











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. Introduction
In the evolutionary history of cetaceans, the Oligocene is a period of outstanding relevance since it marks the appearance of the modern cetacean lineages (Boessenecker and Fordyce, 2014, Fordyce, 2005; 2015a; Marx et al., 2019). The family Eomysticetidae, which is the most basal toothless family of baleen whales (Chaeomysticeti), includes a variety of taxa (11 formally described species), ranging from the early Oligocene to Early Miocene age (Sanders and Barnes, 2002a; Okazaki, 2012; Boessenecker and Fordyce, 2014, 2015a, 2015b, 2017a, 2017b). Fossils have been found along both sides of the Atlantic Ocean (Germany and South Carolina, USA) and around the Pacific Ocean (Mexico, Japan, and New Zealand) (Sanders and Barnes, 2002a; Okazaki, 2012; Boessenecker and Fordyce, 2014, 2017a, 2017b; Hernández Cisneros and Nava Sánchez, 2022; Solis-Añorve et al., 2022).
In Mexico, Baja California Sur has important Upper Oligocene fossil deposits, because it contains fossil-rich strata, which are exposed in two lithostratigraphic units (Figure 1): the San Gregorio Formation, in La Purísima and San Isidro; and the El Cien Formation, distributed W and NW of La Paz (Hausback, 1984; McLean et al., 1986; Schwennicke, 1994; Fischer et al., 1995). These units contain a rich assemblage of still poorly known fossil cetaceans including archaeocetes: kekenodontids (Hernádez-Cisneros and Tsai, 2016), stem Mysticeti, for instance: stem mysticetes (i.e., aetiocetids) and stem chaeomysticetes (i.e. cf. Eomysticetus sp. and other eomysticetids) (Barnes, 1998; 2002; Hernádez-Cisneros et al., 2017; Solís-Añorve et al., 2019), stem odontocetes (i.e. Agorophius), and other material (Cruz-Marin, 1977; Hernández-Cisneros, 2018, 2022; Barnes, 1998; 2002). In the case of eomysticetids, well-preserved specimens have been reported from these formations. Fossils found near San Isidro at the base of the El Pilón Butte in upper Oligocene beds of the San Gregorio Formation (Figure 1) were tentatively referred to Yamatocetus canaliculatus (Cedillo-Ávila, 2018). In addition, a specimen from the El Cien Formation at San Juan de la Costa has been described as cf. Eomysticetus sp. (Hernández-Cisneros et al., 2017; Hernández-Cisneros and Nava Sánchez, 2022).
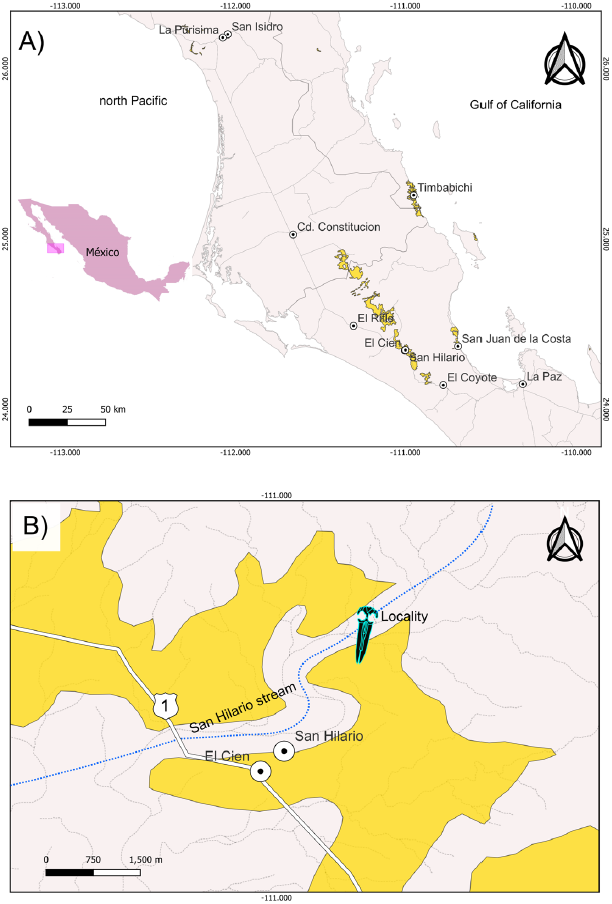
Figure 1 Maps of the Baja California Sur area showing the locality where the Eomysticetidae skull (IGM 7756) was collected. A) Yellow color delimits the areas in the state of Baja California Sur, where the Oligocene marine sediments crop out. The El Cien Formation is observed in the localities Timbabichi, San Juan de la Costa, San Hilario, and El Cien. On the other hand, to the north is the San Gregorio Formation in the locality of La Purisima. B) Detail map to the study locality (whale symbol). The material was collected in the San Hilario stream.
In the present study, we perform an anatomical and taxonomic analysis of an eomysticetid skull from the upper Oligocene San Juan Member of the El Cien Formation in Baja California Sur. The collection site is the so-called "Ten Minute Locality" (24°21'06.84" N, 110°59'27.24" W; Applegate, 1986) in Arroyo (dry stream) San Hilario, near the town of El Cien (Figure 1). The specimen IGM 7756 was collected in 1974 by Shelton P Applegate and Florencio Álvarez Cadena during an expedition in the Baja California Peninsula, sponsored by the National Geographic Society, and only briefly figured out by Downs (1974; Figure 2). This specimen was previously taxonomically identified as cf. Mauicetus sp. by Morris (1978), and posteriorly identified as a toothed mysticete by Barnes (1998; 2002); however, these authors did not provide illustrations and/or descriptions of the specimen. Furthermore, in order to obtain a better understanding and documentation of IGM 7756, we measured a 43.20 m thick geological section which includes the bed from which the fossil was recovered (Figure 3).
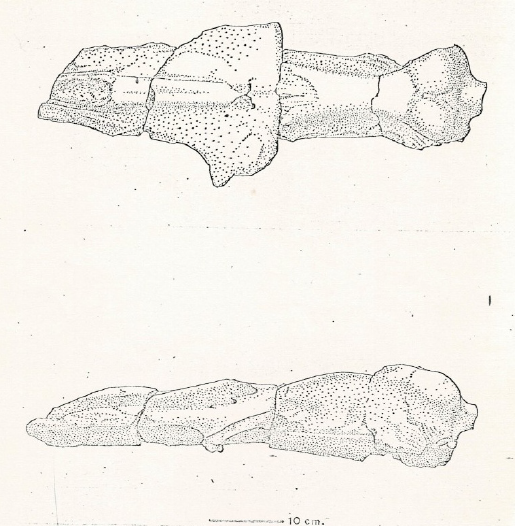
Figure 2 The original drawing of the skull of IGM 7756 in dorsal and left lateral view included by Downs, (1974), and identified like cf. Mauicetis sp.

Figure 3 The stratigraphic column was taken from the San Hilario section, which corresponds to the San Juan member of the El Cien Formation. A) Panoramic photography of the outcrop in the San Hilario stream. B) Facies of phosphatic sandstone, where the study material (IGM 7756) was collected. C) Fragment of tympanic bulla in the phosphatic sandstone layer. D) and E) Red lines mark the granular phosphorite layers that protrude into the stratigraphic column.
2. Geological Setting
The specimen was collected from in the El Cien Formation, a marine sedimentary succession exposed W and NW of La Paz along Federal Highway 1 (Figure 1), also exposed at San Juan de la Costa, and around Timbabichi towns (Hausback, 1984; Applegate, 1986; Fischer et al., 1995; Plata-Hernández, 2002; Drake et al., 2017). This formation comprises several lithostratigraphic subdivisions, named the San Juan, Cerro Colorado, and Timbabichi members (Fischer et al. 1995; Föllmi, et al., 2019; Plata-Hernández, 2002). The San Juan Member comprises depositional settings ranging from the coast to the outer shelf (Schwennicke, 1994). At the Ten Minute locality (Applegate, 1986), where the studied specimen IGM 7756 was collected, the middle part of the San Juan Member is exposed. Generally, the lithology at the Ten Minute Locality is dominated by alternating siliceous mudstone and tuffaceous mudstone, in addition to some clayey siltstone and tuffaceous clayey siltstone, tuff beds, and a few muddy phosphatic sandstone beds (Figure 3). The white-weathering siliceous mudstone ranges from nearly black to dark gray and may exhibit parallel lamination. Fish scales and planktonic foraminifera are common, in addition to a few mm-sized phosphate nodules. This lithology is more common in the lower part of the measured section. The tuffaceous mudstones display lighter tones ranging from dark to light grey and predominate in the middle and upper part of the column. The tuff beds are generally up to a few centimeters in thickness and include primary and resedimented tuffs. Phosphatic sandstone beds are rare in the lower and middle part of the measured column and are only a few centimeters in thickness, but there are thicker beds in the upper part. The most prominent bed at 32.10-32.90 m (about 80 cm in thickness) is a granular phosphorite; it contained the studied fossil. Another very phosphate-rich bed is found at 33.90 - 34.20 m and bears some mudstone intraclasts, shells, fish impressions, and turtle bone fragments. Altogether, the whole exposed succession reflects an open-shelf depositional environment.
The age of the succession exposed at the Ten Minute Locality is constrained by radiometric and biostratigraphic age data. A tuff bed interbedded in the middle part of the San Juan Member at Arroyo San Hilario dated a K/Ar age in biotite of ~ 25.5 +/- 0.4 Ma (Chattian, late Oligocene; Hausback, 1984). This dating is consistent with biostratigraphy data, specifically with the presence of the diatom index species Rocella vigilans (Fenner, 1984) and Bogorovia veniamini (Jousé, 1974) (Kim and Barron, 1986). In addition, new U-Pb zircon ages from interbedded tuffs in the middle part of the San Juan Member at San Juan de la Costa range between 28.37 +/- 0.45 and 26.75 +/- 0.17 Ma (late Rupelian -early Chattian; Schöllhorn et al., 2020).
3. Materials and methods
Anatomical identification and abbreviations are based on Mead and Fordyce (2009) and Boessenecker and Fordyce, (2014, 2015a, 2015b). Skull measurements were taken from ventral, dorsal, posterior, and lateral views. Measurements are summarized in Table 1. Photographs were taken with a camera Nikon D3300 with a lent AF-S Nikkor 18 - 55mm, using the "stacking" technique (Bercovici et al., 2009) to recover deepness and improve the focused coverage of the dorsal, ventral, posterior, lateral, and anterior views.
Table 1 Skull measurements.
| Dorsal view | mm |
| Preserved total length | 614* |
| Preserved length of the nasal | 209* |
| Nasals width in posterior portion | 11 |
| Width of the nasal at the most anterior portion | 19 |
| Maximum transverse width of the nasals | 28 |
| Preserved length of premaxilla | 300* |
| Posterior width of preserved length the premaxilla | 14* |
| Anterior width of the premaxilla | 18 |
| Length of maxilla | 333* |
| The greater width of the preserved maxilla | 65 |
| Length of parietal, from occipital apex to fronto parietal suture | 184 |
| Ventral view | |
| Width of the basioccipital crest | 28 |
| Width of the foramen magnum preserved | 49 |
| Tympanic Bulla | |
| Preserved length of the tympanic bulla (ventral view) | 51* |
| Total width of the tympanic bulla (taken at the midpoint) | 34* |
In addition, the skull was CT-scanned in a clinical tomograph from Imagenus laboratory, with a 0.26 mm width spacing, to generate 3D models and delimitate internal and external sutures. Tridimensional models of the ear region and skull were carried out with digital data in 3D Slicer 5.0.3 (Fedorov et al., 2012); the models were visualized in the program MeshLab v.2022.02 (Cignoni et al., 2008).
4. Results
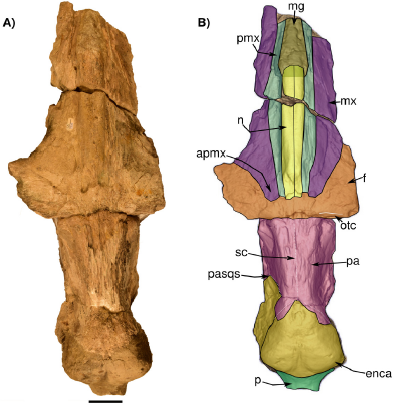
Figure 4 Skull in dorsal view. A) Photograph of IGM 7756. B) 3D Model obtained with the CT images highlighting its bones. The dotted line at the supraoccipital marks approximately its original rounded shape. Abbreviations: apmx, ascending process of the maxilla; enca, endocranial cast; f, frontal; mg, mesorostral groove; mx, maxilla; n, nasal; p, pedicle at the base of the occipital condyles; pa, parietal; pmx, pasqs, squamosal parietal suture; premaxilla; otc, orbitotemporal crest; sc, sagittal crest. Scale bar 5 cm.
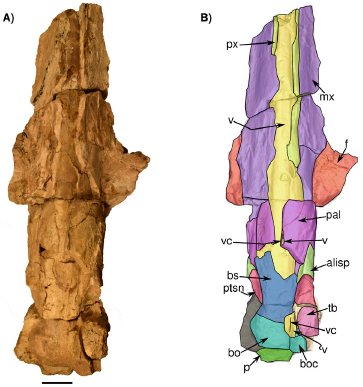
Figure 5 Skull in ventral view IGM 7756. A) a Photograph of the individual and B) 3D Model obtained with the CT. Abbreviations: boc, basioccipital crest; bs, basisphenoids; fsp, fragments of the sigmoid process; mx, maxilla; p, pedicle at the base of the condyles; pal, palatine; px, premaxilla; ptsn, pterygoid sinus; tb, tympanic bulla; v, vomer and vc, vomer crest. Scale bar 5 cm.
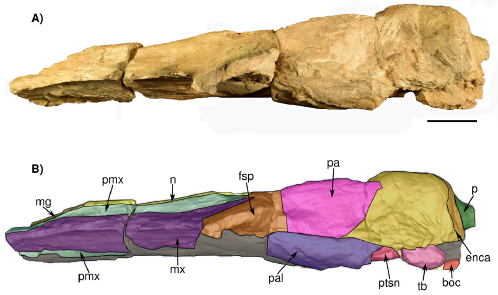
Figure 6 Skull in left lateral view IGM 7756. A) Photograph and B) 3D Model obtained with the CT. Abbreviations boc, basioccipital crest; enca, endocranial cast; fsp, supraorbital process of the frontal; mg, mesorostral groove; mx, maxilla; n, nasal; p, pedicle at the base of the condyles; pa, parietal; pal, palatine; pmx, premaxilla; ptsn, pterygoid sinus; ppt, pterygoid; tb, tympanic bulla. Scale bar 5 cm.
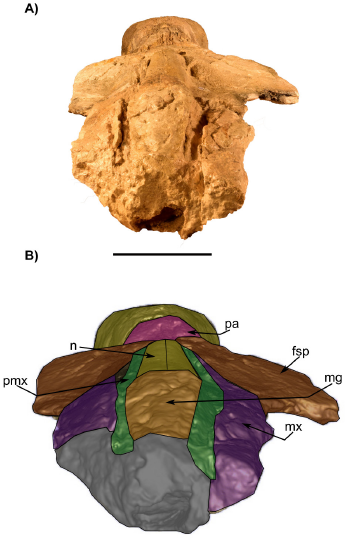
Figure 7 Skull in anterior view IGM 7756. A) photograph and B) 3D Model obtained with the CT. fsp, supraorbital process of the frontal; mx, maxilla; n, nasal; pa, parietal; pmx, premaxilla; mg, mesorostral groove. Scale bar 5 cm.
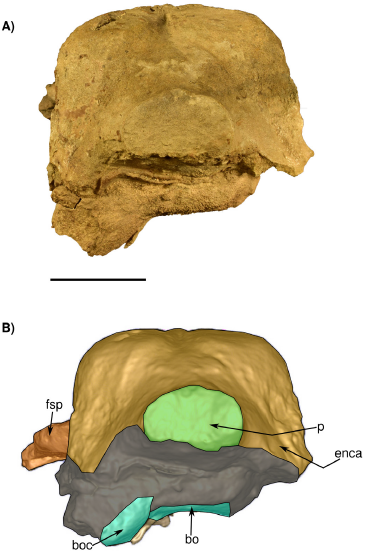
Figure 8 Skull in posterior view IGM 7756. A) Photograph and B) 3D Model obtained with the CT. bo, basioccipital and boc, basioccipital crest; enca, endocranial cast; fsq, supraorbital process of the frontal; p, pedicel at the base of the condyles. Scale bar 5 cm.
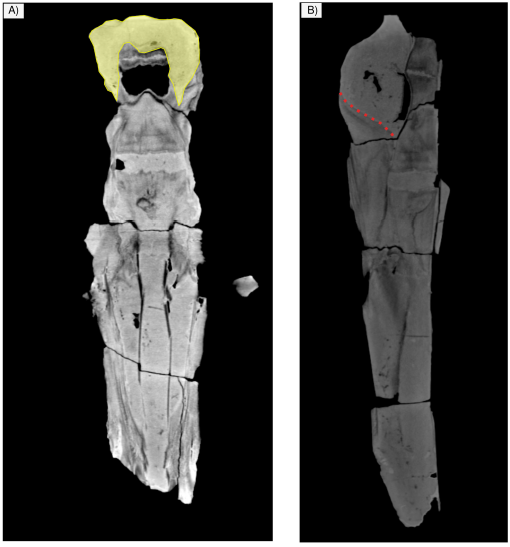
Figure 9 Tomography images of IGM 7756. A). The skull in dorsal view shows the endocast of yellow color is observed. B) The skull in lateral view shows the squamo-parietal suture is observed with dotted lines, this suture is not observed with the naked eye in the material.
Institutional abbreviations IGM-Instituto de Geología, Universidad Nacional Autónoma de México, México.
Referred material. IGM 7756, incomplete skull including left tympanic bulla.
Locality precedence. Ten Minute locality, 24° 21' 06.84" N; 110° 59' 27.24" W, 1.5 kilometer east of kilometer 100 of Mexican Highway 1, Baja California Sur, Mexico (Figure 1).
Horizon and age. San Juan Member, El Cien Formation, upper Oligocene; Chattian (Hausback, 1984; Kim and Barron, 1986; Schöllhorn et al., 2020) (Figure 3).
Systematic attribution. IGM 7756 is identified as a Neoceti based on the following derived characters: partly open mesorostral groove; a posterior displacement and expansion of the premaxilla and maxilla; and a narrow intertemporal region. Further differs from odontocetes in the lack of facial concavity and posterior expansion of maxilla over the supraorbital region; the lack of premaxillary sac fossa and premaxillary foramen; and in having the maxilla-frontal suture straight. IGM 7756 is identified as a Mysticeti based on: bulbous and transversally thickened basioccipital crest; posterior region of the maxilla placed lateral to nasal; and elongated nasals. IGM 7756 differs from Mystacodon selenensis for the absence of the following characteristics: the maxilla forms excluding the anteriormost portion of the rostrum formed by the premaxilla and most of the palate; the nasal being longer than the length of the frontal + parietal; the ascending process of the maxilla and premaxilla extends posteriorly at the posterior margin of the nasals. IGM 7756 is different from Llanocetus denticrenatus in having an anteroposteriorly longer intertemporal region, in lacking a sagittal trough on the dorsal surface of the parietals; the nasals at their posterior limit are almost at the level of the maxillae. IGM 7756 differs from aetiocetids in not presenting a triangular anterior extension of parietal-frontal suture; supraorbital processes of the frontal are not elevated and at the same level as the vertex of the skull; and showing greater transverse intertemporal width. It differs from mammalodontids because it; lacking a triangular wedge of frontal that separates the posteromedial edge of the ascending process of maxilla from the posterolateral margin of the nasal and is viewed laterally; not presenting a waker such a marked intertemporal constriction. IGM 7756 further differs from basal Chaeomysticeti in the absence of a frontal-maxilla contact that is loose and sutured only along the ascending process of maxilla. Differs from Maibalaena nesbittae in lacking frontal-parietal sutures that converge posteriorly with the frontals penetrating between the parietals and in the posterior border of the nasal not being at the level of the preorbital. IGM 7756 differs from Sitsqwayk cornishorum in the maxilla at its posterior border is almost at the level of the nasals; not having a dorsal surface of the nasals that forms a sagittal keel. IGM 7756 is identified as an Eomysticetidae according to the following characteristics: elongated nasals; postotbital ridge absent; including a frontal with anteromedial projection; anteroposteriorly long and transversely narrow intertemporal region, and oval temporal fossa (longer anteroposteriorly than trancersely wide. IGM 7756 differs from crown Chaeomysticeti parietal exposure at the vertex is the posterior half of the temporal fossa; the nasal-frontal suture reaching the level of the anterior border of the interorbital region of the frontal; and the anterolateral intertemporal constriction being long.
General description. In general, the skull was probably exposed to weathering for an extended time since the dorsal and ventral surfaces are eroded and have an irregular texture; therefore, most of the sutures are difficult to trace. Much of the ear region is lost and deformed. In dorsal view, the posterior region is fragmented, lacking most of the squamosals and part of the exoccipitals, and the "occipital condyles" only preserve the pedicle at the base of the condyles where they originally projected. The foramen magnum is filled with sediment. The occipital shield is not preserved (i.e. supraoccipital) and only the exposed endocranial cast is observed (Figures 4, 8, and 9). Most of the rostrum is lacking, only preserving nasals, premaxillae, and a posterodorsal portion of both maxilla. Much of the ventral surface is missing, so the palatine, vomer, and rostrum cannot be described in detail. A portion of the right tympanic bulla is preserved in situ. However, no diagnostic features could be found, since it is eroded, even the dorsal region as seen in the CT scan.
Nasal. The nasals are almost completely preserved, lacking the anteriormost margin and probably part of the posteriormost. No exposed foramina were observed on the dorsal surface. Each nasal is anteroposteriorly elongated and transversally narrow, and both nasals are sutured along the midline. In dorsal view, the nasals extend slightly more posteriorly than the premaxillae and contact with both premaxillae along a visible and straight suture (Figure 4). Posteriorly, the naso-frontal suture is irregular, and extends posteriorly until almost the same level as the maxilla. Anteriorly to the nasal, the mesorostral canal is filled with sediment. In the left lateral view (Figure 7), the nasals are placed slightly above the level of the premaxilla with an almost flat-shaped dorsal outline.
Premaxilla. Only the posterior region of both premaxillae is preserved. They extend posteriorly, forming the ascending processes of the premaxillae almost to the level of the posterior margin of the nasals; the left premaxilla seems to develop more posteriorly because this area is fragmented on the right side (Figure 4); parallel longitudinal sutural ridges and grooves are present on the frontal, showing the premaxilla-frontal suture. The premaxilla is narrow at the level of the ascending process; it becomes wider at the level of nasals, and narrows again in anterior direction. Medially, the premaxilla contacts with the nasal while laterally it contacts with the maxilla; in lateral view, a fracture of the anterior portion of the rostrum produced a convex raised dorsal surface of the premaxilla. In ventral view, the premaxilla is visible in the palatal region, with more preserved portions on the right than on the left side; it develops as a narrow bone that medially contacts the vomer (Figure 5).
Maxilla. The maxillae are incomplete and only the ascending process and a thin sheet of bone in the posterior region of the rostrum are preserved. In dorsal view, over the posterior region, the ascending process of the maxilla forms a narrow protrusion that extends almost to the margin posterior tip of the nasals. Posteriorly, the frontomaxilla suture is well-defined (Figure 4). The lateral edges of the maxilla are not preserved. In dorsal view, the maxillae are wide in the anteroposterior direction so that their width is twice the wide of the premaxilla. Also, in lateral view, the dorsal surface of both maxillae is slightly concave to flat in their whole extension, but in the region of the ascending process of the maxilla the dorsal surface becomes convex (Figure 6). In ventral view, the maxillae are medially separated by the vomer and premaxillae (Figure 4). No palatal foramina were identified along the ventral surface of the maxilla.
Frontal/Parietal. The frontal is preserved in the skull roof and both supraorbital processes are broken at their base (Figure 4, 5, 6, 7). In lateral or anterior view, the preserved part of the left supraorbital process is horizontal, and the anterior margin is slightly concave. In dorsal view, on the right side, there is a low orbitotemporal crest that develops lateromedially, near the posterior border of the frontal bone (Figure 4). Besides, there are supraorbital foramina on the dorsal surface of the supraorbital process. The parietal is well exposed along the intertemporal region, being longer than wide (length = 184 mm; wide = 131 mm). Both parietals are fused along the midline, forming a low sagittal crest, detectable only by touch due to the preservation of the bone surface. Moreover, it forms the medial wall of the temporal fossa, which is slightly concave. The parietal-frontal suture is not very clear, but as far as can be traced on the left side it has an irregular-shaped outline. On the other hand, an interparietal is not evident in this specimen.
Both zygomatic processes are missing, while the squamosal forming the medial wall of the temporal fossa is badly preserved, exposing in some regions part of the endocast (Figure 6). The parietal-squamosal suture (better seen on the left side) is well-defined and develops dorso-ventrally and slightly in an anterior direction (Figure 6).
Occipital. The supraoccipital is not preserved (Figure 4), and in the occipital region, the exposed endocast is observed in dorsal, lateral and posterior views. (Figure 9). The posterior region of the skull including the exoccipital, condyles, and foramen magnum is also badly preserved (Figures 4 and 8). The occipital condyles are almost lacking only preserving part of the condylar neck. The foramen magnum is oval in transverse section and dorsoventrally compressed (Figure 8). The ventral surface (and all the associated structures) of the exoccipitals is not preserved.
Palatine/Pterygoid/Vomer. A small portion of the palatine is preserved (Figure 5). In ventral view, some palatine fragments are located posteriorly on the left and right sides. Medially, due to the incomplete preservation of the maxilla/palatine, the vomer is exposed in the ventral surface of the skull. A fragment of vomer is observed on the ventro-lateral surface of the basioccipital. Posteriorly to the palatines, portions of the left and right pterygoids are preserved. On the left side, a well-defined cavity corresponding to the pterygoid sinus fossa develops anteriorposteriorly (Figure 5). In dorsal view, the vomer is not seen in detail in the mesorostral groove as it is filled with sediment (Figure 4).
Basioccipital/Basisphenoid. The basioccipital is poorly preserved. In ventral view, it is wide and rectangularly shaped, and the preserved left basioccipital crest projects latero-ventrally. The left basioccipital crest is deformed posteriorly, and the medial surface is flattened; it is transversally wide. In the posterior view, the ventral surface of the basilar part is flat (Figure 5). In ventral view. the basioccipital-basisphenoids suture is found, running horizontally, below the most anterior point of the pterygoid. his suture is very tenuous but, on the sides, it is well defined.
Tympanic bulla. Only the left tympanic bulla is preserved, and it is in situ. Its ventral surface is eroded and slightly deformed and most of the posterior and lateral portions of the bulla is missing. Due to the erosion of the ventral surface, it is difficult to observe if the lateral and medial lobules present any kind of elevation or globose shape (Figure 5). The tympanic bulla anteroposterior length is 51 mm and its lateromedial width is approximately 31 mm. On the medial margin of the tympanic bulla, a ventromedial ridge is observed, extending in anteroposterior direction. The internal part of the tympano-periotic region was analyzed using tomography. However, the periotic could not be identified and segmented based on the available CT scan data. The dorsal surface of the tympanic bulla is eroded and slightly deformed as observed in the CT scans.
5. Discussion
The fossil record of the family Eomysticetidae is represented by at least 11 described species that show an extensive record in the Pacific basin, as well as on the Atlantic side, due to the biogeographical interactions recorded during the Oligocene, with a record from Kitakyushu, Japan (Okazaki, 2012), South Carolina, USA (Sanders and Barnes, 2002b), Kaiserswerth, Germany (Sanders and Barnes, 2002a), Baja California Sur, Mexico (Cedillo-Ávila, 2018; Hernández-Cisneros and Nava-Sánchez, 2002; Solís-Añorve et al., 2022), and South Island, New Zealand (Boessenecker and Fordyce, 2014, 2015a; 2015b; 2017a, 2017b).
The Eomysticetidae specimen IGM 7756 reported in this work is the third record for this baleen whale family for the late Oligocene of Baja California Sur, Mexico, and the second for the El Cien Formation (Cedillo-Ávila, 2018, Hernandez-Cisneros and Nava-Sánchez, 2022, Solis-Añorve et al., 2022). Even though the specimen is very incomplete, after a detailed comparison with other cetaceans and mysticetes (see diagnosis), we can confidently attribute it to Eomysticetidae based on a), elongated and thin nasals, b) a narrow intertemporal region, and c) an anteroposteriorly elongated temporal fossa, d) postorbital ridge absent (see Sanders and Barnes, 2002b; Boessenecker and Fordyce, 2015a, 2015b). Originally Downs, (1974) and Morris (1978) identified IGM 7756 as cf. Mauicetus sp. Later, Mauicetus (Benham, 1937; 1939; 1942), was reclassified into Eomysticetidae as Tokarahia lophocephalus, and Tohoraata waitakiensis (Steeman, 2007; Boessenecker and Fordyce, 2015b). However, IGM 7756 differs from Tokarahia lophocephalus in lacking numerous foramina in the supraorbital process of the frontal, nasal, and premaxillae (Boessenecker and Fordyce, 2015b). We were unable to compare IGM 7756 with the holotype of T. waitakiensis because there are not enough elements preserved in IGM 7756. Subsequently, Barnes (2002) identified IGM 7756 as a toothed mysticete, based on the presence of long nasals, a narrow intertemporal region, and laterally sloping maxillae; however, the specimen does not preserve teeth to consider it a toothed mysticete. Moreover, it differs from the toothed mysticete (i.e. Aetiocetus) in the following ways: the posterior border of the nasals is not at the same level as the premaxilla and maxillae; it lacks a triangular anterior extension of parietal-frontal suture; the supraorbital process of the frontal is not at the same level as the vertex; and the transverse intertemporal process is wider than in aetiocetids. (e.g. Deméré and Berta, 2008; Marx et al., 2015; Peredo and Pyenson, 2018; Shipps et al., 2019). In addition, it differs from other contemporary eomysticetids, for example, Eomysticetus whitmorei in having a straight facial portion of the rostrum in lateral view, a basioccipital slightly vaulted transversely, and a sagittal crest at the top of parietals in the intertemporal region (Sanders and Barnes, 2002b). Also, it differs from Yamatocetus canaliculatus the anterior margin of the nasals is anteriorly convex. It further differs from Micromysticetus rothauseni in having a basioccipital crest angular; with the ventral surface of the basioccipital being flat (Sanders and Barnes, 2002a). The specimen IGM 7756 also differs from Matapa waihao in having an orbitotemporal crest positioned entirely dorsally, a premaxilla extending further posterior than the nasal, and anterolaterally convex subtemporal crest (Boessenecker and Fordyce, 2017b). Likewise, it differs from Waharoa ruwhenua in having a basisphenoid slightly narrower than the basioccipital crests, and nasals with subparallel margins (Boessenecker and Fordyce, 2015a).
Due to diagnostic regions of the skull missing in IGM 7756, it can only be confidently assigned to the Eomysticetidae family, as a gen. et sp indet. However, the specimen shares some characteristics with Eomysticetus, such as an elongate and narrow intertemporal region (although in IGM 7756 the intertemporal region is not as narrow as in Eomysticetus), basioccipital crest short anteroposteriorly and wide transversely and an irregular fronto-parietal suture. We hope that the collection of more complete specimens will allow a more precise taxonomic assignation.
Previously, the specimen MHN-UABCS ECSj1/129/142, from the El Cien Formation at San Juan de la Costa locality, was identified with an affinity to Eomysticetus sp. (Hernandez-Cisneros and Nava-Sánchez, 2022). This material is poorly preserved, making comparisons difficult. However, it differs from IGM 7756 the following anatomical characteristics: it displays wide premaxillae parallel to the nasals, an ascending process of the premaxilla extending farther than the posterior region of the nasals, and a wider mesorotral groove, all those features are not found in IGM 7756.
Our knowledge of the evolutionary history of the Eomysticetidae has recently been extended by new records from the Pacific Ocean in both the Northern and Southern hemispheres (e.g., eomysticetids from New Zealand and Mexico). In particular, the presence of eomysticetids along the northeastern Pacific coast of Mexico (Barnes, 1998; 2002; Hernández-Cisneros et al., 2017, Hernández-Cisneros and Nava-Sánchez, 2022) has provided more information on paleobiogeographic events for this early baleen whales family.
6. Conclusion
In Baja California Sur the family Eomysticetidae is reported from the El Cien Formation and the San Gregorio Formation. The El Cien Formation is the second record of this family and the first for the San Hilario area. Although the specimen is poorly preserved, we can confidently assign it to this family as gen. et sp. indet. with more similarities to the North Atlantic (e.g. Eomysticetus) than to the South Pacific taxa (e.g. New Zealand). In recent years, the study of cetaceans from the Baja California Sur peninsula, Mexico, has improved our understanding of the evolution of cetacean diversity in the North Pacific basin. Future discoveries of better-preserved eomysticetid specimens from the Oligocene of Baja California Sur may provide us with new data that can help us better understand the evolutionary history and biogeography of this extinct mysticete family.
Contributions of authors
All authors contributed to the project planning. Solis-Añorve Azucena, description, research, writing, analysis, and comparative work. González-Barba Gerardo contributed to discussions, writing, and interpreting geology locality. Buono R. Mónica, writing, comparative and analysis of taxon. Schwennicke Tobias, stratigraphy section, interprets and describes the geology locality. Díaz-Cruz Jesús Alberto, analysis CT scanning and modeling of digital data, and editions to figures. Financing The first author received scholarship support of Consejo Nacional de Ciencia y Tecnología (CONACYT).
Conflicts of Interest
We have no conflicts of interest for the publication of this work.
Handling editor
Alexander Correa Metrio

