











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. Introduction
Our knowledge on the Miocene mollusk fauna of Makran region from Iranian platform is very sparse and still largely ambiguous, whereas this significant group of invertebrates shows widespread distribution in the overall Tethys Ocean during the Miocene timespan. These assemblages are in fact well studied in different basins from the western Tethys/proto-Mediterranean and the Paratethys (Mandic et al., 2020a, 2020b; Harzhauser et al., 2023), the eastern Tethys/proto Indo-Pacific (Harzhauser et al., 2009; Bose et al., 2021, 2023) and even the Caribbean region (Perrilliat et al., 2010). In Iran, only one deep study was carried out on this same group of the Miocene deposits in detail by Harzhauser et al.(2017).
However, they considered Burdigalian (Early Miocene) as the age for the mollusk assemblages with emphasis on the findings of McCall et al. (1994) and Ghaedi et al. (2016) from the Bande-Chaker Unit.
In this study, we aim to show a high diversity of mollusk faunas in accordance with calcareous nannofossil biozones that are assigned to the Upper Miocene (Tortonian) sediments for the first time from the Makran region.
2. Regional geology
The Iranian plateau was involved in the Alpine-Himalayan orogeny and has been classically subdivided into eight sedimentary-structural provinces, each one characterized by a different geological history (Stöcklin, 1968). From the north to the south these are: (1) the Kopet-Dagh Zone, NE Iran, (2) the Alborz Zone, N-NW Iran, (3) the Central Iranian Block or Cimmeria, consisting of three major continental blocks (Lut, Tabas, and Yazd) separated by major faults (Alavi, 1991), (4) the eastern Iranian suture zone, (5) the Urumieh-Dokhtar (Sahand-Bazman) magmatic arc, (6) the Zagros Zone, (7) the Sanandaj-Sirjan Zone, and (8) the Makran Zone (Figure 1a). The Makran zone is one of the largest accretionary wedges in the world about 350 km wide that is results from nearly N-S convergence between Arabia and Eurasia (e.g. DeMets et al., 2010) and grows both vertically and laterally by scraping and incorporating sediments off the subducting Arabian Plate lithosphere (Platt et al., 1985; Burg, 2018). Makran accretionary wedge is represented by high sediment thickness of about 7 km in the Gulf of Oman (Fruehn et al., 1997; Kopp et al., 2000). This zone is known to have complex tectonic settings being near to the Triple Junction, where three tectonic plates (i.e. Eurasian, Indian, and Arabian) are interacting (e.g. Abbasi, 2020). Makran region is separated from the Zagros Mountain Belt, to the west, by the dextral Minab-Zendan Transform Fault and to the east it is restricted by the sinistral Chaman Transform Fault System (Stöcklin 1968; Bird et al. 1975).
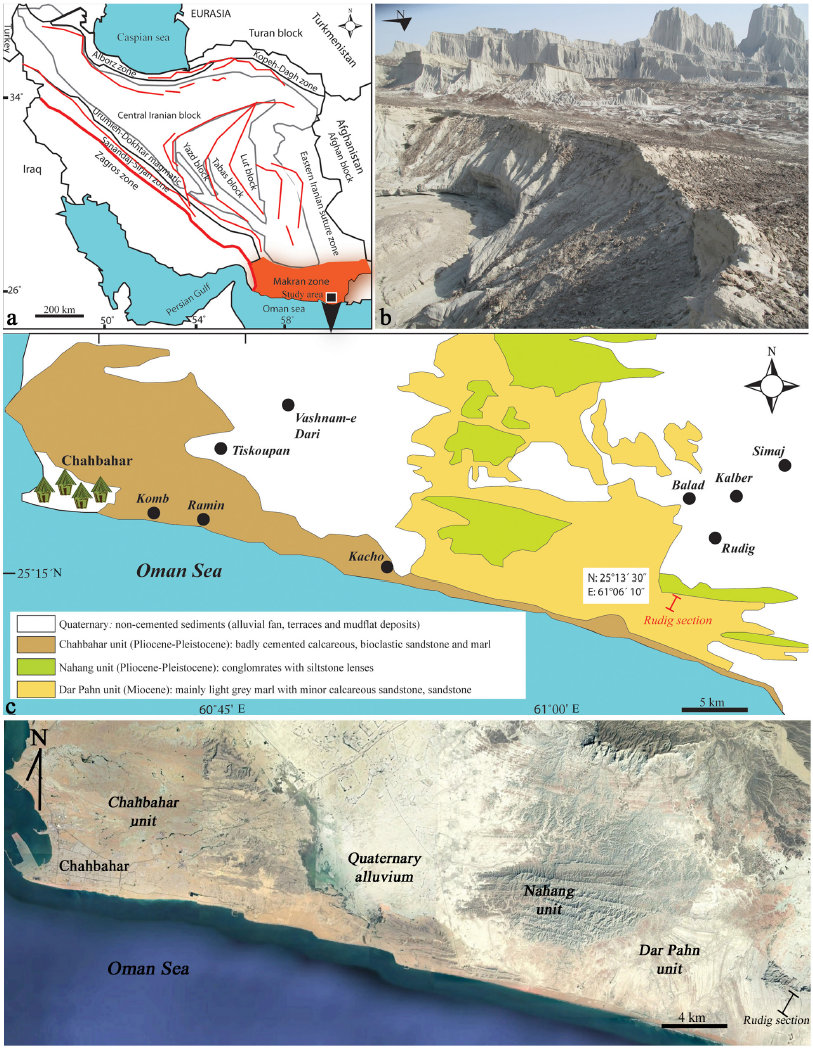
Figure 1 Sketch map of Iran showing the geological provinces (modified from Stöcklin, 1968). b) Outcrop view of the Dar Pahn Unit in the Rudig section. c) Geologic map of northern Iran with the Cenozoic lithostratigraphic units (modified after Dolati, 2010). d) Satellite image of location of Rudig section, east of Chabahar, taken from Google Earth Pro.
The tectonic and structural styles of Makran accretionary complex have developed throughout the Cenozoic at the convergent in the result of the subduction of the oceanic part of the Arabian plate beneath Eurasia (Lut and Afghan blocks) (e.g. Byrne et al., 1992; Kopp et al., 2000; Haghipour, 2014). Makran is still less known area, difficult to access region that has therefore been the object of relatively few modern studies (Burg, 2018). Moreover, this zone contains four main provinces with different sedimentary and igneous associations, which reflect different stages in the evolution of the accretionary wedge (Dolati, 2010; Burg et al., 2013) that includes (1) North Makran area, (2) Inner Makran, (3) Outer Makran, and (4) Coastal Makran. The Rudig section here under study is located about 6 km south of Rudig village, 50 km east of Chabahar (Figure 1c). This section with total thickness of 100 m is composed of different lithological units of sandstone, silty marl, marly limestone and siltstone that are considered as "Dar Pahn" Unit (McCall, 1985) within the Coastal Makran area (Figure 1a, b, c, d). In previous studies, the deposits of the Dar Pahn Unit are attributed to the Middle-Upper Miocene to Pliocene timespan by McCall (1985) and Dolati (2010) (Figure 2). The lower stratigraphic boundary of the Unit is cut-off by the Chah Khan Thrust in the northern areas (approximately 70-80 km north of Rudig village), while the marl-dominated member is covered unconformably by the continental deposits of the Pliocene-Pleistocene Nahang Unit (Dolati, 2010) (Figure 1c).

3. Materials and Method
This study relies on 27 rock samples and 371 free specimens that were collected with 2-3 m sampling intervals composed of marl, silty marl, siltstone and sandstone beds containing abundant well-preserved mollusks specimens.
The calcareous nannofossil assemblages analyzed in the 26 slides from 13 samples that were prepared by the simple smear slide techniques (Bown and Young, 1998; modified by Parandavar and Hadavi, 2019). Slides were studied by using standard light microscope technique (Olympus BX53) in parallel-polarised (PPL) and cross-polarised (XPL) light with adding the Quartz (QP) and Gypsum plates (GP) at a magnification of 1250-2000x. In the current study, calcareous nannofossils were determined using taxonomic concepts of various authors (Theodoridis, 1984; Aubry, 1984; Perch-Nielsen, 1985, Bown, 1998). According to the biozones classification and age establishment, the low-latitude zonation schemes of Martini (1971; NN zones) and Backman et al. (2012; CNN zones) are followed here. Specimens are deposited at the paleontological collection of the Geology Department of Zanjan University, Iran, under acronym GDZU.
4. Biostratigraphy
Calcareous nannofossils of Late Miocene are diverse (Young, 1998) and provide high resolution biostratigraphy. Neogene nannofossil biozonation have been used by some specialists to define Miocene ages (Gartner, 1969; Martini, 1971; Okada and Bukry, 1980). Backman et al.(2012) worked with high resolution zonation scheme (CNM and CNPL zones) for low to mid latitude areas. Based on the scheme by Backman et al.(2012), the currently studied interval spans Chrons C5n to C4n, confined to the Tortonian stage. In terms of calcareous nannofossils, these zones are included in the NN9 to NN11 biozones (Figure 2). The calcareous nannofossil assemblages show moderate to good preservation and older reworked specimens are also found. Although limited etching, dissolution and calcite overgrowth occurred on large size specimens of Coccolithus, Helicosphaera, Discoaster, Reticulofenestra, all are identifiable. Stratigraphically important species from the studied section are shown in Figure 3.

Figure 3 Index and commonly identified calcareous nannofossils from the Rudig section (Dar Pahn Unit) (scale bar is 2 pm in all images). a-b) Coccolithus pelagicus with Bar (Ml; fig. a: XPL, fig. b: GP); c-d) Ceratolithus sp. (M2; fig. c: XPL, fig. d: QP); e-f) Calcidiscus leptoporus (M6; fig. e: XPL, fig. f: GP); g-h) Helicosphaera stalis (M6; fig. g: XPL, fig. h: GP); i-l) Sphenolithus abies (M7; figs. i&k: XPL, figs. j&l: GP, figs. i&j: 0°, figs. k&l:45°); m) Discoaster cf. calcaris (M7; QP); n) Discoaster bollii (M7; QP); o-p) Discoaster hamatus (M7; figs. o&p: QP); q-r) Reticulofenestra rotaria (M20; fig. q: XPL, fig. r: GP); s) Pontosphaera multipora (M20; XPL); t) Discoaster variabilis (M20; QP); u-w) Helicosphaera princei (figs. u&v: M26; fig. w: M27; figs. u&w: XPL, fig. v: GP; fig. w:45°); t) Discoaster berggrenii (M.26; QP). Abbreviations: M= Sample No.; XPL= Cross Polarizing Light; GP= Gypsum Plate; QP= Quartz Plate.
5. Systematic palaeontology
| Class Gastropoda Subclass Caenogastropoda Order (Caenogastropoda) Superfamily Cerithioidea Family Turritellidae Lovén, 1847 |
Description. Except for the terebridae, this family is known as turriform sea slugs. Slender for its tall, conic and stretch sculpture. With 10 to 12 spiral rounds a little convex, and a smooth suture that separates it. The profile of the turn is straight and sharp, generally with a weak axial sculpture. The aperture tends to be oval or semi round and occupies the thick of the spire's body, thick external lip. Horny operculum. They dwell from the lowest line of the tide, in a calcareous sandy substratum with relatively shallow, in a 10 m deep normally. They are more commonly buried in sandy and muddy bottoms, in warm and tropical waters (Abbott and Dance, 1983).
| Subfamily Turritellinae Genus Turritella Lamarck, 1799 Turritella sp. Figure 4.1a, b |

Figure 4 Gastropods from the Rudig section, Upper Miocene Dar Pahn Unit. 1a, b) Turritella sp. a, apertural view; b, adapertural view. 2a, b) Thylacodes sp. 3a, b) Polineces sp. a, apertural view; b, adapertural. 4) Tibia sp., lateral view. 5a, b) Bursa sp. a, apertural view; b, adapertural view. 6a, b) Bullia sp. a, apertural view; b, adapertural view.
Material. 53 specimens, GDZU-0001 to GDZU-0053.
Description. Thick and robust shell, coiled and stretched in a spiral with various round turns and spire rings from faint to prominent. The colors are variated with irregular stains. They inhabit in colonies, buried in sandy to muddy bottoms from the infralittoral zone to 80 m deep. Mainly distribution on high latitudes, whereas in tropical latitudes are not so common (Abbott and Dance, 1983).
| Order Littorinimorpha Superfamily Vermetoidea Family Vermetidae Rafinesque, 1815 |
Description. Shells in worm, tubular and stretched forms, very irregular in its growing. With its spire more shut when the individual is young. It adheres to rocks and forms loosed and grouped tubes, in occasions, two or more individuals merges into forming matted like masses. The irregular growth and its adaptation to the substratum allows it to have a vast morphological plasticity on the adults. These gastropods are one of the hardest to find complete. The Operculum is made of chitin.
| Genus Thylacodes Guettard, 1770 Thylacodes sp. Figure 4.2a, b |
Material. 11 specimens, GDZU-0054 to GDZU-0064.
Description. The shells of this genus are somewhat stretched and are only attached to the rock in its juvenile state. With shell subcylindrical, twisted, inner side tends to be flat, and the external surface it is composed of tiny longitudinal stretch marks; each one can note of a single line of granulates. The continuity of the internal tube is interrupted by oblique diaphragms. Sometimes the form is almost spiral, and a specimen has three whole round turns very irregular. The mollusk of this genus resembles of big tubes, and the rollup has a form of a flat spiral. It is difficult to determinate the species of this group because of (i) the base of the juveniles (ii) an external structure almost cleanand (iii) the contour of the aperture and the operculum. Species like Thylacodes squamigerus are distributed all along the north Pacific Ocean coasts (Abbott and Dance, 1983).
| Superfamily Naticoidea Family Naticidae Guilding, 1834 |
Description. Small to medium solid shell, with a tendency of oval shape, smooth and shiny surface. With the turn short, the body of the primary spire is big; the aperture is from semicircular to oval, angular in the superior part and rounded in the inferior. The umbilicus normally has a big and thick callus. It characterizes the presence of a funiculus in the umbilicus (closed or open). With corneal operculum. An extensively distributed family around the world (Abbott and Dance, 1983).
| Subfamily Polinicinae Genus Polinices Montfort, 1810 Polinices sp. Figure 4.3a, b |
Material. 16 specimens, GDZU-0065 to GDZU-0080.
Description. Consistent shell, from concave to oval, generally low, with the apex curved and bulky. The external sculpture is almost smooth with a shiny coloring. Even though it is generally found eroded, it is possible to observe tiny and thin radial lines from brownish to more dark colors, more notorious to the border, it is also possible to appreciate fine lines of concentric growing; the umbilicus does not have a real funiculus, so in which case, the major part of the callus is in the superior side of the internal lip. It shows a weak spiral cord that forms an angle, giving an origin to a shoulder less notorious, which could have a more clarified coloring than the rest of the round turn. The last turn occupies almost the totality of the shell. The aperture is big, with a "D" form; external lip thin and smooth, which could be easily eroded on empty shells. The umbilicus aperture is slightly closed by a thick callus, which is thicker in the parietal zone. They live in low tide zones and have operculum. They are temperate and tropical species. Nowadays, some species of the Polinices genus can be found commonly in the southern Atlantic coastlines (Abbott and Dance, 1983).
| Superfamily Stromboidea Family Rostellariidae Gabb, 1868 |
Description. The shell is robust and solid, fusiform or in spindle forms, with a spire of numerous turns, a large aperture, and an external jagged margin. Small operculum. Subtropical and tropical distribution. They have wide distribution, primary from the Red Sea to the southeast Asian, reaching to the northwest of Australia and New Guinea (Abbott and Dance, 1983).
| Genus Tibia Röding, 1798 Tibia sp. Figure 4.4 |
Material. 1 specimen, GDZU-0081.
Description. Elongated shell and smooth surface, it projects with a conic, large and sharpened spire, elliptic aperture, very long syphon channel that projects like a big, elongated thorn. Jagged external lip. It dwells in shallow areas, in tropical seas. Many species are distributed in the Red Sea (Eisenberg, 1984).
| Superfamily Tonnoidea Family Bursidae Thiele, 1925 |
Description. Big heavy shell, lightly flattened on the back-ventral side, with strong ornaments by nodules and axial varicose veins with prominences that sometimes turns into thorns, robust and relative short spire angled. The posterior part of the aperture's posterior channel is oval and compressed sideways with a deep channel. The posterior channel is short, and the external lip is jagged. The operculum is made of chitin and is oval. Many species live in corals. They dwell in rocks and reefs of tropical seas. Small family that inhabits temperate and tropical waters (Eisenberg, 1984).
| Genus Bursa Röding, 1798 Bursa sp. Figure 4.5a, b |
Material. 89 specimens, GDZU-0082 to GDZU-0170.
Description. The shell lacks periostracum and the spire has primary two varicose veins. The aperture has a deep slot, with a deep and short syphon channel. The internal lip is jagged. Distributed through the Caribbean, Mediterranean, South Africa coastline, Western coast of Australia and the Yellow Sea (Eisenberg, 1984).
| Order Neogastropoda Superfamily Buccinoidea Family Nassariidae Iredale, 1916 (1835) |
Description. Shells from small to medium sizes, marine, of thick complex and with a parental callus well developed, also they have axillary edges. The color of the shells may vary from dark gray to white, with dark brownish bands. Some specimens show numerous spire chords. They feed on carrion and live buried underneath the sand or mud ((Abbott and Dance, 1983).
| Subfamily Bulliinae Genus Bullia Gray, 1834 in Griffith & Pidgeon Bullia sp. Figure 4.6a, b |
Material. 1 specimen, GDZU-0171.
Description. This genus was erected by Gray (Griffith, 1834) with the purpose of distinguishing a group of species, which was thought to be in between Buccinum and Terebra; owing to poor knowledge of its anatomy, the taxonomic description of this genus is constraining. Fusiform shell, stretched with a proto-shell elongated, smooth, shiny, spire with a superfluous marked suture, absent umbilicus, oval aperture, simple external lip, sharpened, rounded. Simple internal lip and narrow callus. The columella possesses strong folds in a spiral, located between the middle and inferior thirds of each whorl. They live in sandy bottoms. Distributed in the Mediterranean sea and the northwest coasts of Africa (Eisenberg, 1984).
| Superfamily Mitroidea Family Mitridae Swainson, 1831 |
Description. Shells are thick, elongated, oval or fusiforms; from small to large sizes. The spire is high and sharply pointed. It can show smooth or notorious spiral grooves and axial ribs. Colors may vary, some very colorful with or without stains. While other can be pale colors. The aperture is elongated and narrow. The outer lip is smooth, and the columella has three or more prominent folds. The periostracum tends to be thin, and the siphonal notch to be short. They present a venomous gland. The distribution tends to be wide in temperate to tropical waters, but mainly in the Indo-Pacific (Abbott and Dance, 1983). They dwell in sand, seaweed, or coral environments, some buried or underneath rocks. Not only can be found mainly in the intertidal zone, but also can reach deep waters (Abbott and Dance, 1983).
| Subfamily Mitrinae Genus Mitra Lamarck, 1798 Mitra sp. Figure 5a, b |

Figure 5 Gastropods and bivalve from the Rudig section, Upper Miocene Dar Pahn Unit. 1a, b) Mitra sp. a, apertural view; b, adapertural view. 2a-c) Conus sp. a, apertural view; b, adapertural view; c, spiral view. 3a, b) Leucosyrinx sp. a, apertural view; b, adapertural view. 4a, b) Architectonica sp. a, adapertural view; b, apertural view. 5a-c) Anadara sp. a, interior of right valve; b, exterior of right valve; c, dorsal view.
Material. 4 specimens, GDZU-0172 to GDZU-0175.
Description. Thick and fusiform shell, with an elongated spire and sharp at the apex. they have a small and narrow aperture, and an oblique and folded columella. Colors vary from specie to specie, while some can be very colorful with stains and bands, others can be pale colors. They can be found in temperate to tropical waters, buried underneath sand or in coral reefs; in seas like the Indo-Pacific, East of Africa, Red Sea and Japan's coastlines (Eisenberg, 1984).
| Subclass Orthogastropodaa Superorder Caenogastropoda Order/cladus Neogastropoda Superfamily Conoidea Family Conidae J. Fleming, 1822 |
Description. The shells characterize, as its name suggests, for having the shape of a cone. They do not reach big sizes. The shell is heavy and thick, broad in the superior part and the last turn forms the body, which reduces gradually in a narrow base. The colors may vary from very pallid to very colorful. The spire has a turret form and it is ornamented by spiral nodules in the last turn, to which many species have the shape of a crown. The tip of the apex is flat. Smooth ornamentation or with superficial grooves. The aperture is long, narrow that runs through all its length; external lip is thin and sharp, very susceptible on breaking.
The periostracum is dark and fibrous, which hidden the color of the shell. The operculum is corneous, narrow and elongated in the shape of a nail. They are primary located on tropical areas (Eisenberg, 1984).
| Genus Conus Linnaeus, 1758 Conus sp. Figure 5.2a, b, c |
Material. 37 specimens, GDZU-0176 to GDZU-0212.
Description. Only genus in this family, being the color pattern or the design of the shell the characteristic diagnostic more used in the differentiation to species level. Associated with a poisonous. A very diverse group. Tropical and Subtropical distribution (Abbott and Dance, 1983).
| Family Pseudomelatomidae Morrison, 1966 |
Description. This group constitutes of a grand variety of the Conoidea, they are mostly denominated turret shells. The shells can be from small to big, in a fusiform shape. The majority of the species have an elongated sculpture with a conspicuous ornate in spirals or in axial shapes, and a subsuture fold, with a proto shell normally paucispiral; but it can be multispiral. The anal chord is well developed. They dwell in tropical and subtropical waters on a sandy substrate, in a vast range of depths (Abbott and Dance, 1983).
| Genus Leucosyrinx Dall, 1889 Leucosyrinx sp. Figure 5.3a, b |
Material. 57 specimens, GDZU-0213 to GDZU-0269.
Description. Shell from small to medium, thin, fusiform with high spire, small proto shell, two smooth spires. In adults, with axial nodules in the periphery and with spiral chords. extremely deep anal sinus, largely concave, external lip in bow shape. They can attain more than 1000 m of depth. Widely distributed in South America coastlines (Eisenberg, 1984).
| Subclass Heterobranchia Infraclass Heterobranchia Superfamily Architectonicoidea Family Architectonicidae Gray, 1850 |
Description. Snails that are called sun watches, with solid shell, generally discoidal, conic, with low spire, trochoid shape, strongly spiraled, ornamentations with nodule chords in a rosary shape in spiral, deep wide circular umbilicus surrounded by a thick chord of nodules and/or outgoing helicoidal like a snail stairs shape. With a corneal operculum. They show a wide distribution in the Panamic province, as to the Atlantic and Mediterranean regions (Abbott and Dance, 1983).
| Genus Architectonica Röding, 1798 Architectonica sp. Figure 5.4a, b |
Material. 2 specimens, GDZU-0270 and GDZU-0271.
Description. Medium size, spires folded strongly, the ornamentation with shiny enamel has many marbled stains underneath the sutures, that tends to vary between species. Belt with conspicuous nodules that border specifically the umbilicus. They dwell on tropical and temperate areas, with a wide distribution in the Indo-Pacific Ocean (Abbott and Dance, 1983).
| Class Bivalvia Subclass Autobranchia Infraclass Pteriomorphia Order Arcida Superfamily Arcoidea Family Arcidae Lamarck, 1809 |
Description. Shells are generally equivalves, triangle forms, quadrangular or oval, with a strong radial ornament. The hinge is more or less straight, with numerous teeth straight or in "V" form, umbus wide and rolled up, equivalent muscular trace, and paleal line continuous. Shells coated by a periostracum, which can be shown as fibrous or downy (Abbott and Dance, 1983).
| Genus Anadara Gray, 1847 Anadara sp. Figure 5.5a, b, c |
Material. 19 specimens, GDZU-0272 to GDZU-0240.
Thick shell, heavy, from medium to relatively big, oval oblong form, with strong radial sculpture, grooves between the ribs are generally narrow and deep, teeth are more or less the same size. Crenulations in the margin of the shell. Without a basal aperture. They are tropical and subtropical. Grand and main distribution in the Mediterranean Sea (Eisenberg, 1984).
| Order Ostreida Superfamily Ostreoidea Family Ostreidae Rafinesque, 1815 |
Description. Left (inferior) valve is big and deep; the right (superior) valve is almost flat; the hinge line with or without teeth; the margin of the hinge is or not jagged or comata that can extend all along the border of both valves depending on the species. Ribs are radial and irregular that tends to divide on the growth (Abbott and Dance, 1983).
| Subfamily Ostreinae Genus Ostrea Linnaeus, 1758 Ostrea sp. Figure 6.1a, b |

Figure 6 Bivalves from the Rudig section, Upper Miocene Dar Pahn Unit. 1a, b) Ostrea sp. a, interior of right valve; b, exterior of right valve. 2a, b) Pecten sp. a, interior of left valve, b, exterior of left valve. 3a, b) Anomia sp. a, interior of right valve, b, exterior of right valve. 4a, b) Dosinia sp. a, exterior of right valve; b, dorsal view. 5a, b) Pophia sp. a, interior of left valve; b, exterior of left valve. 6a, b) Tagelus sp. a, interior of right valve; b, exterior of right valve. 7a-c) Corbula sp. a, interior of right valve; b, exterior of right valve, c, dorsal view.
Material. 2 specimens, GDZU-0241 and GDZU-0242.
Description. Subcircular shell, with the valve shell relatively deep; the right or superior is flat. Muscular mark is subcentral, with or without teeth sideways of the hinge and alongside of the internal border of the valves. Margins are almost smooth without big folds. Their distribution is worldwide in all oceans (Eisenberg, 1984).
| Order Pectinida Superfamily Pectinoidea Family Pectinidae Rafinesque, 1815 |
Description. Shells not very big, equivalves or inequivalves, closely circular, or in fan shape, commonly channeled ornaments. Their auricles are almost the same, localized at the sides of the hinge. The fixation valve is bulky, with a notch or byssal cleft. They attach to hard objects with the byssus. The right superior valve has more shine and/or coloring. They are found in almost all seas of the world (Eisenberg, 1984).
| Subfamily Pectininae Tribe Pectinini Genus Pecten O.F. Müller, 1776 Pecten sp. Figure 6.2a, b |
Material. 17 specimens, GDZU-0243 to GDZU-0260.
Description. The right valve is generally more strongly arcuated than the left valve, which goes from flat to concave. The "ears" to the sides of the resilium of the hinge are almost the same in size and shape. Ornamentation with radial ribs. Wide distribution, but high concentrations in the European Atlantic coasts, and the Mediterranean Sea (Eisenberg, 1984).
| Superfamily Anomioidea Family Anomiidae Rafinesque, 1815 |
Description. Irregular Shell in its margins, thin and fragile, with relative transparency, maintaining a pearly shine in the inner side. They are inequivalves, right valve has an orifice near the hinge, in which a calcareous byssus can be projected, which can be attached to corals or rocks. It has three muscular prints over the central area of the right valve. The hinge is lack jagged. They are principally sessile (Abbott and Dance, 1983).
| Genus Anomia Linnaeus, 1758 Anomia sp. Figure 6.3a, b |
Material. 3 specimens, GDZU-0261 to GDZU-0263.
Description. The pattern of the muscular adductor mark is a guide for recognizing the genus. Translucid shell, shiny pearly in the interior. The right valve is concave and perforated. The left valve is convex with three muscular marks; one big and two little. They show a great worldwide distribution in all oceans (Eisenberg, 1984).
| Infraclass Heteroconchia Subterclass Euheterodonta Superorder Imparidentia Order Venerida Superfamily Veneroidea Family Veneridae Rafinesque, 1815 |
Description. A big family of variated equivalent shells, trigonal, oval or almost circular with a vast variety of external ornaments, from smooth to radial. Umbos generally to the prior half, producing normally a strong or soft curvature to the final prior part. External ligament. Lunula and escutcheon are normally well developed. Three cardinal teeth on each valve. Lateral posterior teeth are weak or absent, the previous lateral is present or absent. Variable paleal sinus, but present in a certain degree. Distribution in all seas (Eisenberg, 1984).
| Subfamily Dosiniinae Genus Dosinia Scopoli, 1777 Dosinia sp. Figure 6.4a, b |
Material. 13 specimens, GDZU-0264 to GDZU-0276.
Description. Solid shell of medium size, discoidal, inequilateral, equivalve, lunula well-defined, escutcheon defined or absent. Concentric grooves. Deep paleal sinus. Strong hinge, with a great similarity between species. Distribution in various seas around the world: Caribbean, European North Atlantic, and Indo-Pacific (Eisenberg, 1984).
| Subterclass Euheterodonta Superorder Imparidentia Order Venerida Superfamily Veneroidea Family Veneridae Rafinesque, 1815 |
Description. A grand family of variated equivalve shells, trigonal, oval or almost circular with a great variety of external ornamentations, from smooths to radials. Umbo is generally to the former half, commonly producing a strong or smooth curvature to the former final side. External ligament. Lunula and escutcheon normally are well developed. With three cardinal teeth on each valve. Weak posterior lateral tooth; or absent. Former lateral tooth also present or absent. Variable paleal sinus, but present at some degree. In almost all seas (Eisenberg, 1984).
| Subfamily Tapetinae Genus Paphia Röding, 1798 Pophia sp. Figure 6.5 a, b |
Material. 5 specimens. GDZU-0277 to GDZU-0281.
Description. Relatively small shell, solid, with a bean-like ventral contour, characterized by a plasticity of phenotype. They dwell in sediments, with wide geographic distribution. The clams of this genus inhabit in salty and oceanic waters. There are fossil records from the Cretaceous to the Quaternary. Distributed with great concentrations in the Mediterranean and North Atlantic seas (Eisenberg, 1984).
Description. Shells of medium size, elongated, quadrangular, with subcentral hinge, convex, equivalve and inequilateral. The umbo is orthogyrus. The valves are ajar by the sides. Hinge is narrow and thin. White color and yellow, the periostracum is brownish. They dwell buried in muddy bottoms, in the infra and circumlateral zones (Abbott and Dance, 1983).
| Genus Tagelus Gray, 1847 Tagelus sp. Figure 6.6 a, b |
Material. 7 specimens, GDZU-0282 to GDZU-0288.
Description. Shell is narrow, elongated with the dorsal and ventral borders almost parallel on one each other, with rounded borders. The external sculpture shows smooth concentric rounded lines. The valves have a whitish-violet colour. The periostracum is thing, brown yellowish to dark brown. The muscular marks are notorious. The umbo is small, almost centerical. The hinge has two cardinal teeth, the ones on the right valve are bigger and taller. On the left valve, the posterior cardinal is big and sharp. It is common to find them in bays up to 20m deep, buried underneath sandy substratum from the North Atlantic seas (Abbott and Dance, 1983).
| Order Myida Superfamily Myoidea Family Corbulidae Lamarck, 1818 |
Description. Small shell, thick, strongly and consistent, slightly inequivalve, inequilateral, subtrigonal, inflated with a weak development of teeth in the hinge. Internal ligament. The left valve has a spoon shape. Generally ornamented with strong concentric stretch marks. Paleal sinus is small or absent. They are found near the coast of all seas (Eisenberg, 1984).
| Subfamily Corbulinae Genus Corbula Bruguiére, 1797 Corbula sp. Figure 6.7a, b, c |
Material. 25 specimens, GDZU-0289 to GDZU-0313.
Description. Shell robust, oval and subtrigonal, with the umbo next to the mean line. The right valve is convex, it surrounds and flap the left and smaller one. Sculpture formed by concentric ribs and ridges, and some radial stretch marks on the left valve. Right valve with a simple tooth, prior to the chondrophore, and other simple and elongated posterior and prior. The left valve has a chondrophore like a slightly gap. The mark of the adductor is visible and paleal sinus is shallow or absent. They live buried underneath the sand. Indo-Pacific distribution, with great concentrations in the northern coasts of Australia (Abbott and Dance, 1983).
6. Discussion
The excellent preservation of the specimens, along with features such as multiple articulated bivalves, suggests that this assemblage suffered moderate to null transport from the place where they were buried (Figure 7). Although the forms might represent different species within the same genus, the uniform morphology observed for the representatives of the genera suggest that they could be placed in one species. The preservation observed in mollusks from older Cenozoic stratigraphic units of Iran show a range of details in completeness of the shall and ornamentation, but the morphology of the mollusks here reported represent one of the best preservation among fossil mollusks from Iran, suggesting a rapid burial and practically no transport. The presence of different sizes of specimens from the same genera indicates that the assemblage includes the diverse stages of the paleocommunity (Figure 8), and thus it might be possible to find microconchs and other microfossils such as ostracods and foraminifera. We are aware that it would be desirable to present a more detailed identification of the specimens reported here, but at the moment we consider the value of this report important in terms of an overview of the potential diversity of invertebrates, which may also include crustaceans and other invertebrate groups that lived in the shallow areas of the Late Miocene coast of SE Iran.

Figure 7 1) Coquina integrated by articulated Dosinia sp., indicating little to no transport from the burial place of the bivalves, pencil for scale. 2) Cardium sp. still articulated and some even closed, notice no damage on delicate radial ribs of shells, pencil for scale. 3) Several complete delicate shells of Turritella sp., and one nearly complete Bursa sp., finger and pencil for scale. 4) Nearly complete specimens of the vermetid, mucous feeder gastropod Thylacodes sp., pencil for scale. 5) Complete shell of Tibia sp., preserving delicate spire but specially thin siphonal channel, marker for scale.

Figure 8 1, 2) Different stages of Turritella sp., possibly at least two different species. 3, 4) Different preservation and stages of Bursa sp. 5) Different stages of Conus sp. 6) Dorsal view of articulated Dosinia sp. 7, 8) Different stages of Pecten sp. 9) Different stages of articulated Dosinia sp.
7. Conclusion
A total of 313 mollusk specimens from the Dar Pahn Unit (Upper Miocene, Tortonian) are represented by ten gastropod and eight bivalve genera. The detailed calcareous nannofossil biostratigraphy indicates a Late Miocene (Tortonian) age for this diverse mollusk assemblage, whose excellent preservation was probably due to little or no transport and a rapid burial. Most of the species suggest an infaunal habit which also helped in their preservation, supported by many articulated bivalves and fine fragile gastropod structures, such as the long and delicate siphonal channel of Tibia sp. All the reported mollusk assemblage comes from one single outcrop sequence (Rudig section) (Figure 7). The potential of the Dar Pahn Unit to contain more mollusk species as well as other littoral invertebrates such as crustaceans must be explored, since this might be one of the most productive and diverse fossil assemblages of the Cenozoic from Iran.
Contributions of authors
Mehdi Hadi fieldwork and sampling, conceptualization, investigation, writing original draft. Majid Mirzaie Ataabadi fieldwork and sampling, investigation. Jafar Sharifi-investigation. Mohammad Parandavar-conceptualization, investigation, writing original draft. Zoila Graciela Castillo-Rodríguez conceptualization, investigation, writing original draft. Francisco A. Vega-Sandoval conceptualization and investigation. Francisco J. Vega conceptualization, methodology, investigation, writing original draft and supervision of manuscript structure.
Financing
This research was developed with the own resources of authors.
Conflict of interest
We declare no conflict of interest.
Handling editor
Alessandro Garassino.

