











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. Introduction
Trace fossils are regarded as autochthonous indicators of palaeoenvironmental conditions and have been widely used in sedimentary environment and facies analysis (Hertweck, 1972; Bromley, 1990, Wetzel et al., 2009). They provide crucial palaeoecological data that facilitates more accurate characterization of a depositional setting, particularly in deposits where body fossils are scarce or absent (e.g.Buatois and Mángano, 2011). Many environmental parameters such as nutrient supply, salinity, sedimentation rates, substrate consistency, temperature, hydrodynamic regime, and oxygenation all exert control over the type of behavior (ethology) of fauna in a given environment (e.g.Pemberton et al., 1992a, 1992b; Bayet-Goll et al., 2015) and define, therefore, the specific ecological niches utilized by organisms. However, some limiting factors (e.g. oxygen and food content) condition the colonization and establishment of organisms (Pickerill and Brenchley, 1991; Benton and Harper, 2009). Hence, many factors are not documented in the sedimentary record and can only be indirectly inferred from the fossil record (Benton and Harper, 2009).
The sensitivity of many animals to environmental conditions means that the produced trace fossils and their ichnoassemblages can be correspondingly more sensitive environmental indicators than inorganic sedimentary structures (Crimes, 1975). Moreover, according to Allington-Jones et al. (2010), these ichnoassemblages tend to change progressively with depth, thereby providing a foundation for relative bathymetry. Consequently, the composition of ichnoassemblages and the distribution of trace fossils can provide a wealth of information about environmental factors, depositional environments, and palaeoecology (e.g.Gingras et al., 2011; Bann and Fielding, 2004; Buatois and Mángano, 2011; Bayet-Goll and Neto De Carvalho, 2015; Bouchemla et al., 2020). In Algeria, the palaeoenvironmental interpretations of the Mesozoic strata have been significantly improved by an integrated approach that combines ichnological and sedimentological aspects (Bouchemla et al., 2020, 2021; Mekki et al., 2023). Accordingly, this study aims to describe and illustrate, for the first time, the trace-fossil assemblage from the Aptian Djebel Djerma sequence (Bellezma Mountains) to deduce the paleoenvironmental setting of the deposits.
2. Geological setting
The study area is located in northeastern Algeria, where two major orogenic systems developed between the Mediterranean Sea and the Sahara platform during the Late Cretaceous in response to the convergence of the African and Eurasian plates. The Tell domain to the north and the Atlas mountainous system to the south are the resultant orogenic belts (Figure 1). During the Mesozoic, both belts were situated in the southernmost region of the Tethyan margin.
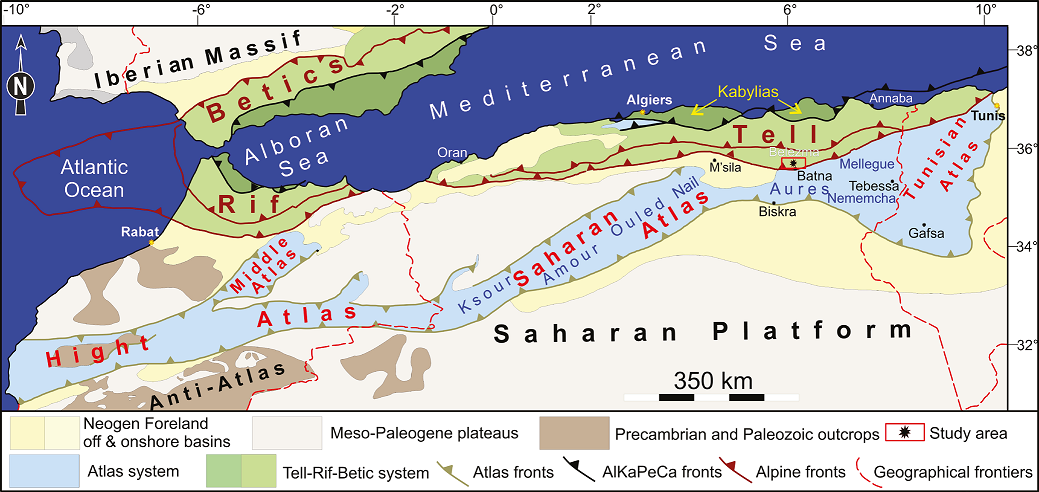
Figure 1 Structural map of the Western Mediterranean region showing the main orogenic systems and the position of the study area.
The Atlas includes the Saharan Atlas sensu stricto (Ksour Mountains, Djebel Amour, and Ouled Nail Mountains), the Aures Mountains, and the alignment formed by the Nememcha, Tebessa, Morsott, and Mellegue Mountains.
The section investigated (geographic coordinates: X = 6°26’03’’E; Y = 35°68’20” N) belongs to the southern Bellezma Mountains, at the confluence of the Tell Atlas and the Saharan Atlas (Bellion, 1972), more precisely, in the Djerma area, a few tens of kilometers from Batna town (Figure 1). The sedimentation in this area is mainly dominated by carbonate deposits, with a minor presence of sandstones. In the northern region, the deposits are distinguished by a siltstone-dominated facies, reaching a thickness of approximately 300 meters , which was deposited in a shallow marine environment. This sequence overlain by a carbonate series.
3. Material and methods
During the fieldwork in the studied area (southern Bellezma Mountains), a meticulous approach was adopted at the outcrop while documenting and describing the trace fossils since all the recorded ichnotaxa are associated with comparatively indurated limestone beds. In the light of these circumstances, all the trace fossils were photographed under natural light, documented, and identified in the field while respecting different parameters of trace morphology and pattern, following the standard procedural practice of Bromley (1996).
Furthermore, in order to frame trace-fossil information, consideration has been given to ichnotaxon identification (e.g.Häntzschel, 1975; Uchman, 1998; Seilacher, 2007), the toponomy classification (Martinsson, 1970), and trophic and ethological categories (cf. Bromley, 1996). A vertical profile section was constructed, highlighting trace fossil horizons and related sedimentary features to support the interpretation of the palaeoenvironment. The sedimentary features can be defined in terms of the lithology, faunal content, geometry of beds and contacts, sedimentary structures, bounding surface, and bed thickness. Textural features of the strata containing trace fossils were also noted and studied, following the nomenclature and classification of carbonate rocks (cf. Dunham, 1962; Embry and Klovan, 1971).
4. Description of the studied section
Within the Bellezma Mountains, on the northern side of the Aurasian basin, the Lower Cretaceous outcrops are continuous on the northern flank of the Batna Mountains. They form parallel ridges oriented roughly WSW-ENE. The succession studied (Figure 2) consists mainly of an alternation of green marl and bioclastic limestone beds ranging from 0.1 to 0.8 m in thickness. The limestone beds extend laterally for tens of meters, exhibiting a channelized morphology with a sharp or slightly erosional base, and forming metric ridges towards the top of the section. Some limestone beds show pinch-and-swell geometries. The upper surfaces of the beds commonly display well-developed ripple marks.

Figure 2 A, measured lithostratigraphic section of the Aptian Djerma section; B, Close view showing orbitolinid, larger benthic foraminifera-rich limestone; C, Hummocky cross-stratified limestone bed; D, Bioclastic limestone rich in benthic crinoids (see arrows); E, Rudist-rich limestone bed; F and G, Ooid-bioclastic grainstone with numerous remains of benthic crinoids (Cr), orbitolinids (Or) and bryozoans (Br).
The part of the succession that revealed trace fossils is located in the middle part of the succession. It consists of green marls interbedded by burrowed bioclastic limestone beds rich in crinoids (Figure 2C) and larger benthic foraminifera (orbitolinids), associated with bryozoan, gastropod, and bivalve fragments (Figure 2B). The marl interval beds between the limestones yielded abundant ostracods, mainly with smooth valves, agglutinated benthic foraminifera, bryozoans, crinoids, and gastropod and echinoid debris. Based on the stratigraphic value of the orbitolinid foraminifera, an age of middle Aptian is constrained for the crinoid- and orbitolinid-bearing beds (Bureau, 1971, p. 379, 383).
The microfacies of the studied trace fossil-bearing limestone beds show ooid-bioclastic grainstone and, less often, bioclastic packstone. Erosional intraclasts formed by mud-dominated wackestone with dispersed fine ooids and fine bioclasts were observed. Among the bioclasts, there are numerous remains of benthic crinoids and orbitolinid, larger benthic foraminifera (Figure 2F and 2G). Moreover, remains of dasycladalean green algae (cf. Bucur et al., 2012), gastropods, small benthic foraminifera (e.g. lenticulinids), echinoid spines, and bryozoans are common in the sediment. In the grain-supported sediments, burrows filled with fine bioclastic packstone were observed. Detritical quartz grains up to 0.2 mm commonly occur in the sparite matrix.
5. Systematic ichnology
In the present study, eight ichnogenera were identified, showing moderate ethological diversity (Table 1). Overall, the trace-fossil assemblage is characterized by low diversity, occurring exclusively in the middle part of the studied succession, in which the most abundant traces are Thalassinoides. The current study uses the binomial system of nomenclature to designate ichnogenera and ichnospecies in accordance with I.C.Z.N. guidelines, and it describes them alphabetically. Recovered trace fossils are as follows:
Ichnogenus ArchaeonassaFenton and Fenton, 1937
Archaeonassa isp.
Table 1 Classification of the recorded ichnotaxa from the studied succession of the Bellezma Mountains. (abundant-common-rare)
| Ichnotaxa | Ethology | Toponomy | Occurrence | Possible producers | References |
| Archaeonassa isp. | Pascichnia | Epichnia | Common | Mollusks | (Buckman, 1994) |
| Cochlichnus cf. anguineus | Repichnia | Epichnia | Common | Annelids | (Buatois et al., 1997) |
| Chondrites isp. | Fodinichnia/Chemichnia/Agrichnia | Endichnia | Rare | Divers (e.g. annelids) | (Baucon et al., 2020) |
| Gyrochorte comosa | Pascichnia | Epichnia | Common | Worm-like organism | (Fürsich et al., 2017) |
| Palaeophycus isp. | Domichnia | Endichnia | Rare | Polychaete annelids | (Pemberton and Frey, 1982) |
| Protovirgularia dichotoma | Repichnia | Epichnia | Common | Malacostracan crustaceans (e.g. isopods) | (Knaust, 2022, 2024) |
| Rhizocorallium cf. commune | Fodinichnia/Domichnia | Epichnia | Rare | Annelids (polychaetes) | (Knaust, 2013, 2017) |
| Thalassinoides isp. A | Domichnia | Endichia | Abundant | Crustaceans | (Frey et al., 1984) |
| Thalassinoides isp. B | Domichnia | Epichnia | Common | Crustaceans | (Frey et al., 1984) |

Figure 3 Invertebrate trace fossils from the Middle Aptian sedimentary succession of the Bellezma Mountains. A, Archaeonassa isp. as positive epirelief in medium-grained limestone; B and C, Cochlichnus cf. anguineusHitchcock, 1858; sinuous trails preserved as negative epirelief in medium-grained limestone; D, Chondrites isp. in full relief (endichnia) preservation, parallel to bedding plane; E and F, biserially arranged plaited ridges of Gyrochorte comosa Heer, 1865, preserved as positive epirelief; G, Palaeophycus isp. in full relief (endichnia) preservation; H, straight or slightly curved, small keel like trails of Protovirgularia dichotomaMcCoy, 1850, preserved as positive epirelief.
Description. Epichnial, horizontal, bilobed simple, unbranched, straight, curved to locally meandering trail flanked by two faints, parallel, more or less discontinuous lateral ridges run in both margins of regular furrow who is usually V-shaped in cross-section (Figure 3A). The trail width is about 5 mm, while the lateral ridges are 2 mm apart, and the maximum length observed is 125 mm. Sometimes the bilobed trails are cross-cutting each other.
Comment. The ichnogenus Archaeonassa is generally interpreted as a pascichnia produced by various invertebrates, including arthropods and mollusks (Buckman, 1994; Mángano et al., 2005). According to Fenton and Fenton (1937), Archaeonassa is typically preserved in intertidal zones where it may be abundant, and it may also occur more rarely in shallow-marine settings. More recently, Bouchemla et al. (2020) reported this ichnotaxon from an offshore environment. On the other hand, Archaeonassa is also known from continental deposits (e.g.Buckman, 1994).
Ichnogenus CochlichnusHitchcock, 1858
Cochlichnus cf. anguineusHitchcock, 1858
Description. Small, thin, sinuous or meandering, smooth, cylindrical, horizontal, unbranched, straight to gently curved path trails, preserved as a concave epirelief on the upper surface of medium-grained limestone bed (Figure 3B and 3C). The diameter of the trails mainly ranges between 3-4 mm. The length of the most complete trail is about 85 mm. The wavelength of the trails ranges between 9 and 11 mm, and the wave amplitude ranges from 4 to 5 mm. From above, the trace appears to have a slightly flattened groove.
Comment. Cochlichnus has been variously interpreted as locomotion, feeding, or grazing burrow (Buatois et al., 1997). The identity of the organisms responsible for the formation of this type of trace fossil remains uncertain, although burrows identical to them have also been found in recent non-marine deposits (Chamberlain, 1975; Ratcliffe and Fagerstrom, 1980; Metz, 1987). However, several candidates have been proposed, including annelids, especially nematodes (Seilacher, 1960) and insect larvae (Metz, 1987). The ichnogenus Cochlichnus has been reported from a variety of depositional settings, including both marine and non-marine deposits from the Ediacaran to Holocene (e.g.Hitchcock, 1858; Gluszek, 1995; Buatois et al., 1997; Keighley and Pickerill, 1997; Dernov, 2023).
Ichnogenus ChondritesVon Sternberg, 1833
Chondrites isp.
Description. Horizontal small tree-like, straight to slightly curved, branched, white-colored, flattened tunnels, with one order of branches, preserved exclusively in full relief (endichnia). The angle of branches is acute, usually between 40° and 45° (Figure 3D). Tunnel diameter is about 3-4 mm, while the overall burrow system can reach 40 mm in length.
Comment. Chondrites is a marine complicated trace fossil that can be classified ethologically as chimichnion, fodinichnion, or even agrichnion, with multiple probable trace maker options (Baucon et al., 2020). Chondrites occur in diverse marine settings, most likely associated with low oxygen and dysoxic conditions (e.g.Bromley and Ekdale, 1984; Fu, 1991; Bouchemla et al., 2021, 2023a, 2024).
Nevertheless, its scarce presence does not always provide conclusive evidence for under oxygenated seawater. In this way, Chondrites could be found within diverse and rich trace-fossil assemblages assigned to the Cruziana ichnofacies in shallow marine shelf settings, reflecting virtuous environmental conditions (e.g.Bouchemla et al., 2020; Mekki et al., 2023).
Ichnogenus Gyrochorte Heer, 1865
Gyrochorte comosa Heer, 1865
Description. Epirelief, unbranched, straight to slightly curved, long, bilobate burrows consisting of an elevated ribbon divided by a more or less distinct 0.5 mm-deep furrow in the middle. The ribbon’s width remains constant at 6 mm, but its length varies significantly across exposures (Figure 3E and 3F). In some cases, the burrow shows submeandering trails with the same color as the host rock, sometimes cross-cutting each other.
Comment. Gyrochorte is commonly interpreted as a pascichnion trace fossil left by a deposit-feeding worm-like organism moving obliquely through the sediment (e.g.Fürsich et al., 2017; Bouchemla et al., 2020; Mekki et al., 2023). This ichnofossil is commonly found in shallow marine environments during the Jurassic and Cretaceous (e.g.Gibert and Benner, 2002; Seilacher, 2007; Bouchemla et al., 2020; Mekki et al., 2023), commonly associated with thin ripple-bedded facies related to shelf setting. It is considered as one of the main components of the Cruziana ichnofacies.
Ichnogenus PalaeophycusHall, 1847
Palaeophycus isp.
Description. Endichnial, full relief, horizontal, cylindrical to sub-cylindrical, unbranched, simple, lined, straight to slightly curved burrow. It measures about 10 mm in diameter and has a variable length of up to 120 mm. The surface wall is clearly smooth without any distinctive annulations or striations. The structure of burrow infilling is indistinguishable from that of the surrounding host rock (Figure 3G).
Comment. Palaeophycus is commonly interpreted as a dwelling burrow (eurybathic domichnion) of predatory worms (?polychaete annelids) (Pemberton and Frey, 1982; Mekki et al., 2023; Bouchemla et al., 2024). However, according to Fürsich et al. (2018), the feeding mode of the producers is not excluded, even though it is more difficult to interpret. Thus, the producers may have been selective deposit-feeders or scavengers. Palaeophycus spans a wide spectrum of depositional settings from the Cambrian to the Recent (Pemberton and Frey, 1982).
Ichnogenus ProtovirgulariaMcCoy, 1850
Protovirgularia dichotomaMcCoy, 1850
Description. Elongated, horizontal, straight to curved, unbranched, plaited, chevron-shaped or keel-like trails, preserved as positive epirelief on the bedding plane. Its length is variable but can reach 130 mm, and its width is 7-8 mm. The burrows are characterized by a median 0.5-1 mm deep furrow, which is smooth and narrower, separating laterally two equidistantly distributed, inclined series of wedge-shaped appendages (Figure 3H). The composition of the burrow fill is identical to that of the host rock.
Comment. The ichnogenus Protovirgularia is commonly interpreted as locomotion activities (repichnion) of protobranch bivalves (Seilacher and Seilacher, 1994). However, according to Knaust (2024), the morphological characteristics support an old, established hypothesis that arthropods are the trace makers in most cases. Therefore, several lines of evidence suggest malacostracan crustaceans (e.g. isopods) as potential producers of many Protovirgularia (Knaust, 2022, 2024). It is commonly recorded from marine and continental deposits worldwide but is frequently associated with shelfal platforms, being considered a common component of the Cruziana ichnofacies (see Toom et al., 2019; Bouchemla et al., 2020; Mekki et al., 2023).
Ichnogenus RhizocoralliumZenker, 1836
Rhizocorallium cf. communeSchmid, 1876

Figure 4 Invertebrate trace fossils from the Middle Aptian sedimentary succession of the Bellezma Mountains. A, U-shaped, horizontal, spreiten burrow of Rhizocorallium cf. communeSchmid, 1876, preserved as epichnial, semi-relief in medium-grained limestone bed rich in Orbitolina fauna; B, T- to Y-shaped burrows of Thalassinoides isp. A in full relief (endichnia) preservation; C, three-dimensional Y-shaped burrows of Thalassinoides isp. B, preserved on the top surface of a medium- to coarse-grained, thin Orbitolina-bearing limestone bed as both convex and concave epirelief.
Description. Epichnial semi-relief, horizontal, unbranched, straight to gently curved, U-shaped spreiten burrow. Limbs are parallel, poorly preserved, and 7-8 mm in diameter. The spreiten are stout and prominent and are filled with finer sedimentary material than the host rock (Figure 4A). Both limbs and spreiten are totally devoid of tiny faecal pellets (Coprulus isp.). Burrow length varies amongst ichnotaxons, with a maximum recorded length of 120 mm and widths of 40 to 50 mm. This ichnotaxon is very common among others in the studied formation.
Comment. Rhizocorallium is a well-known ichnogenus from both siliciclastic and carbonate deposits (Knaust, 2017). It occurs within a wide environmental range, from non-marine to marine settings (e.g.Rodríguez-Tovar and Pérez-Valera, 2008; Knaust, 2013; Mekki et al., 2023). In marine successions, Rhizocorallium is produced most likely by annelids (polychaetes) or decapod crustaceans and is interpreted as a feeding (fodinichnion) or a dwelling (domichnion) behavior (Knaust, 2013, 2017; Zhao et al., 2020; Mekki et al., 2023).
Overall, this ichnogenus is commonly considered an indicator of shallow marine settings and one of the main components of the Cruziana ichnofacies (MacEachern et al., 2007; MacEachern and Bann, 2008; Buatois and Mángano, 2011).
Ichnogenus ThalassinoidesEhrenberg, 1944
Thalassinoides isp. A
Description. Endichnial, full-relief, horizontal, cylindrical to sub-cylindrical, T- to Y-shaped burrow system, spread on bedding surface with smooth to slightly coarse, elliptical to flattened tunnels that bifurcate at an angle of 45°-90°. The maximum observed burrow system is about 150 mm long and varies in diameter from 20-25 mm (Figure 4B). The color, texture, and burrow fill material are different from the host rock.
Thalassinoides isp. B
Description. Straight to slightly curved, horizontal to oblique, cylindrical tunnels, 9-11 mm in diameter, with dominantly regular Y-shaped branches. Some tunnels are truncated due to erosion and left as prominent grooves. The burrow is preserved on the top surface of a medium- to coarse-grained, thin limestone bed rich in Orbitolina fauna as both convex and concave epirelief (Figure 4C). The colour, texture, and burrow fill are identical to those of the host rock; however, the burrow fill shows well-preserved and imbricated Orbitolina fauna along the tunnel.
Comment. Thalassinoides is generally interpreted as a domichnial burrow, passively filled, usually related to well-oxygenated environments and soft but fairly cohesive substrates (Rodríguez-Tovar and Uchman, 2004; Bouchemla et al., 2020, 2023b). It is commonly considered a facies-crossing ichnogenus that occurs in a wide environmental range from shallow to deep sea settings (Myrow, 1995). However, it is a typical element of the shallow-marine environments, mainly attributed to crustaceans (Frey et al., 1984; Knaust, 2017; Bouchemla et al., 2025).
6. Discussion
Ichnological data from the Aptian sedimentary succession exposed in the southern Bellezma Mountains indicate a low diversity trace fossil assemblage, composed of eight ichnogenera: Archaeonassa, Cochlichnus, Chondrites, Gyrochorte, Palaeophycus, Protovirgularia, Rhizocorallium, and Thalassinoides. The latter is the most abundant ichnotaxon being recorded from the lower to the top of the section. The ichnoassociation is represented by post-depositional, non-graphoglyptid trace fossils dominated mainly by repichnia, pascichnia, fodinichia, and domichnia of deposit feeders (Table 1), reflecting the Cruziana ichnofacies, which is characterised by a wide variety of ethological categories, including dwelling, locomotion, resting, feeding, and grazing traces (MacEachern et al., 2007; Buatois and Mángano, 2011), with the secondary presence of irregularly inclined to horizontal components such as Thalassinoides (MacEachern et al., 2007). All obtained trace fossils in the studied section are horizontal to the bedding plan, and no vertical trace fossils were seen, probably reflecting the absence of suspension-feeders’ (infaunal) organisms in the bottom water. The vertical distribution of the recorded ichnofossils demonstrates an exceptionally high level of bioturbation in the middle part of the studied section within medium- to coarse-grained limestone beds. Its microfacial analysis evidenced an oobioclastic grainstone texture, less often, bioclastic packstone with abundant benthic crinoids, orbitolinids, larger benthic foraminifera, and numerous remains of dasycladalean green algae and bryozoans. These microfacial features represent transgressive, open marine (sub-)tropical shallow subtidal facies that developed in moderate energy within the inner platform carbonate sand shoals in normal salinity water conditions (e.g.Flügel, 2004). The extensive distribution of detrital quartz grains provides evidence of the impact of the surrounding land.
However, the frequency of high-energy hydrodynamic structures (e.g. HCS) and channelized limestone beds with sharp to slightly erosional bases may suggest that intense storm activity has impacted the bottom water between the fairweather wave base and the storm wave base (e.g.MacEachern et al., 2007; Bouchemla et al., 2020; Mekki et al., 2023), thus reflecting upper offshore to offshore transitional environments. In this context, the extremely abundance and high bioturbation index of the documented trace fossils in the middle part of this sedimentary succession is interpreted as resulting from an increase in food supply and oxygen levels following storm events.
Among the recorded ichnofossils, Gyrochorte comosa commonly occurs in nearshore and shallow marine deposits formed in moderate to moderately high energy environments, including storm-dominated shelves, bars, shorefaces of beach complexes, and embayment areas (e.g.Schlirf, 2000; Gibert and Benner, 2002; Seilacher, 2007).
More recently, Bouchemla et al. (2020) and Mekki et al. (2023) have reported this kind of trace fossil from storm-influenced continental shelf setting, predominantly in offshore environments, which means that this burrow is linked to tempestite layers and is considered a post-event trace (Bouchemla et al., 2020; Wetzel et al., 2020), implying that their producers managed to exploit the seafloor during quiet periods between high-energy events, most commonly storms (Gibert and Benner, 2002; Wetzel et al., 2020; Bouchemla et al., 2020; Mekki et al., 2023).
Archaeonassa is regarded as a repichnion burrow, produced by variant trace-makers including predatory gastropods (Fenton and Fenton, 1937; Buckman, 1994), nereidid polychaetes (Yuanyuan et al., 2019), and crustaceans (Yochelson and Fedonkin, 1997) occurring in both Skolithos and Cruziana ichnofacies, often recorded in the intertidal zones (e.g.Chen et al., 2013). However, Bouchemla et al. (2020) reported this ichnotaxon from offshore deposits within the distal part of the Cruziana ichnofacies.
Despite the ichnogenus Cochlichnus being widely reported from non-marine sediments (e.g.Dernov, 2023), it has recently been recorded within a storm-dominated shallow marine setting (upper offshore zone) by Bendella et al. (2022).
The authors identified this ichnogenus as one of the components of the Cruziana ichnofacies within the studied succession of the Teferguenite Formation. Protovirgularia has been documented by Bouchemla et al. (2020) from the Faïdja Formation (Kimmeridgian), Nador Mountains, and by Mekki et al. (2023) from the Teniet El Klakh Formation (Bajocian), Ksour Mountains, predominantly from upper offshore settings. According to Knaust (2023), Protovirgularia is facies-crossing, with only P. dichotoma and P. rugosa occurring in marine and, infrequently, continental settings, whereas the remaining ichnospecies are only known from marine environments.
However, Protovirgularia is a common ichnogenus of the Cruziana ichnofacies in shallow-marine, related every so often to oxygen-depleted environments within episodic tempestite deposits, which characterize upper offshore-offshore settings (e.g.MacEachern et al., 2007; Buatois and Mángano, 2011; Bouchemla et al., 2020; Mekki et al., 2023).
Rhizocorallium is a common complex trace fossil that is reported from continental to marine settings (Rodríguez-Tovar et al., 2012; Knaust, 2013; Zhang et al., 2016; Mekki et al., 2023). However, according to Zhao et al. (2020), this ichnofossil is considered as an indicator of shallow marine settings and regarded as a good tool to assess some ecological features (Knaust, 2013).
In the studied succession, Rhizocorallium is interpreted as a feeding burrow of deposit-feeders belonging to the Cruziana ichnofacies; it is considered as a component of quiet environments that appear in post-storm deposits when the hydrodynamic regime is rather calm and background sedimentation occurs (Rodríguez-Tovar et al., 2007; Mekki et al., 2023).
Thalassinoides is a facies-crossing genus and has been reported from a wide range of environments, from continental (Bouchemla et al., 2023b) to marine (Bouchemla et al., 2025) settings. However, the horizontal biogenic structures (i.e., Thalassinoides, Palaeophycus, and others) indicate a reduced energy level corresponding to lower shoreface-offshore marine settings (Ozukum et al., 2022). The presence of Chondrites, as observed by Bromley and Ekdale (1984), indicates lower oxygen levels. However, their occasional and rare occurrence, particularly in association with other endobenthos (e.g. Thalassinoides), suggests well-oxygenated bottom water (e.g.Ozukum et al., 2022).
The sedimentary features and ichnological data indicate that the middle part of the studied Aptian succession from the southern Bellezma Mountains was characterized by a well-oxygenated environment rich in organic matter and food supply. This allowed the development of an ichnoassociation that reflects the Cruziana ichnofacies, which developed in the upper offshore zone.
The data also show that the benthic community was predominantly deposit-feeding. However, the studied trace fossils show that at least some parts of these deposits were accumulated very close to or just below the fair-weather wave base (Pemberton et al., 2001). Furthermore, despite the favorable environmental conditions, the limited diversity of the studied assemblage may be attributed to the coarse nature of the sediments on the seafloor, which impedes the development and occurrence of the benthic community and, consequently, the documentation of their trace fossils.
7. Conclusion
The ichnological analysis of the Aptian sedimentary succession exposed in the southern Bellezma Mountains indicates that it was deposited in an upper offshore shallow marine environment with fluctuating energy regimes due to storm events. A low diversity association of trace fossils is mainly found in the middle part of the studied section within medium-to coarse-grained crinoid and orbitolinid-bearing beds. A total of eight ichnogenera were identified, dominated by feeding, crawling, and dwelling burrows and trails, reflecting the expression of the Cruziana ichnofacies developed in poorly sorted soft substrate in the upper offshore zone. Additionally, the trace fossil assemblages suggest that interstitial waters were well-oxygenated during the deposition of the studied succession.

