributed pathogenic amoeboflagellate Naegleria fowleri. Infection, Genetics and Evolution. 11(7), 1520-1528. DOI: 10.1016/j.meegid.2011.07.023 Links
- De Jonckheere, J.F. 2004. Molecular definition and the ubiquity of species in the genus Naegleria. Protist. 155, 89-103. DOI: 10.1078/1434461000167 Links
- De Jonckheere, J.F. 2002. A century of research on the amoeboflagellate genus Naegleria. Acta Protozool. 41, 309-342. Links
- De Jonckheere, J.F. 1977. Use of an axenic medium for differentiation between pathogenic and nonpathogenic Naegleria fowleri Isolates. Appl Environ Microbiol. 33(4): 751-757. DOI: https://doi.org/10.1128/aem.33.4.751-757.1977 Links
- Di Filippo, M.M., Novelletto, A., Di Cave, D. and Berrilli, F. 2017. Identification and phylogenetic position of Naegleria spp. from geothermal springs in Italy. Exp. Parasitol. DOI:10.1016/j.exppara.2017.08.008 Links
- Gompf, S.G. and Garcia, C. 2019. Lethal encounters: The evolving spectrum of amoebic meningoencephalitis. IDCases. 15. https://doi.org/10.1016/j.idcr.2019.e00524 Links
- Guzmán-Fierros, E., De Jonckheere, J.F. and Lares-Villa, F. 2008. Identificación de especies de Naegleria en sitios recreativos en Hornos, Sonora. Rev. Mex. Biodiv. 79, 1-5. Links
- INAFED. http://www.inafed.gob.mx/work/enciclopedia/EMM26sonora/municipios/26018a.html [Accessed 3/06/2020]. Links
- John, D.T. and Howard, M.J. 1995. Seasonal distribution of pathogenic free-living amebae in Oklahoma waters. Parasitol Res. 81(3), 193-201. DOI:10.1007/BF00937109 Links
- Kao, P.M., Tung, M.C., Hsu, B.M., Hsueh, C.J., Chiu, Y.C., Chen, N.H., Shen, S.M. and Huang, Y.L. 2012. Occurrence and distribution of Naegleria species from thermal spring environments in Taiwan. Letters in Applied Microbiology 56, 1-7 Links
- Kyle, D.E. and Noblet, G.P. 1986. Seasonal distribution of thermotolerant free‐living amoebae. I. Willard’s Pond. J. Protozool. 33(3), 422-434. https://doi.org/10.1111/j.1550-7408.1986.tb05634.x Links
- Lares-Jiménez, L.F., Borquez-Román, M.A., Lares-García, Ch., Otero-Ruiz, A., Gonzalez-Galaviz, J.R., Ibarra-Gámez, J.C. and Lares-Villa, F. 2018. Potentially pathogenic genera of free-living amoebae coexisting in a thermal spring. Exp. Parasitol. 195, 54-58. https://doi.org/10.1016/j.exppara.2018.10.006 Links
- Lares-Villa, F., Guerrero-Dumas, A.M. and López-Osorio, A. 1997. Estudio protozoológico (subphylum Sarcodina clase Lobosea), en ambientes acuáticos de la región del Valle del Yaqui, Sonora, México. ITSON-DIEP. 2:47-55. Links
- Lares-Villa, F. and Hernández-Peña, C. 2010. Concentration of Naegleria fowleri in natural waters used for recreational purposes in Sonora, Mexico (November 2007-October 2008). Exp. Parasitol. 126: 33-36. https://doi.org/10.1016/j.exppara.2010.04.011 Links
- Maclean, R.C., Richardson, D.J., LePardo, R. and Marciano-Cabral, F. 2004. The identification of Naegleria fowleri from water and soil samples by nested PCR. Parasitol. Res. 93(3), 211-217. DOI: 10.1007/s00436-004-1104-x Links
- Marín-Galvín, R. 2019. Fisicoquímica y microbiología de los medios acuáticos tratamiento y control de calidad de aguas. 2ª ed. Editorial Díaz de Santos. Universidad Politécnica de Cartagena. España, p. 440. Links
- Miller, H.C., Morgan, M.J., Walsh, T., Wylie, J.T., Kaksonen, A.H. and Puzon, G.J. 2018. Preferential feeding in Naegleria fowleri; intracellular bacteria isolated from amoebae in operational drinking water distribution systems. Water Res. 15(141):126-134. doi: 10.1016/j.watres.2018.05.004 Links
- Miller, H.C., Morgan, M.J., Wylie, J.T., Kaksonen, A.H., Sutton, D., Braun, K. and Puzon, G.J. 2017. Elimination of Naegleria fowleri from bulk water and biofilm in an operational drinking water distribution system. Water Res 110:15-26. DOI: 10.1016/j.watres.2016.11.061 Links
- Miller, H.C., Wylie, J., Dejean, G., Kaksonen, A.H., Sutton, D., Braun, K. and Puzon, G.J. 2015. Reduced efficiency of chlorine disinfection of Naegleria fowleri in a drinking water distribution biofilm. Environ Sci Technol 49:11125-11131, DOI: 10.1021/acs.est.5b02947 Links
- Morgan, M.J., Halstrom, S., Wylie, J.T., Walsh, T., Kaksonen, A.H., Sutton, D., Braun, K. and Puzon, G.J. 2016. Characterization of a drinking water distribution pipeline terminally colonized by Naegleria fowleri. Environ Sci Technol 50, 2890-2898. DOI: 10.1021/acs.est.5b05657 Links
- Moussa, M., Marcelino, I., Richard, V., Guerlotté, J. and Talarmin A. 2020. An optimized most probable number (MPN) method to assess the number of thermophilic free-living amoebae (FLA) in water samples. Pathogens 24: 9(5):409. DOI: 10.3390/pathogens9050409 Links
- Navarro-Estupiñan, J., Robles-Morua, A., Vivoni, E.R., Espíndola-Zepeda, J., Montoya, J.A. and Verduzco, V.S. 2018. Observed trends and future projections of extreme heat events in Sonora, Mexico. Int J Climatol. 1-14. https://doi.org/10.1002/joc.5719 Links
- Nicolas, M., De Jonckheere, J.F., Pernin, P., Bataille, H., Le Bris, V. and Hermann, S.C. 2010. Diagnostic moléculaire d’une méningoencephalite amibienne primitive à l’occasion d’un cas fatal en Guadeloupe. Bull. Soc. Pat. Exo. 103, 14-18. Links
- Page, F.C. 1988. A new key to freshwater and soil gymnamoebae: with instructions for culture. Freshwater Biological Association. Ambleside, Cumbria, UK. p. 122. Links
- Panda, A., Khalil, S., Mirdha, B.R., Singh, Y. and Kaushik, S. 2015. Prevalence of Naegleria fowleri in environmental samples from northern part of India. PLoS ONE. 10(10): e0137736. DOI: 10.1371/journal.pone.0137736 Links
- Pélandakis, M., Serre, S. and Pernin, P. 2000. Analysis of the 5.8S rRNA gene and the internal transcribed spacers in Naegleria spp. and in N. fowleri. J. Eukar. Microbiol. 47, 116-121. https://doi.org/10.1111/j.1550-7408.2000.tb00020.x Links
- Pernin, P., Pelandakis, M., Rouby, Y., Faure, A. and Siclet, F. 1998.Comparative recoveries of Naegleria fowleri amoebae from seeded river water by filtration and centrifugation. Appl Environ Microbiol 64: 955-959. DOI: 10.1128/AEM.64.3.955-959.1998 Links
- Puzon, G.J., Wylie, J.T., Walsh, T., Braun, K. and Morgan, M.J. 2017. Comparison of biofilm ecology supporting growth of individual Naegleria species in a drinking water distribution system. FEMS Microbiol. Ecol. 93, 1-8. DOI: 10.1093/femsec/fix017 Links
- Puzon, G.J., Lancaster, J.A., Wylie, J.T. and Plumb, J.J. 2009. Rapid detection of Naegleria fowleri in water distribution pipeline biofilms and drinking water samples. Environ Sci Technol 43: 6691-6696. DOI: 10.1021/es900432m Links
- Robinson, B S., Christy, P.E. and De Jonckheere, J.F. 1989. A temporary flagellate (mastigote) stage in the vahlkampfiid amoeba Willaertia magna and its possible evolutionary significance. BioSystems. 23(1), 75-86. DOI:10.1016/0303-2647(89)90010-5 Links
- Rosenfeld, J.S. 2017. Developing flow-ecology relationships: implications of nonlinear biological responses for water management. Freshwater Biol. 62(8), 1305-1324. https://doi.org/10.1111/fwb.12948 Links
- Ruszkiewicz, J.A., Tinkov, A.A., Skalny, A.V., Siokas, V., Dardiotis, E., Tsatsakis, A., Bowman, A.B., da Rocha, J.B.T. and Aschner, M. 2019. Brain diseases in changing climate. Environ. Res. 177,108637. https://doi.org/10.1016/j.envres.2019.108637 Links
- Siddiqui, R., Ali, I.K.M., Cope, J.R. and Khan, N.A. 2016. Biology and pathogenesis of Naegleria fowleri. Acta tropica. 164, 375-394. DOI:10.1016/j.actatropica.2016.09.009 Links
- Sifuentes, L.Y., Choate, B.L., Gerba, C.P. and Bright, K.R. 2014. The occurrence of Naegleria fowleri in recreational waters in Arizona. Journal of Environmental Science and Health, Part A: Toxic/Hazardous Substances and Environmental Engineering 49:11, 1322-1330. http://dx.doi.org/10.1080/10934529.2014.910342 Links
- Stahl, L.M. and Olson, J.B. 2021. Environmental abiotic and biotic factors affecting the distribution and abundance of Naegleria fowleri. FEMS Microbiology Ecology, 97:1-13. https://doi.org/10.1093/femsec/fiaa238 Links
- Streby, A., Mull, B.J., Levy, K. and Hill, V.R. 2015. Comparison of real-time PCR methods for the detection of Naegleria fowleri in surface water and sediment. Parasitol Res 114:1739-1746. DOI: 10.1007/s00436-015-4359-5 Links
- Sykora, J.L., Keleti, G. and Martinez, A.J. 1983. Occurrence and pathogenicity of Naegleria fowleri in artificially heated waters. Appl. Environ. Microbiol. 45(3), 974-979 Links
- Tung, M.C., Hsu, B.M., Tao, C.W., Lin, W.C., Tsai, H.F., Ji, D.D., Shen, S.M., Chen, J.S., Shih, F.C. and Huang Y.L. 2013. Identification and significance of Naegleria fowleri isolated from the hot spring which related to the first primary amebic meningoencephalitis (PAM) patient in Taiwan. Int. J. Parasitol. 43(9), 691-696. DOI:10.1016/j.ijpara.2013.01.012 Links
- Vargas-Zepeda, J., Gómez-Alcalá, A., Vázquez-Morales, J.A., Licea-Amaya, L., De Jonckheere, J.F. and Lares-Villa, F. 2005. Successful treatment of Naegleria fowleri meningoencephalitis by using intravenous amphotericin B, fluconazole and rifampicin. Case report. Arch. Med. Res. 36, 83-86. Links
- Weik, R.R. and John, D. T. 1977. Agitated mass cultivation of Naegleria fowleri. J. Parasitol. 868-871. Links
- Xue, J., Lamar, F.G., Zhang, B., Lin, S., Lamori, J.G. and Sherchan, S.P. 2018. Quantitative assessment of Naegleria fowleri and fecal indicator bacteria in brackish water of Lake Pontchartrain, Louisiana. Sci. Total Environ. 622: 8-16. DOI:10.1016/j.scitotenv.2017.11.308 Links
- Yoder, J.S., Straif-Bourgeois, S., Roy, S.L., Moore, T.A., Visvesvara, G.S., Ratard, R.C., Hill, V.R., Wilson, J.D., Linscott, A.J., Crager, R., Kozak, N.A., Sriram, R., Narayanan, J., Mull, B., Kahler, A.M., Schneeberger, C., da Silva, A., Poudel, M., Baumgarten, K.L., Xiao, L. and Beach, M.J. 2012. Primary amebic meningoencephalitis deaths associated with sinus irrigation using contaminated tap water. Clin. Infect. Dis. 55(9): e79-e85. https://doi.org/10.1093/cid/cis626 Links
- Zhang, L.L., Wu, M., Hu, B.C., Chen, H.L., Pan, J.R., Ruan, W. and Yao, L.N. 2018. Identification and molecular typing of Naegleria fowleri from a patient with primary amebic meningoencephalitis in China. Int. J. Infect. Dis. 72: 28-33. DOI:10.1016/j.ijid.2018.05.001 Links
- Zhou, L., Sriram, R., Visvesvara, G.S. and Xiao, L. 2003. Genetic variations in the internal transcribed spacer and mitochondrial small subunit rRNA gene of Naegleria spp. J. Eukaryot. Microbiol. 50, 522-526. DOI:10.1111/j.1550-7408.2003.tb00617.x Links
- Zysset-Burri, D.C., Müller, N., Beuret, C., Heller, M., Schürch, N., Gottstein, B. and Wittwer, M. 2014. Genome-wide identification of pathogenicity factors of the free-living amoeba Naegleria fowleri. BMC Genomics. 15: 496. DOI: 10.1186/1471-2164-15-496 Links