











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El género Colubrina Rich. ex Brong, perteneciente a la familia Rhamnaceae Juss., comprende aproximadamente 30 especies distribuidas en diversas regiones tropicales y subtropicales, con una notable presencia en México donde se concentra el 70% de los taxones conocidos. La distribución abarca también otras áreas de América, Asia y Australia, manifestando una adaptabilidad ecológica que le permite colonizar desde matorrales xerófilos hasta bosques de pino y encinos (Fernández-Nava, 1993 p.90). La variabilidad morfológica y ecológica del género son retos para entender la taxonomía y evolución del mismo, especialmente en el contexto donde la delimitación de especies y subgéneros sigue siendo un desafío. La familia Rhamnaceae ha sido estudiada desde el punto de vista taxonómico-florístico como lo muestran las investigaciones de Standley (1923); Johnston., (1963, 1969 y 1971); Wiggins (1964, 1980); Martínez, (1979), (Fernández_Nava (1993, 1996, 2010), (Pool ( 2015) y (Nesom, 2013, 2023a, 2023b).
En el ámbito de la anatomía y arquitectura foliar de la familia, destacan los trabajos de Meyer y Meola (1978); Schirarend (1991); Colares y Arambarri (2008); Shisode y Patil (2011); y Hernández-Peñaloza et al. (2021), quienes han aportado información valiosa para la comprensión de la variabilidad estructural de varios géneros de Rhamnaceae. Así también, Fernández-Nava, et al. (2024) caracterizaron algunas especies de Colubrina sección Serrataria a través de la arquitectura foliar y superficies epidérmicas como apoyo a la taxonomía del grupo. En cuanto a la parte etnobotánica de Colubrina se destacan en Fernandez-Nava (2010).
En la historia de la taxonomía de Colubrina, los géneros afines son Cormonema Reissek ex Endlicher e Hybosperma Urb., que han sido objeto de debate y finalmente Johnston (1971, p. 7), los incorporó al género Colubrina, este mismo autor realizó el estudio monográfico que identificó dos subgéneros: Colubrina, que incluyó las secciones Barcena, Cowania, Colubrina y Capuronia (esta última no presente en México) y Serrataria que no se subdivide en secciones.
Esta clasificación, aunque ampliamente aceptada, en los últimos años sugiere la necesidad de reevaluar los caracteres utilizados para distinguir los taxones dentro del género (Fernández-Nava et al., 2024). En este contexto, la presente investigación se enfocó en la arquitectura foliar y las superficies epidérmicas de las láminas foliares como herramientas para la diferenciación taxonómica de Colubrina. El estudio se realizó en nueve especies mexicanas, cuyas características morfológicas fueron comparadas con los otros siete taxones previamente analizados por Fernández-Nava et al. (2024). El propósito de este trabajo fue identificar caracteres distintivos que permitan una mejor delimitación de los taxones y evaluar la validez de la circunscripción actual de los subgéneros y secciones propuestos por Johnston (1971).
Materiales y método
El material vegetal utilizado para el estudio se encuentra depositado en los Herbarios de la Escuela Nacional de Ciencias Biológicas del Instituto Politécnico Nacional (ENCB) y además se examinaron especímenes del Herbario del Instituto de Biología de la Universidad Nacional Autónoma de México (MEXU). La identificación de los taxones fue confirmada por el Dr. Rafael Fernández Nava, especialista en Rhamnaceae. Las muestras de las exsicatas estudiadas fueron:
C. arborescens (Mill.) Sarg., C. Chan 6061 (ENCB); O. Téllez 1230 (ENCB); E. Ventura y E López 1654 (ENCB).
C. californica (Miller) Sarg., A.Carter, 4903 (MEXU).
C. celtidifolia Cham. & Schlecht., A. Goméz-Pompa 743 (MEXU); J. C. Soto Núñez 9371 (MEXU).
C. ehrenbergii Schlecth., S. Zamudio 2382 (ENCB); L. González-Quintero 3578 (ENCB); J. Rzewdoski 5379 (ENCB).
C. elliptica (Swartz) Brizicky & Stern. L. González 2556 (ENCB); D. E. Beedlove 1404 (ENCB); H. Puig 2917(ENCB).
C. heteroneura (Griseb.) Standley, W. R. Anderson 12712 (ENCB); C. Davidson 7650 (ENCB); F. Ventura A. 15684 (ENCB).
C. johnstonii Wendt, C. Chan, 6061. (ENCB); O. Téllez, 1230 (ENCB); T. Wendt, 2535. (ENCB).
C. triflora Brongn., V.W. Steiiman 1877 (ENCB); S. Zamudio y E. Pérez 7558 (ENCB).
C. viridis (M. E. Jones) M. C. Johnston, J. M. Alcocer s.n. (ENCB); Pinkavaet al. 21674 (ENCB); C. L. Díaz 2251 (ENCB).
De cada espécimen se tomaron cuatro hojas plenamente desarrolladas ubicadas entre el tercero y quinto nudo de las ramas montadas en las muestras de herbario y de preferencia, de tres ejemplares de cada especie. En algunos de estos taxones las hojas se encuentran pegadas a las cartulinas y no fue posible extraer muestras o bien, ciertas especies están poco representadas en los herbarios y su distribución es muy específica, lo que explica la poca representación en las colecciones como C. californica y C. stricta de las que se tomaron una muestra y con C. celtidifolia, fueron dos especímenes. Una vez que se obtuvieron las muestras para la observación de la arquitectura y anatomía foliar de las especies mexicanas de Colubrina se utilizaron dos técnicas para transparentar las láminas foliares, la de Aguirre-Claverán y Arreguín-Sánchez (1988, p. 11) y la de Sheffy y Potter (1969) tomada de Dilcher (1974, p.60), las cuales se describen con detalle en Fernández-Nava et al. (2024).
En el caso de Colubrina arborescens que presentaron glóbulos en la superficie foliar que se apreciaron en el material diafanizado aun sin teñir en el microscopio óptico, en estas preparaciones se colocaron dos gotas de Sudán IV y se dejó actuar por 15 minutos, los glóbulos se colorearon debido a que este reactivo se caracteriza por teñir en tonalidades de rojo los lípidos.
La densidad de los tricomas en todos los taxones se consideró contando el número de los mismos en un 1 mm2 y la densidad estomática se calculó según (Colares y Arambarri, 2008. p. 570). La modificación de la base de los tricomas se explica e ilustra en (Fernández-Nava, 2024 p. 7). En las medidas del largo de los pelos no se incluyen las células epidérmicas modificadas en caso de presentarse.
Para las descripciones de la arquitectura foliar y de las superficies epidérmicas de la lámina se tomaron en consideración los trabajos de Hickey (1973, 1974), Dilcher (1974), (Hickey y Wolfe (1975) y Ash et al. (1999). Para la epidermis foliar se consideraron los criterios de Gola, Negri, y Cappelletti (1965); Dilcher (1974); Cortés (1982) y Azcárraga et al. (2010). Los caracteres que se tomaron en consideración se muestran en la Tabla 1.
Tabla 1 Caracteres exomorfológicos de arquitectura foliar y de la epidermis foliar.Table 1. Exomorphological characteristics of leaf architecture and leaf epidermis.
| Caracteres | Descripción |
|---|---|
| Caracteres morfológicos y de arquitectura foliar |
Tipos de hojas: simples o compuestas Simetría Forma de la lámina foliar Medidas de largo y ancho de la lámina foliar. Ápice Base Textura Margen Presencia o ausencia de glándulas Largo del peciolo Tipo de arquitectura foliar Venas de primera, segunda, tercera y cuarta categoría. |
| Caracteres de Epidermis Foliar |
Tipos de tricomas: glandulares, eglandulares Largo de los tricomas Base de tricomas: con o sin base radial de células epidérmicas Densidad de los tricomas por mm2 Forma de células epidérmicas (superficie adaxial y abaxial) Tipos de paredes (contorno): rectas, onduladas Tipos de estomas. Tamaño de los estomas longitud y ancho Densidad estomática |
Para estas descripciones se tomaron como base los trabajos de Dilcher (1974); Ash et al. (1999); Johnston (1971); Wendt (1983); Fernández-Nava (1993). El dato de la textura de la lámina foliar se tomó de las descripciones de las especies y en algunas etiquetas de herbario se cita esta información, además, al realizar la diafanización de las láminas foliares de consistencia membranácea tardan menor tiempo en transparentarse con el hipoclorito de sodio al 30%, en algunas solo basta incluirlas en el reactivo para que se aclaren en seguida. Las hojas coriáceas o subcoriáceas tardan algunos minutos en transparentarse, así que el tiempo es variable y la técnica indica que una vez que se aclaren las láminas foliares, hay que sacarlas y colocarlas en agua sin especificar un tiempo determinado. Para las observaciones de la arquitectuta foliar se examinaron bajo el microscopio estereoscópico American Optical y las superficies epidérmicas en el microscopio óptico binocular Zeiss. La toma de fotomicrografías se llevó a cabo con una cámara digital Samsung Hmx-e10 con 8 megapixeles.
Se elaboró una base de datos en Excel para obtener la matriz de los caracteres de las especies para realizar el análisis de agrupamiento por el método UPGMA, utilizando el índice de similitud de Jaccard a través del programa estadístico Past 4.3 (Hammer et al., 2001) y el posterior dendograma. Se colocaron en las filas las especies y en las columnas los caracteres.
Resultados
Se describió la arquitectura foliar y las superficies epidérmicas de nueve especies de Colubrina que se organizaron en orden alfabético de las especies.
1.- Colubrina arborescens (Mill.) Sarg. (Lámina 1).
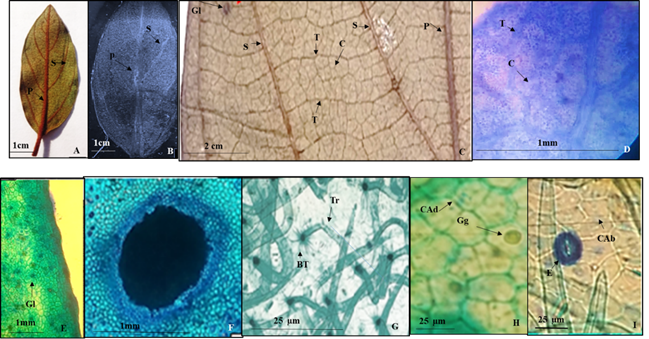
Lámina 1 Colubrina arborescens. A.- Lámina foliar del herbario. B.- Lámina foliar diafanizada con vena de primera y las de segunda categoría. C.- Vena de primera, segunda, tercera y cuarta categoría. D.- Detalle de venas reticuladas de tercera y cuarta jerarquía. F.- Glándulas en la superficie abaxial de la lámina foliar. F.- Detalle de una glándula. G.- Tricomas con base radial de células epidérmicas modificadas. H.- Células de la superficie adaxial isodiamétricas con paredes rectas con gotas de lípidos comprobado con Sudan IV (Gg). I.- Células epidérmicas de la superficie abaxial isodiamétricas con paredes rectas y estomas anomocíticos. Referencias: P vena de primera categoría. S venas de segunda categoría. T venas de tercera categoría. C venas de cuarta categoría. Tr tricoma. BT base del tricoma. Gl glándula. Gg glóbulo de grasa. CAd células epidérmicas de la superficie adaxial. CAb células epidérmicas de la superficie abaxial. E estoma
Hojas simples, simétricas, ovado-oblongas a oblongo-obovado u oblongo de 4 a 15 cm de largo a 2 a 9 cm de ancho, ápice agudo a corto acuminado, base acuminada a redondeada o cordiforme, lámina membranácea a subcoriácea, margen entero (Fig. 1A, 1B), glándulas circulares u ovadas, oscuras distribuidas en la superficie abaxial (Figs.1E, 1F). Peciolo de 5 a 30 mm de largo y 1 a 2 mm de diámetro (Fig. 1A). Vena primaria pinnada de curso recto (Figs. 1A, 1B) las de segunda categoría craspedódromas con 3 a 4 venas libres de cada lado y con el par de venas inferiores no ramificadas (Figs. 1A, 1B), las terciarias opuestas percurrentes (Fig. 1C) y las de cuarta categoría reticuladas sin venas libres incluidas (Figs. 1C, 1D).
Epidermis en vista superficial. Células epidérmicas adaxiales y abaxiales isodiamétricas de forma pentagonal con paredes de contorno recto (1H, 1I), con glóbulos de lípidos que se detectaron con sudán IV que es un colorante que pone de manifiesto la presencia de grasas (Fig. 1H). Estomas anomocíticos, en la superficie de la epidermis adaxial de 29 a 24 µm de largo y ancho respectivamente, densidad estomática de 26 estomas/mm2 en promedio, los estomas en la superficie abaxial de 26.1 a 17.3 µm de largo y ancho, densidad estomática en promedio de 26 estomas/mm2 (Fig. 1I). Tricomas unicelulares eglandulares, en promedio de 25.3 µm de largo, base radial de células epidérmicas modificadas (Fig. 1G) distribuidos en toda la superficie, aunque predominan en la vena central, la densidad promedio fue de 21 tricomas/mm2
2.- Colubrina californica I. M. Johnston (Lámina 2).

Lámina 2 Colubrina californica. A.- Lámina foliar del ejemplar de herbario. B.- Lámina foliar diafanizada mostrando las vena de primera categoría pinnada y el par inferior de venas secundarias con ramificaciones libres. C.- Arco de venas secundarias. D.- Arcos apicales. E.- Venas de tercera categoría opuestas percurrentes, las de cuarta categoría reticuladas sin venas libres incluidas. F.- Tricomas con base radial de células epidérmicas modificadas y células de la superficie adaxial. G.- Detalle de las células epidérmicas abaxiales (CAb). H.- Base de un tricoma con células epidérmicas modificadas. I.- Estomas anomocíticos. Referencias: P vena de primera categoría. S vena de segunda categoría. T vena de tercera categoría. C vena de cuarta categoría. Arcos (Ar). RS ramificación de venas de segunda categoría. Tr tricoma. BT base del tricoma. CAd células de la superficie adaxial. CAb células de la superficie abaxial. E estoma.
Hojas simples, simétricas, oblongas a obovadas a veces ovado-oblongas, de 1.2 a 3 cm de largo y 0.5 a 1.5 cm de ancho, ápice redondeado a emarginado, base redondeada a anchamente cuneada, con textura membranácea a subcoriácea, margen entero o con 2 a 6 pequeños dientes a cada lado distribuidos de la mitad de la lámina foliar hacia el ápice, sin glándulas en la lámina foliar (Figs. 2A, 2B). Peciolo de 1 a 5 mm de largo y 1 a 2 mm de diámetro (Fig. 2A). Vena de primera categoría pinnada de curso recto, las de segunda categoría, broquidódroma formando 3 a 4 arcos en ambos lados de la superficie foliar, que no llegan al margen de la hoja (Figs. 2B, 2C), el par basal se ramifica (Fig. 2B) y el par de venas superiores apicales se enrollan formando dos arcos (Fig. 2B, 2D), las de tercera jegarquía alternas percurrentes (Fig. 2E), las de cuarta reticuladas, sin venas libres incluidas (Fig. 2E).
Epidermis en vista superficial. Células epidérmicas de la superficie adaxial y abaxial isodiamétricas, de forma pentagonal con paredes de contorno recto (Fig. 2F, 2G). Estomas anomocíticos, en la superficie de la epidermis adaxial de 28.4 x 21.1 µm de largo y ancho, con densidad estomática de 15 estomas/mm2 y en la abaxial de 28.4 x 21.1 µm, con densidad estomática de 25 estomas/mm2 (Fig. 2I). Tricomas adaxiales unicelulares eglandulares, con base radial de células epidérmicas modificadas, en promedio de 29.8 µm, de largo, densidad promedio de 129 tricomas/mm2, los abaxiales densos de 32 µm de largo con 314 tricomas/mm2, (Figs. 2F, 2H).
3.- Colubrina celtidifolia (Cham. & Schlecht.) Schlecht. (Lámina 3).
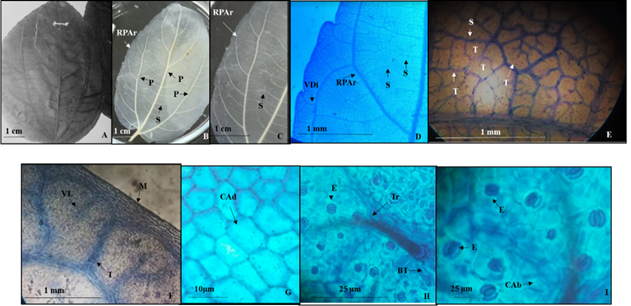
Lámina 3 Colubrina celtidifolia. A.- Lámina foliar del ejemplar de herbario. B.- Lámina foliar diafanizada mostrando las tres venas de primera categoría, las dos externas formando arcos. C.- Detalles de las ramificaciones formando arcos. D.- Arco de la ramificación de la vena primaria de donde deriva una vena libre que penetra al pequeño diente marginal. E.- Venas de segunda y tercera categoría, estas últimas sin venas o con venas libres ramificadas. F.- Venas de tercera jerarquía que llegan al margen con una o dos venas libres. G.- Células epidérmicas adaxiales isodiamétricas. H.- Tricoma con base radial de células epidérmicas modificadas y estomas. I.- Estomas anomociticos con células oclusivas engrosadas y células epidérmicas abaxiales (E). Referencias: P vena de primera categoría. S vena de segunda categoría. T vena de tercera categoría. RPAr ramificación del par de venas primarias externas que forman arcos. VDi vena libre del diente. VL vena libre. M. margen. Tr tricoma. BT base del tricoma. CAd células de la superficie adaxial. CAb células de la superficie abaxial. E estoma.
Hojas simples, simétricas, anchas a estrechamente ovadas a lanceoladas de 7 a 14 cm de largo y 3 a 8 cm de ancho, ápice corto acuminado, base redondeada a veces levemente cordada, textura membranácea (Figs. 3A, 3B), margen de la lámina foliar con 60 a 80 dientes diminutos sin glándulas (Fig. 3D). Peciolo de 7 a 15 mm de largo y 1 a 2 mm de diámetro. Venas de primera categoría acródroma basal imperfecta, con tres venas (Fig. 3B), las dos nervaduras externas se ramifican formando arcos (Figs. 3B, 3C) y en el margen se diferencia una vena que alimenta a cada diente (Fig. 3D) las de segunda categoría cruzan entre las venas primarias anastomosándose (3C) y las de tercera poligonales reticuladas, con o sin venas libres ramificadas (Fig. 3E, 3F).
Epidermis en vista superficial. Células epidérmicas adaxiales isodiamétricas de forma pentagonal con paredes de contorno recto (Fig. 3G), las abaxiales isodiamétricas, poligonales irregulares con paredes de contorno recto (Fig. 3I). Estomas en la superficie adaxial ausentes, los abaxiales anomocíticos de 21.1 a 18 µm de largo y ancho respectivamente (Fig. 3I), densidad estomática de 28/mm2. Tricomas adaxiales y abaxiales unicelulares eglandulares con la base radiada de células epidérmicas modificadas de 14.7 µm de largo, con densidad de 4/mm2, los abaxiales de 16.8 µm de largo con densidad de 18/mm2 en ambos casos distribuidos en toda la superficie (Fig. 3H).
4. - Colubrina ehrenbergii Schlecth. (Lámina 4).
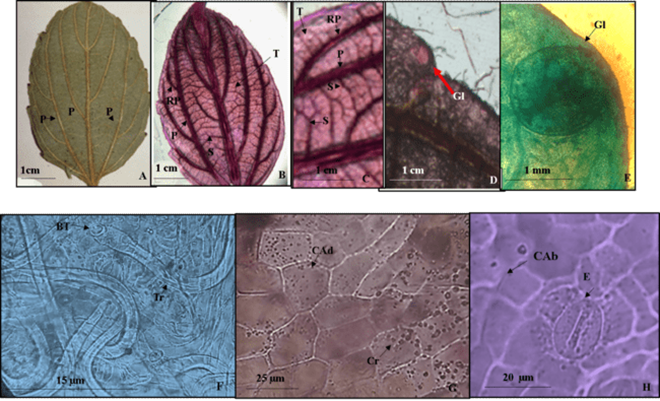
Lámina 4 Colubrina ehrenbergii. A.- Lámina foliar del ejemplar de herbario. B.- Material diafanizado mostrando tres venas de primera categoría ramificadas, las de segunda recorren trayectos predominantemente paralelos entre las venas de primarias y sus ramificaciones, las de tercera categoría libres ramificadas. C.- Detalle de venas de primera, segunda y tercera categoría, estas últimas ramificadas. D.- Diente con glándula marginal esférica. E.- Acercamiento de una glándula marginal. F.- Tricomas sin base de células epidérmicas modificadas. G.- Células adaxiales isodiamétricas con cristales circulares. H.- Células abaxiales isométricas y estoma anomocítico. Referencias: P vena de primera categoría. S vena de segunda categoría. T vena de tercera categoría. RP ramificaciones de venas primarias. Glándulas. BT base del tricoma. Tr tricoma. Cad células de la superficie adaxial. CAB células de la superficie abaxial. E estoma.
Hojas simples, simétricas ovadas a elíptico-ovadas de 3 a 8 cm de largo y 1.6 a 4 cm de ancho, ápice agudo a acuminado, base redondeada, textura coriácea a subcoriácea, margen dentado con 5 a 8 dientes por cada lado de la lámina foliar (4A, 4B) en el ápice de cada diente se aprecia una glándula circular de 1 a 2 mm de diámetro donde penetra una vena de segunda categoria (4D, 4E). Peciolo de 4 a 15 mm de largo y 1.5 a 2 mm de diámetro. Venas de primera categoría acródroma basal imperfecta con tres venas que se ramifican (Fig. 4B), las de segunda recorren trayectos predominantemente paralelos entre las venas primarias y sus ramificaciones, las de tercera categoría libres ramificadas (4C).
Epidermis en vista superficial. Células epidérmicas adaxiales y abaxiales isodiamétricas pentagonales con paredes de contorno recto, las primeras con cristales circulares (Figs. 4G, 4H). Estomas anomocíticos en la superficie adaxial de 21.1 y 16.8 µm de largo y ancho, densidad estomática promedio de 26 estomas/mm2, en la abaxial de 31.2 x 27 µm de largo y ancho, con densidad de 39 estomas/mm2. Tricomas adaxiales unicelulares eglandulares, sin células epidérmicas modificadas (Fig. 4F) de 25.3 µm de largo con 28 tricomas/mm2, se distribuyen en toda la superficie predominando en la vena primaria, tricomas abaxiales del mismo tipo de 25.3 µm de largo con densidad de 35 tricomas/mm2, predominan en todas las venas.
5.- Colubrina elliptica (Sw.) Brizicky & W.L. Stern. (Lámina 5).
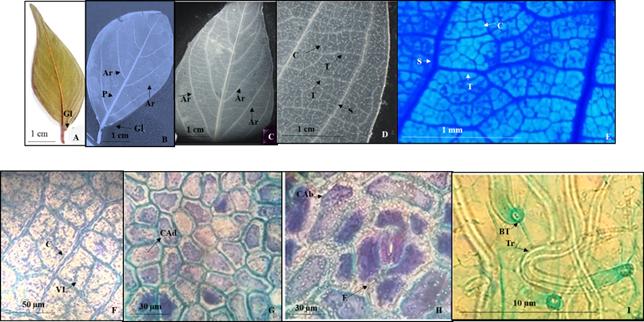
Lámina 5 Colubrina elliptica. A.- Lámina foliar del ejemplar de herbario. B.- Material diafanizado mostrando los arcos. C.- Detalle de los arcos que forman las venas secundarias. D.- Nervaduras de segunda, tercera y cuarta categoría. E.- Detalle de venaciones de segundo, tercero y cuarto nivel, las últimas con venas libres ramificadas o no. F.- Acercamiento de nervaduras de cuarta categoría con venas libres ramificadas y no. G.- Células epidérmicas adaxiales poligonales con paredes rectas. H.- Células epidérmicas poligonales abaxiales con paredes rectas y estoma anficiclocitico. I.- Tricomas unicelulares sin células epidérmicas modificadas. Referencias: P vena de primera categoría. S vena de segunda categoría. T vena de tercera categoría. Ar arcos de las venas secundarias. VL venas libres. Gl glándulas. Tr tricoma. BT base del tricoma. CAd células de la superficie adaxial. CAb células de la superficie abaxial. E estoma.
Hojas simples, simétricas, ovado-elíptica a veces ovadas y raras veces obovadas de 2.5 a 12 cm de largo y 1.5 a 5.3 cm de ancho, ápice agudo a acuminado, base redondeada a ampliamente cuneada, raramente redondeada (Figs. 5A, 5B) con dos glándulas que se presentan en la base de la lámina y en el margen (Figs. 5A, 5B), textura membranácea, margen entero. Peciolo de 5 a 25 mm de largo y de 0.5 a 1 mm de diámetro (Figs. 5A, 5B). Vena de primera categoría pinnada, las de segunda categoría broquidódroma, con 3 a 4 arcos que no llegan al margen, sin ramificarse (Fig. 5C), las de tercera categoría alternas percurrentes, las de cuarta con o sin venas ramificadas (Figs. 5D, 5E).
Epidermis en vista superficial. Células de la epidermis isodiamétricas adaxiales y abaxiales pentagonales y abaxiales cuadrangulares ambas paredes de contorno recto (Figs. 5G, 5H). Estomas de la superficie abaxial anficiclocíticos de 32.1 a 32.7 µm de largo y ancho con densidad estomática de 32 estomas/mm2. Tricomas ausentes en la superficie adaxial, en la abaxial unicelulares eglandulares sin células epidérmicas modificadas de 11.3 µm de largo y densidad de 32 tricomas/mm2, en toda la superficie aunque predominan en la vena central.
6.- Colubrina heteroneura (Griseb.) Standley (Lámina 6).
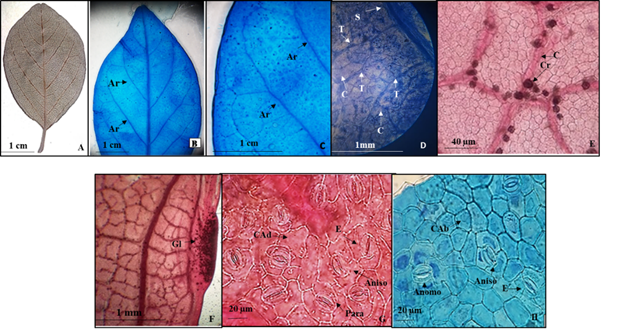
Lámina 6 Colubrina heteroneura. A.- Lámina foliar del ejemplar de herbario. B.- Lámina foliar diafanizada mostrando los arcos de las venas de segunda categoría. C.- Acercamiento de los arcos. D.- Detalle de las nervaduras de segunda, tercera y cuarta categoría, estas últimas sin venas incluidas. E.- Detalle de venas de cuarta categoría y cristales prismáticos. F.- Detalle de una glándula marginal. G.- Células epidérmicas adaxiales con paredes onduladas y estomas paracíticos y anisocítico H.- Células epidérmicas abaxiales isodiamétricas con paredes rectas y estomas anomociticos y anisocíticos. Referencias: P vena de primera categoría. S vena de segunda categoría. T vena de tercera categoría. C vena de cuarta categoría. Ar arcos de las venas secundarias. Gl glándula. CAd células de la superficie adaxial. CAb células de la superficie abaxial. E estoma Aniso anisocítico. Para paracítico. Anomo anomicítico.
Hojas simples, simétricas, obovadas a veces ovadas de 2.5 a 8.5 cm de largo y 1.7 a 6 cm de ancho, ápice agudo, levemente emarginado a redondeado, base redondeada a anchamente cuneada (Figs. 6A, 6B), textura membranácea, margen entero con escasas glándulas marginales presentes cercanas a la base de la lámina foliar, poco evidentes en los ejemplares de herbario de 1 a 1.5 mm de largo por 0.4 a 0.5 mm de ancho (Fig. 6F). Peciolo de 3 a 18 mm de largo y de 1.5 a 2 mm de diámetro (Fig. 6A). Vena de primera categoría pinnada (Figs. 6A, 6B) las de segundas broquidódromas formando cinco a seis arcos que no terminan en el margen (Figs. 6A, 6B, 6C) las de tercera alternas percurrentes y las de cuarta categoría reticuladas sin venas libres incluidas y con cristales (Figs. 6D, 6E).
Epidermis en vista superficial. Células de la epidermis adaxial con paredes de contorno ondulado (Fig. 6G), las abaxiales pentagonales isodiamétricas con paredes de contorno recto
(Fig. 6H). Estomas en la superficie adaxial anisocíticos y paracíticos (Fig. 6G) de 21.1 a 16.8 µm de largo y ancho con densidad estomática de 23 estomas/mm2, en la abaxial anisociticos y anomotetracíticos (Fig. 6H), de 25.3 a 21.7 µm de largo y ancho, con densidad de 31 estomas/mm2. Tricomas ausentes en ambas superficies de la lámina foliar.
7. - Colubrina johnstonii T.Wendt (Lámina 7).
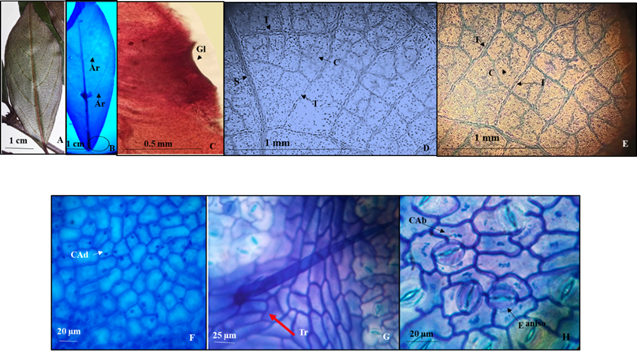
Lámina 7 Colubrina johnstonii. A.- Lámina foliar del ejemplar de herbario. B.- Lámina foliar diafanizada mostrando las tenues venas de segunda categoría formando arcos y el círculo que está en la base de la fotografía es el lugar donde se encuentra un par de pequeñas glándulas basales. C.- Detalle de una glándula basal. D.- Venas de segunda tercera y cuarta categoría. E.- Acercamiento de venas de tercera y cuarta categoría, estas últimas con venas libres o ramificadas. F.- Células adaxiales pentagonales con paredes rectas. G.- tricoma sin base radial de células epidérmicas modificadas. H.- Células abaxiales con paredes onduladas y estomas anomocíticos. Referencias: P vena de primera categoría. S vena de segunda categoría. T vena de tercera categoría. C vena de cuarta categoría. Ar arcos de las venas secundarias. Gl glándula. CAd células de la superficie adaxial. CAb células de la superficie abaxial. E estoma.
Hojas simples, simétricas, obovadas u obovado-lanceoladas de 4 a 15 cm de largo y 2 a 3.4 cm de ancho, ápice acuminado a redondeado, base cuneada a aguda, textura membranácea, margen entero con un par de glándulas cerca de la base de la lámina foliar poco visibles en los ejemplares de herbario de 0.2 a 0.3 mm (Figs. 9A, 9B). Peciolo de 4-17 mm de largo y 1 a 2 mm de ancho (Fig. 7A). Vena de primera categoría pinnada de curso recto (Figs. 7A, 7B), las de segunda categoría broquidódroma formando de 6 a 7 arcos de cada lado que no llegan hasta el margen (Figs. 7A, 7B) y en la base de la lámina foliar (ver círculo de la Figura 7B) con glándulas poco visibles a simple vista y que se aprecian con la diafanización (Fig. 7C), venas de tercera categoría alternas percurrentes (Fig. 7D, 7E), las de cuarta reticuladas con una o varias venas libres incluidas (Fig. 7E).
Epidermis en vista superficial. Células de la epidermis adaxial isodiamétricas de forma pentagonal con paredes de contorno recto (Fig. 7F), las abaxiales con células epidérmicas pentagonales con paredes de contorno ondulado (Fig. 7H). Estomas ausentes en la superficie adaxial, en la abaxial anisocíticos de 21.1 a 12.6 µm de longitud y ancho, densidad estomática de 12/mm2. Tricomas ausentes en la superficie adaxial, en la abaxial muy escasos, unicelulares eglandulares, sin base radial de células epidérmicas modificadas de 18.4 a 23.21 µm de largo (Fig. 7G), densidad promedio de 1/mm2.
8. - Colubrina triflora Brongn. (Lámina 8)
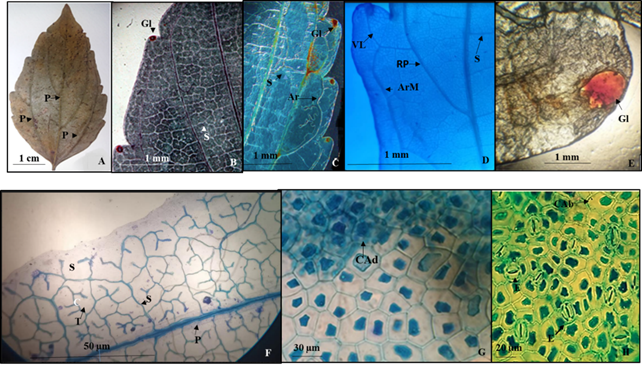
Lámina 8 Colubrina triflora. A.- Lámina foliar de ejemplare de herbario. B.- Detalle de parte de una hoja mostrando venas de segunda y tercera categoría penetrando la glándula del diente. C.- Parte de una hoja diafanizada mostrando las ramificaciones de las venas primarias formando arcos marginales y la vena que se dirige a la glándula. D.- Arco marginal que se forma de una ramificación de una vena primaria, de donde deriva una vena que penetra a la glándula. E.- Glándula. F.- Venas de primera, segunda y tercera categoría, estas últimas con venas libre ramificadas. G.- Células adaxiales pentagonales con paredes rectas. H.- Células abaxiales pentagonales con paredes rectas y estomas anomocíticos. Referencias: P venas primera categoría. S vena segunda categoría. T vena de tercera categoría. RP ramificación de una vena de primera categoría. ArM arco marginal. VL vena libre. CAd células de la superficie epidérmicas adaxiales. CAb células de la superficie epidérmica abaxial. E estomas.
Hojas simples, simétricas, ovadas a lanceolado-ovadas de 3 a 14 cm de largo y 1.0 a 6.5 cm de ancho, ápice acuminado, base redondeada, textura de la lámina foliar membranácea, margen dentado de 3 a 10 dientes de cada lado, distantes (Figs. 8A) y con una glándula de 1 a 2 mm de diámetro de diámetro (Fig. 8B, 8C, 8D, 8E). Venación acródroma basal imperfecta con tres venas primarias (Fig. 8A), el par inferior externo de las venas de primera categoría se ramifican cerca del margen formando arcos marginales de donde deriva una vena que penetra a la glándula que se encuentra en el diente (Figs. 8C, 8D), en la parte apical de la lámina foliar, en el margen no se forman arcos y las glándulas del diente son alimentadas por una vena de segunda categoría (Fig. 8B), en el resto de la lámina, las venas de segunda categoría cruzan entre las venas de primera categoría y sus ramificaciones, se anastomosan (Fig. 8B) y las de tercera con venas libres ramificadas (Fig. 8F).
Epidermis en vista superficial. Células epidérmicas adaxiales isodiamétricas con arreglo pentagonal (Fig. 8G), las abaxiales isodiamétricas con arreglo poligonal irregular (Fig. 8H), ambas superficies con paredes de contorno recto. Estomas de ambas superficies de la lámina foliar anomocíticos, los adaxiales de 18.9 a 14.7 µm de longitud y ancho, con densidad estomática de 24/mm2, los abaxiales de 19.4 µm y 18.9 µm, con densidad estomática de 34/mm2 (Fig. 8H). Hojas glabras.
9.- Colubrina viridis (M. E. Jones) M. C. Johnston (Lámina 9)
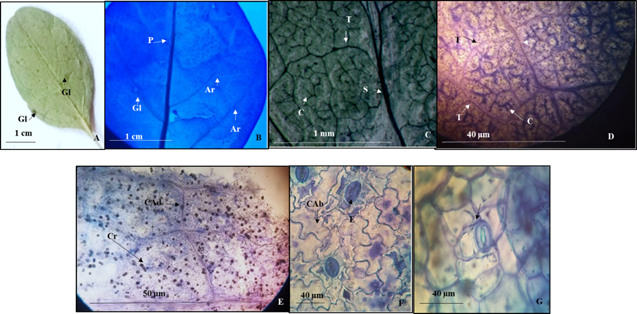
Lámina 9 Colubrina viridis. A.- Lámina foliar del ejemplar de herbario mostrando las glándulas en la superficie abaxial. B.- Lámina foliar diafanizada mostrando parte de los arcos de las venas secundarias y una glándula. C. Venas de segunda, tercera y cuarta categoría. D. Detalle de las venas de tercera y cuarta categoría, estas últimas con venas libres ramificadas. E.- Células epidérmicas adaxiales con paredes recta y con cristales prismático. F.- Células epidérmicas abaxiales con paredes onduladas y estomas anomotetracíticos. G.-. Detalle de un estoma. Referencias: P vena primera categoría. S segunda categoría. T vena de tercera categoría. C vena cuarta categoría. Ar arcos que forman las venas secundarias. Cr cristales. CAd células de la superficie epidérmica adaxial. CAb células de la superficie epidérmica abaxial. E estomas.
Hojas simples, simétricas, obovadas a veces elípticas a ovadas de 0.6 a 4.2 cm de largo y 0.3 a 2 cm de ancho, ápice redondeado o agudo a veces retuso, base redonda a cuneada (Fig. 9A), textura membranácea, margen entero, glándulas presentes de 0.1 a 0.2 mm de diámetro, redondas poco evidentes debido a que son de un color muy semejante al de la lámina foliar (Figs. 9A, 9B). Peciolo de 1 a 16 mm de largo y 1 mm de diámetro (Fig. 9A). Vena de primera categoría pinnada con curso recto (Figs. 9A, 9B). Venas de segunda tenues broquidódromas (Fig. 9B) formando 3 a 4 arcos por cada lado sin ramificaciones, venas de tercera categoría alternas percurrentes (Figs. 9C, 9D), las de cuarta reticuladas con venas varias veces ramificadas (Figs. 9C, 9D).
Epidermis en vista superficial. Células epidérmicas adaxiales con paredes de contorno recto y con cristales prismáticos (Fig. 9E) y las abaxiales con paredes de contorno ondulado (Fig. 9F). Estomas anomotetracíticos en ambas superficies (Figs. 9F, 9G), en la adaxial los estomas de 42.2 y 21.1 µm de longitud y ancho con densidad estomática promedio de 21/mm2, la abaxial de 24.3 y 28.7 µm de longitud y ancho con densidad estomática de 28 /mm2. Tricomas ausentes.
En el trabajo de Fernández-Nava et al. (2004) y en esta investigación se han documentado las peculiaridades de de arquitectura foliar y de la morfología epidérmica de las láminas foliares, de los diferentes subgéneros y secciones del género Colubrina presentes en México.
En la Tabla 2 se presenta el resumen de las características de los subgéneros según Johnston (1971) y las respectivas secciones, así como la distribución geográfica de las mismas. Dicha clasificación fue la base de este trabajo para la discusión y permanencia de algunos subgéneros y secciones, así como la posible reedescripción de estas categorías taxonómicas.
Tabla 2 Características y distribución de los Subgéneros y Secciones de Colubrina presentes en México (Johnston, 1971).Table 2. Characteristics and Distribution of Colubrina Subgenera and Sections present in Mexico (Johnston, 1971).
| SUBGÉNERO COLUBRINA | ||
|---|---|---|
| Láminas foliares con margen entero, crenado o aserrado, los dientes pueden o no presentar glándulas marginales, de encontrarse estas últimas, entonces el número de dientes es menor de diez por lado. | ||
| I. Sección Colubrina Láminas foliares con margen entero u onduladas-crenadas, con glándulas longitudinales entre la nervadura central y l os márgenes, a menudo las dos más cercanas a la base de la lámina foliar son más grandes que el resto. Se distribuye en Hawái con una especie y cuatro taxones en los trópicos americanos. |
II. Sección Cowania Láminas foliares con al menos dos glándulas marginales, una a cada lado, a menudo cerca de la base de la lámina foliar, margen entero. Ramas de la inflorescencia comúnmente con espinas. Distribución América tropical |
III. Sección Barcena Lámina con el margen de la hoja con 3 a 10 dientes de cada lado, cada uno asociado a una glándula marginal. Distribución México y América Central |
| Especie presente en México: C. arborescens |
Especies presentes en México: C. elliptica C. heteroneura C. johnstonii C. viridis C. spinosa var. mexicana* (Nayarit) |
Especies presentes en México: C. ehrenbergii C. triflora |
| SUBGÉNERO SERRATARIA | ||
| >Láminas foliares con márgenes aserrados, más de diez dientes en cada lado y cada uno asociado con una glándula marginal. Distribución América y Asia. | ||
| Se incluyeron las siguientes especies: C. californica, C. celtidifolia, C. texensis
1 (sin. C. texensis var. texensis), C. stricta 1 (sin.C. texensis var. pedunculata), C. angustior 1 (sin. C. greggii var.angustior), C.greggii 1 (sin.C.greggii var greggii), C. yucatanensis 1 (sin. C. greggii var. yucatensis), C. macrocarpa 2 (sin. C. macrocarpa var. macrocarpa) y C. lanulosa 2 (sin. C. macrocarpa var. lanulosa). C. sordida* (Guerrero) C. macrocarpa var. macrocarpoides* (Querétaro). | ||
Las especies que incluyeron* correspondieron aquellas que se citan para México y solo se cuenta con el ejemplar tipo y no se han vuelto a recolectar, estos taxones no fueron considerados para este estudio. Algunos nombres de las especies se han tenido que actualizar o darles la categoría taxonómica de especies. Los superíndices 1 corresponde al trabajo de Nesom (2013) y el 2 al de Fernández-Nava et al. (2024).
Una vez que se obtuvieron los datos de la arquitectura foliar y superficies epidérmicas de las nueve especies estudiadas y con el propósito de establecer las características de los 16 taxones presentes en México, se elaboró la Tabla 3 que se encuentra después de la bibliografía, donde se sintetizaron las descripciones de las nueve especies aquí estudiadas más las siete que están incluidas en Fernández-Nava et al. (2024) y con base en estos resultados se realizó la clave dicotómica para la separación de los taxones así como el análisis UPGMA, índice de similitud de Jaccard y el respectivo dendrograma. Las actividades anteriores permitieron separar y agrupar las 16 especies a partir estudios morfológicos y anatómicos y relacionarlos con los trabajos taxonómicos de Colubrina .
Tabla 3 Comparación de la arquitectura foliar y las superficies epidérmicas de las especies estudiadas del género ColubrinaTable 3. Comparison of leaf architecture and epidermal surfaces of the studied species of the genus Colubrina
| Especie | Primera categoría |
Segunda categoría |
Par de venas secundarias inferiores ramificadas/ no |
Tercera categoría |
Cuarta categoría |
Alimentación diente |
Con o sin glándulas posición |
Células adaxiales |
Células Abaxiales |
Tipos de estomas |
Base de los tricomas |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1.- C. angustior | Pinnada | Craspedó-dromas, con 6-7 pares de venas secundarias de cada lado | primer par basal ramificado | Reticuladas opuestas percurrentes, en el margen alimentan a los dientes | Reticula-das | Venas tercera categoría alimentan los dientes del margen, pueden o no formar arcos | Sin glándulas | Hexago-nales, paredes rectas | Paredes onduladas | Anomo-cíticos Adaxial ausen-tes | Sin base radial de células epidér-micas modifi-cadas |
| 2.- C. arbores-cens | Pinnada | Craspedo-droma 3-4 pares de venas secunda-hhrias de cada lado | Par inferior no ramificado | Reticuladas opuestas percurrentes | Reticula-das | Margen entero | Distri-buidas en la super-ficie de la hoja | Pentagona-les con paredes rectas | Rectan-gulares con paredes rectas | Ano-mo-cíticos ambas super-ficies | Base radial de células epidér-micas modifi-cadas |
| 3.- C. californica | Pinnada | Broquidó-dromas 3-4 arcos | Par inferior ramifica-do, arcos superiores enrollados | Reticuladas alternas per-currentes | Reticula-das | Margen entero | Sin glándulas | Pentagona-les con paredes rectas | Pentagonales con paredes rectas | Ano-mocíti-cos en ambas super-ficies | Base radial de células epidér-micas modifi-cadas |
| Especie | Primera categoría | Segunda categoría | Par de venas secundarias inferiores ramificadas/ o no | Tercera categoría | Cuarta categoría | Alimenta-ción diente | Con o sin glándu-las posición | Células adaxiales | Células Abaxiales | Tipos de esto-mas | Base de los tricomas |
| 4.-C. celtidifolia | Acródroma basal imperfecta con tres venas, las dos externas se ramifican hacia el margen formando arcos y de ellos sale una vena que entra al diente | Cruzan entre las venas primarias anastomosándose | Poligonales reticuladas con o sin venas libres ramifica-das | Las ramifi-caciones de las venas de primera categoría forman arcos y de ahí deriva una vena que alimenta al diente | Sin glándulas | Células hexagona-les con paredes rectas | Células hexago-nales con paredes rectas | Ano-mocíti-cos, abaxiales | Base radiada de células epidér-micas modifi-cadas | ||
| 5.- C. ehrenbergii | Acródroma basal imperfecta con tres venas con ramificacio-nes libres | Recorren trayectos paralelos entre las venas pri-marias y sus ramifi-caciones | Libres ramificadas | Glándu-las en cada diente donde entra una vena de segunda categoria | Pentagonales con paredes rectas | Rectangu-lares con paredes rectas | Ano-mocíti-cos ambas super-ficies | Sin base radial de células epidér-micas modifi-cadas | |||
| 6.-C. elliptica | Pinnada | Broquidó-droma con 3-4 arcos | No rami-ficados | Reticuladas alternas percurrentes | Reticula-das con o sin venas libres | Margen entero | Con dos glándulas basales | Pentagonal-hexagona-les con paredes rectas | Rectangu-lares con paredes rectas | Anficicloci-ticos abaxial | Sin base radial de células epidér-micas modifi-cadas |
| 7.-C. greggii | Acródroma basal imperfecta con las tres venas primarias ramificadas | reticuladas | Reticuladas alteñas percurrentes | Ramifica-ciones de venas prima-rias forman arcos y una vena va al diente | Sin glándulas | Hexago-nales paredes rectas | Hexago-nales paredes rectas | Ano-mo-cíticos | Sin base radial de células epidér-micas modifi-cadas | ||
| 8.-C. heteroneura | Pinnada | Broquidó-droma con 5-6 arcos | No ramificada | Reticulada alternas percurrentes | Reticula-das sin venas libres incluidas | Margen entero | Marginales basales | Arreglo irregular con paredes onduladas | Pentagonales con paredes rectas | Adaxial anisocitico y paraci-tico, abaxial aniso-citicos y ano-mote-tracíti-cos | Sin tricomas |
| 9.-C. johnstonii | Pinnada | Broquidó-droma con 6-7 arcos | No se ramifica | Reticuladas alternas percurrentes | Reticula-das con una o varias venas ramifica-das | Margen entero | Un par basales margina-les | Pentagonal paredes rectas | Irregulares paredes ondula-das | Aniso-citico aba-xial | Sin base radial de células epidér-micas modifi-cadas |
| 10.-C. lanulosa | Pinnada | Craspedó-dromas, par inferior lateral ramifi-cadas con 5-8 pares de venas secunda-rias de cada lado | Con ramifica-ciones | Reticuladas, alternas percurrentes | Reticula- das sin venas incluidas o de tenerlas solo una | Venas de segunda y tercera categoría alimentan los dientes del margen | Sin glándulas | Forma irregular paredes rectas | Forma irregular paredes rectas | Ano-mo- citicos y ano-mote-traciti-cos abaxial | Base radial con células epidér-micas modifi-cadas |
| 11.-C. macrocarpa | Acródroma basal imperfecta con las tres venas primarias ramificadas | Reticuladas | Reticuladas, incluyen venas libres | Venas de segunda y tercera categoría alimentan los dientes del margen | Sin glándulas | Rectangulares paredes rectas | Forma irregular paredes rectas | Aniso-cíticos y peri-cítico abaxial | Base radial con células epidér-micas modifi-cadas | ||
| 12.- C. stricta | Pinnada | Craspedó-dromas con 5-6 pares de venas secunda-rias de cada lado | primer par basal no ramificado | Reticuladas, alternas percurrentes | Reticula- das con 1 vena no ramifica-da | Venas de tercera categoría alimentan los dientes del margen | Sin glándulas | Hexago-nales paredes rectas | Hexago-nales paredes rectas | Ano-mo-tetra-citicos Abaxial | Base radial con cé-lulas epidérmicas modi-ficadas |
| 13.- C. texensis | Pinnada | Craspedó-dromas con 3-4 venas | Primer par basal ramificado | Reticuladas alternas percurrentes | Reticula-das con venas 2 o más ramifica-das | Venas de segunda categoría y muy pocas de tercera alimentan los dientes del margen | Sin glán-dulas | Con paredes onduladas | Con paredes ondula-das | Ano-mo-cítico Aba-xial | Base radial con células epidérmicas modifi-cadas |
| 14.- C. triflora | Acródroma basal imperfecta con tres venas | Recorren entre las venas de primera categoría y sus ra-mificacio-nes, reticu-ladas | Ramifica-das | Reticuladas con venas libres | Ramificacio-nes de venas primarias forman arcos y una vena de segunda categoría entra al diente | Presen-tes en cada diente | Pentagonales con paredes rectas | Pentagonales paredes rectas | Ano-mociti-cos en ambas super-ficies | Sin trico-mas | |
| 15.- C. viridis | Pinnada | Broquidó-droma con 3 a 4 arcos | No ramifica-da | Reticuladas alternas percurrentes | Reticuladas con venas ramifica-das | Margen entero | Glán-dulas espar-cidas en la lámina foliar | Rectangu-lares paredes rectas | Forma irregular paredes onduladas | Ano-mote-traci-tico en ambas super-ficies | Sin trico-mas |
| 16.- C. yucatensis | Pinnada | Craspedó-dromas, con 6 a 7 pares de venas secundarias de cada lado | primer par basal ramificado | Reticula- das opuestas percurrentes | Reticula- das sin venas libres incluidas | Venas tercera categoría alimentan los dientes del margen a veces forman arcos | Sin glándu-las | Poligona-les, paredes rectas | Forma irregular, paredes onduladas | Ano-mo-cítico Aba-xial | Base radial con células epidér-micas modifi-cadas |
Clave dicotómica para la identificación de especies de colubrina con base en la arquitectura foliar y superficies epidermicas incorporando datos deFernández-Nava et al. (2024).
Dichotomous key for the identification of colubrina species based on leaf architecture and epidermal surfaces incorporating data fromFernández-Nava et al. (2024).
1.- Venas de primera categoría pinnada (vena central evidente)
2.- Venas de segunda categoría craspedódromas (venas secundarias terminan en el margen)
3.- Glándulas distribuidas en ambas superficies foliares, lámina foliar con 3 a 4 pares de
venas a cada lado de la lámina foliar……………………………………C. arborescens
3.- Glándulas ausentes en la superficie, margen o base de las láminas foliares
4.- Par inferior de las venas de segunda categoría no ramificadas. Lámina foliar con 5 a 6
pares de venas de cada lado……………………………………………….…..C. stricta
4.- Par inferior de las venas de segunda categoría ramificadas
5.- Tricomas sin la base de células epidérmicas modificadas con 6 a 7 pares de
venas en ambos lados de la lámina foliar…………………………………...C. angustior
5.- Tricomas con base radial de células epidérmicas modificadas
6.- Venas de cuarta categoría sin venas libres incluidas, o de tenerlas solo una. Lámina foliar
con 6 a 8 pares de nervaduras de segunda categoría de cada lado
7.- Células epidérmicas adaxiales y abaxiales con paredes de contorno recto, estomas
anomocíticos y anomotetracíticos. Lámina foliar con 7 a 8 pares de venas secundarias de
cada lado………………………………………………………………….…..C. lanulosa
7.- Células epidérmicas adaxiales y abaxiales con paredes de contorno recto u ondulado
respectivamente, estomas anomocíticos. Lámina foliar con 6 a 7 pares de venas
secundarias de cada lado………………………………………………...…C. yucatensis
6.- Venas de cuarta categoría con varias venas libres ramificadas. Lámina foliar con de 3 a 4
pares de nervaduras secundarias de cada lado…………………………….C. texensis
2.- Venas de segunda categoría broquidódromas (venas secundarias unidas en una serie de
arcos)
8.- Láminas foliares sin glándulas, par inferior de arcos que forman las venas de segunda
categoría ramificadas y las nervaduras apicales enrolladas………..……C. californica
8.- Láminas foliares con glándulas basales o en toda la superficie
9.- Láminas foliares con glándulas en toda la superficie, tricomas ausentes
……………………….C. viridis
9.- Láminas foliares con glándulas basales marginales, tricomas presentes o ausentes
10.- Tricomas ausentes, venas de segunda categoría forman 5 a 6 arcos de cada
lado………………………………………………………………..…..C. heteroneura
10.- Tricomas presentes
11.- Venas de segunda categoría forman de 3 a 4 arcos a cada lado de la lámina foliar
………..C. elliptica
11.- Venas de segunda categoría forman de 6 a 7 arcos a cada lado de la lámina foliar,
……….C. johnstonii
1.- Venas de primera categoría acródromas basal imperfecta (tres o más venas primarias
divergiendo desde un solo punto)
12.- Glándulas presentes en cada diente del margen de la lámina foliar, tricomas ausentes o
presentes
13.- Tricomas sin base radial de células epidérmicas modificadas, ramificaciones de las tres
venas primarias libres, margen dentado y en cada diente se encuentra una glándula
……………………….…………………………………………...….C. ehrenbergii
13.- Tricomas ausentes, las ramificaciones del par basal externo de las venas primarias
forman arcos de donde deriva una vena que penetra en cada diente donde se encuentra
una glándula …………………………………………………….………..C. triflora
12.- Glándulas ausentes, lámina foliar con tricomas
14.- Tricomas sin células radiales epidérmicas modificadas, nervaduras de tercera categoría
sin venas libres……………………………………………………………C. greggii
14.- Tricomas con base radial de células epidérmicas modificadas
15.- Venas de primera categoría externas con ramificaciones libres y los dientes
del margen son alimentados por venas de segunda o tercera categoría…..C. macrocarpa
15.- Venas de primera categoría externas se ramifican y forman arcos y de ellas deriva una
nervadura que alimenta los dientes marginales………………………….C. celtidifolia
Al analizar los caracteres de las 16 especies de Colubrina, con base en los tipos de arquitectura foliar, craspedódroma, broquidódromas, acródroma basal imperfecta; presencia o ausencia de glándulas; con o sin tricomas y en caso de presentarse con o sin base radial de células epidérmicas modificadas; ramificaciones del primer par de venas primarias y secundarias ramificadas o no; venas de tercera categoría con venas libres incluidas o no; venas de cuarta categoría con venas libres o no; tipos de tricomas anisocíticos, anomocíticos, paracíticos, anficíclicos, anomotetracíticos, pericíticos. Para validar los caracteres de relevancia utilizados en la identificación de especies se realizó el análisis UPGMA y se obtuvo del índice de similitud de Jaccard, con base en la relación presencia-ausencia entre los taxones (Tabla 4) y el respectivo dendrograma (Fig. 10).
Tabla 4 Índice de similitud de Jaccard entre las 16 especies de Colubrina.Table 4. Jaccard similarity index between the 16 species of Colubrina.
| agustior | arboresc | californ | celtidif | ehrenber | elliptica | greggii | heteron | johnston | lanulosa | macrocar | stricta | texense | triflora | viridis | yucate | |
| agustior | 0 | 2.645751 | 2.236068 | 2.6458 | 2.236068 | 3.4641 | 2.236 | 3 | 3 | 2 | 3 | 2.6457513 | 1.73205 | 2.902 | 3.317 | 1.732 |
| arboresc | 2.64575 | 0 | 2.44949 | 2.4495 | 2.44949 | 2.6458 | 2.449 | 2.4495 | 2.82843 | 2.2361 | 2.828727 | 2.4494897 | 2.44949 | 2.294 | 2.449 | 2 |
| californ | 2.23607 | 2.44949 | 0 | 2.4495 | 2.828427 | 3 | 2.449 | 2.8284 | 3.16228 | 1.7321 | 2.828427 | 2.8284271 | 2 | 3.078 | 2.828 | 1.414 |
| celtidif | 2.64575 | 2.44949 | 2.44949 | 0 | 2.44949 | 3 | 2 | 2.8284 | 3.16228 | 2.6458 | 2 | 2.4494897 | 2.44949 | 2.294 | 3.162 | 2.449 |
| ehrenber | 2.23607 | 2.44949 | 2.828427 | 2.4495 | 0 | 3 | 2 | 2.4495 | 2.44949 | 3 | 2.828427 | 3.1622777 | 2.82843 | 1.777 | 2.828 | 2.828 |
| elliptica | 3.4641 | 2.645751 | 3 | 3 | 3 | 0 | 3.317 | 2.6458 | 2.23607 | 3.4641 | 3 | 2.6457513 | 3 | 2.902 | 2.646 | 3.317 |
| greggii | 2.23607 | 2.44949 | 2.44949 | 2 | 2 | 3.3166 | 0 | 2.8284 | 2.82843 | 2.6458 | 2.44949 | 2.8284271 | 2.82843 | 2.294 | 2.828 | 2.449 |
| heteron | 3 | 2.44949 | 2.828427 | 2.8284 | 2.44949 | 2.6458 | 2.828 | 0 | 2.44949 | 3.3166 | 3.162278 | 3.1622777 | 3.16228 | 1.777 | 2 | 3.162 |
| johnston | 3 | 2.828427 | 3.162278 | 3.1623 | 2.44949 | 2.2361 | 2.828 | 2.4495 | 0 | 3.6056 | 2.828427 | 2.8284271 | 3.16228 | 2.513 | 2.449 | 3.464 |
| lanulosa | 2 | 2.236068 | 1.732051 | 2.6458 | 3 | 3.4641 | 2.646 | 3.3166 | 3.60555 | 0 | 3 | 2.236068 | 1.73205 | 3.244 | 3 | 1 |
| macrocar | 3 | 2.828427 | 2.828427 | 2 | 2.828427 | 3 | 2.849 | 3.1623 | 2.82843 | 3 | 0 | 2.4494897 | 2.82843 | 2.513 | 3.162 | 2.828 |
| stricta | 2.64575 | 2.44949 | 2.828427 | 2.4495 | 3.162278 | 2.6458 | 2.828 | 3.1623 | 2.82843 | 2.2361 | 2.44949 | 0 | 2 | 3.078 | 2.828 | 2.449 |
| texense | 1.73205 | 2.44949 | 2 | 2.4495 | 2.828427 | 3 | 2.828 | 3.1623 | 3.16228 | 1.7321 | 2.828427 | 2 | 0 | 3.078 | 3.464 | 1.414 |
| triflora | 2.90191 | 2.294157 | 3.077935 | 2.2942 | 1.777047 | 2.9019 | 2.294 | 1.777 | 2.51312 | 3.2444 | 2.513123 | 3.0779351 | 3.07794 | 0 | 2.294 | 3.078 |
| viridis | 3.31662 | 2.44949 | 2.828427 | 3.1623 | 2.818427 | 2.6458 | 2.828 | 2 | 2.44949 | 3 | 3.162278 | 2.8284271 | 3.4641 | 2.294 | 0 | 3.162 |
| yucaten | 1.73205 | 2 | 1.414214 | 2.4495 | 2.828427 | 3.3166 | 2.294 | 3.1623 | 3.4641 | 1 | 2.828427 | 2.4494897 | 1.41421 | 3.078 | 3.162 | 0 |
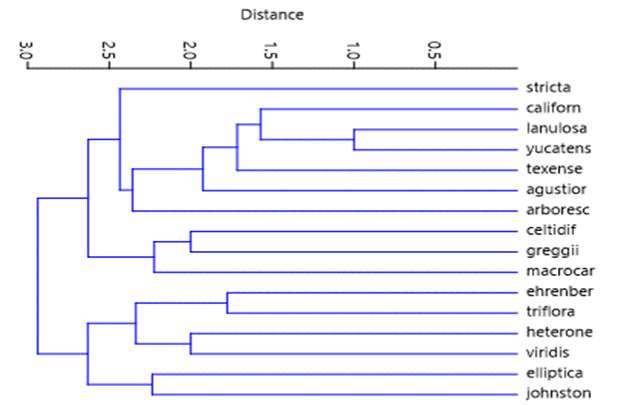
Figura 10 Dendrograma mostrando las relaciones de afinidad entre las diferentes especies de Colubrina.
Al observar el dendrograma se aprecian dos grandes grupos, el primero incluye a Colubrina ehrenbergii, C. triflora, C. heteroneura, C. elliptica, C. johnstonii y C. viridis que se separan por la presencia de glándulas generalmente en el margen y en la última especie en la superficie de la lámina. En el segundo conjunto se diferencian aquellas especies que no presentan glándulas como C. celtidifolia, C. greggii, C. macrocarpa, C. angustior, C. texensis, C. yucatanensis, C. lanulosa, C. californica, C. stricta y C. arborescens que las presentan en toda la superficie de la lámina, son oscuras y evidentes. De este último conjunto de diez especies se separan a su vez en dos agrupamiento, uno incluye a los tres primeros taxones con arquitectura foliar acródroma basal imperfecta y el segundo al resto de las especies con arquitectura foliar craspedódroma y broquidódroma.
Discusión
Después de describir la arquitectura foliar y las superficies epidérmicas de las especies de Colubrina que se distribuyen en México se encontraron caracteres para separar cada uno de los taxones, como la arquitectura foliar; número de pares de venas secundarias en la superficie foliar; ramificación del par basal de venas secundarias o no, y si la ramificación es libre o formaban arcos; la presencia de glándulas y su ubicación; el tipo de tricomas que se presentan con base radial de células epidérmicas modificadas o si carecen de ellas, o bien la ausencia de tricomas. El contorno de las paredes celulares de las células epidérmicas (rectas u onduladas) y los tipos de estomas no fueron relevantes para la separación de los taxones.
Tomando en consideración el trabajo de Johnston (1971, pág. 8) , quien dividió a las especies de Colubrina en varios subgéneros y secciones, las especies de México se incluyen en los subgéneros Colubrina con las secciones Colubrina, Cowania y Barcena y el subgénero Serrataria no presenta secciones (Tabla 2).
El subgénero Colubrina (Johnston 1971, pág. 9) incluye a especies con los siguientes caracteres: láminas foliares con margen entero, crenado o aserrado, los dientes no asociados con glándulas marginales o de encontrarse en el margen, entonces con menos de diez dientes por cada lado. Se incluye a siete especies, que cumplen estas condiciones según las observaciones del trabajo arriba citado: C. arborescens, C. ehrenbergii, C. elliptica, C. heteroneura, C. johnstonii, C. triflora y C. viridis, (Tabla 2).
La subdivisión del subgénero Colubrina en secciones propuesta por (Johnston 1971, pág. 9) incluyen los caracteres que se mencionan:
Subgénero Colubrina sección Colubrina con láminas foliares con margen entero u onduladas-crenadas, con glándulas longitudinales entre la nervadura central y los márgenes, a menudo las dos glándulas más cercanas a la base son más grandes que el resto. En México se encuentra A. arborescens, en este trabajo se corrobora la presencia de glándulas en la superficie foliar, más visibles en la superficie abaxial de la lámina foliar, pero no son longitudinales (Figs. 1E, 1F).
Subgénero Colubrina sección Cowania con láminas foliares con al menos dos glándulas marginales una a cada lado, a menudo cerca de la base de la hoja. Los ejemplares de C. elliptica presentaron un par de glándulas más o menos visibles en los ejemplares de herbario y en hojas diafanizadas que se observaron en el microscopio estereoscópico (Fig.5A, 5B). En C. heteroneura (Fig. 6F) y C. johnstonii (Fig. 7C) también con un par de glándulas basales, poco visibles en ejemplares de herbario y es necesario diafanizar las hojas y realizar cortes para observarlas en microscopio óptico. En C. viridis las glándulas se aprecian en la superficie de la lámina foliar y son de color claro (Figs. 9A, 9B).
Subgénero Colubrina sección Barcena presenta el margen de la hoja con 3 a 10 dientes de cada lado, asociados a una glándula marginal que se observan muy bien. En este trabajo se constató la presencia de glándulas en los dientes del material diafanizado de C. ehrenbergii (Figs. 4D, 4E) y C. triflora (Figs. 8B, 8C, 8E).
El subgénero Serrataria según Johnston (1971, pág. 10) las láminas foliares presentan márgenes aserrados, más de diez dientes en cada lado y cada uno asociado con una glándula marginal donde se incluyen a C. californica, C. celtidifolia, C. texensis, C. stricta C. angustior, C.greggii, C. yucatanensis, C. macrocarpa y C. lanulosa, sin embargo, en los resultados de este trabajo y en el de (Fernández-Nava et al., 2024) ninguna de estas especies presenta glándulas en los dientes. Lo que se apreció en las especies de ejemplares herborizados fue una pequeña protuberancia en los dientes, sin embargo no son glándulas y este abultamiento se debe a que los dientes están alimentados por una vena de segunda o tercera categoría como sucede en C. angustior (Fernández-Nava et al. 2024 p. 16 Fig. 3D), C. californica (Figs. 2B, 2D), C. celtidifolia (Fig. 3D), C.greggii (Férnandez-Nava et al. 2024 p. 14 3D), C. lanulosa (Fernández-Nava et al. 2024 p. 19, Figs. 6C, 6D), C. macrocarpa (Fernández-Nava et al. 2024 p. 18, 5D), C. stricta (Fernández-Nava et al., 2024 p. 10, Fig. 7C), C. texensis (Férnandez-Nava et al. 2024 p. 21, Fig. 8D y C. yucatanensis (Férnandez-Nava et al. 2024 p. 17, Fig. 4D). Lo anterior lleva a reconsiderar las características del subgénero Serrataria según (1971, págs. 9-10).
En el trabajo de Johnston (1971, pág. 10) se describen las características de las especies del subgénero Colubrina sección Capuronia, con hojas enteras o con pequeños dientes, en uno o ambos lados o irregularmente crenulado dentado, no asociados con glándulas marginales. Estas características son las que se encontraron en los taxones estudiados para México del subgénero Serrataria, sin embargo, el autor citado da un peso importante a la distribución de las especies y la sección Capuronia que incluye a cuatro taxones son de Madagascar y además hay características en el fruto que no se presentan en las especies de Serrataria, así que más bien las especies de este subgénero tendrán que redescribirse considerando la ausencia de glándulas y la penetración en los dientes marginales a través de venas de tercera o cuarta categoría.
Con base en los datos de arquitectura foliar y superficies epidérmicas que se observaron en este trabajo, fue posible encontrar patrones en común con las especies citadas por (Johnston 1971, pág. 9), en el subgénero Colubrina sección Cowania con C. elliptica (Fig 5A,5B) C. heteroneura (Fig. 6A), C. johnstonii (Figs. 7A, 7B) y C. viridis (9A) que presentan en común además de la presencia de glándulas, las venas de segunda categoría broquidódromas y los arcos inferiores no se ramifican.
Así también con los datos de arquitectura foliar y superficies epidérmicas, fue posible encontrar aspectos en común con las especies citadas por (Johnston 1971, pág. 9) para el subgénero Colubrina sección Barcena con C. ehrenbergii y C. triflora, con la presencia de las glándulas en cada diente del margen se encontró que el patrón de venación fue acródroma basal imperfecta, con tres o más venas primarias divergiendo desde un solo punto.
El subgénero Serrataria que está representado en México con nueve especies (Johnston, 1971, pág. 10) se encontraron tres tipos de arquitectura foliar, la craspedódroma presente en C. angustior, C. lanulosa, C. stricta, C. yucatensis y C. texensis. La acródroma basal imperfecta en C. greggii, C. macrocarpa y C. celtidifolia, el patrón broquidódromo en C. californica.
En relación a los tricomas para el subgénero Serrataria se presentaron tres variables en los taxones: con base radial de células epidérmicas modificadas, los que no tienen estas células epidérmicas modificadas y las especies que no presentan tricomas.
Las especies con venación craspedódroma como C. angustior, los tricomas no tienen base radial de células epidérmicas modificadas y C. lanulosa, C. stricta, C. texensis y C. yucatensis, si los presentan.
En los taxones con arquitectura foliar acródroma basal imperfecta, en C. greggii los tricomas no presentan la base radial de células epidérmicas modificadas; C. celtidifolia y C. macrocarpa los pelos tienen la base radial de células epidérmicas modificadas. En el patrón de venación broquidódromo, C. californica presenta tricomas con la base radial de células epidérmicas modificadas. Por lo anterior, entre las especies del subgénero Serrataria variaron los patrones de la arquitectura foliar y de los tricomas así que el carácter común del subgénero fue la ausencia de glándulas en los dientes marginales.
El trabajo de Wendt (1983, p. 87) incluye una clave para separar las cinco especies de la sección Cowania a nivel mundial y la base de la separación incluye las glándulas y como se mencionó en el presente trabajo, en especímenes de herbario son poco visibles, sin embargo, las especies que prosperan en México tienen un carácter en común que es presentar el tipo de venación broquidódroma y el par basal de estos arcos no ramificados y como una segunda opción de separación las glándulas y así por ejemplo, C. elliptica presenta de 3 a 4 pares de arcos en la lámina foliar, tricomas con base radiada de células epidérmicas modificadas; C. johnstonii con 6 a 7 pares de arcos en la lámina foliar y tricomas sin base radial de células epidérmica modificadas; C. heteroneura con 5 a 6 arcos en la lámina foliar, sin tricomas y las nervaduras de cuarta categoría sin venas libres incluidas o con solo una y C. viridis con 3 a 4 arcos, sin tricomas y venas de cuarta categoría libres ramificadas; estos caracteres podrían apoyar la separación de las especies de la sección Cowania.
Al realizar el análisis UPGMA, el índice de similitud de Jaccard y el respectivo dendrograma (Fig. 10) se aprecian dos grandes grupos, uno que incluye a Colubrina ehrenbergii, C. triflora, C. heteroneura, C. viridis, C. elliptica y C. johnstonii. Las dos primeras especies pertenecientes al subgénero Colubrina sección Barcena y los cuatro últimos taxones son del subgénero Colubrina sección Cowaiana (Johnston, 1971 p. 9-10).
Esta separación se complementa con el presente estudio debido a que Colubrina ehrenbergii, C. triflora pertenecen a la sección Barcena con arquitectura foliar acródroma basal imperfecta y presentan glándulas en los dientes del margen. La sección Cowania con C. elliptica, C. heteroneura, C. johnstonii y C. viridis con arquitectura foliar broquidódroma, con el par de arcos inferiores no ramificados y la presencia de dos glándulas generalmente en el margen y base la hoja o bien distribuidas en la superficie de la lámina foliar.
El segundo grupo del dendrograma incluye al Subgénero Colubrina sección Colubrina y todas las especies del subgénero Serrataria. En el primer subgénero y sección se encuentra C. arborescens que se separa por presentar arquitectura foliar craspedódroma y glándulas distribuidas en toda la superficie, aunque no como las describe (Johnston, 1971, p. 12), este autor cita la presencia de glándulas longitudinales entre la nervadura central y los márgenes de la lámina foliar, también refiere que hay dos razas en el área de distribución de la especie, la que se localizan en las Antillas y otra de la Península de Yucatán, Chiapas y Guatemala (Johnston, 1971, mapa p. 13). Los ejemplares examinados en el presente trabajo pertenecen a los estados de Campeche, Quintana Roo y Chiapas y podrían tratarse de otra especie debido a que las glándulas no son longitudinales son circulares a ovadas, oscuras y se distribuyen en toda la lámina foliar (Figs. 1C, 1E).
En el subgénero Serrataria excluyendo a C. arborescens por las razones arriba citadas, se separan dos conjuntos, uno de ellos que incluye a Colubrina celtidifolia, C. greggiiy C. macrocarpa con taxones con arquitectura foliar acródroma basal imperfecta y un segundo grupo con C. angustior, C. lanulosa, C. stricta, C. texensis y C. yucatensisc con venación craspedódroma y a C. californica con venación broquidódroma con los arcos inferiores ramificados.
Este último taxón presentó tres caracteres diferentes al resto de las especies de esta sección, la primera la arquitectura foliar broquidódroma presente más bien en la sección Cowania, sin embargo, difiere de ella por la ausencia de glándulas y los arcos inferiores están ramificados y la tercera es que las venas secundarias superiores se enrollan sobre su propio eje (Figs. 2B, 2D).
Con base en las características anteriores el subgénero Serrataria tendría en común la ausencia de glándulas en los dientes marginales y la subdivisión en secciones podría darse con base en la arquitectura foliar.
Nesom (2023a, p. 1, 5) menciona que estudios de Rhamnaceae han reconocido que Colubrina celtidifolia es nativa del sur de México y Guatemala y es similar al árbol asiático Hovenia dulcis. Las similitudes se confirman por la presencia de los pedúnculos carnosos e hinchados de la inflorescencia, razón por la cual este autor transfirió C. celtidifolia al género Hovenia como H. celtidifolia (Cham. & Schlecht.) Nesom. Así también, Pool (2014, p. 60-61) indica que hay “una sola especie de Colubrina con las ramas de la inflorescencia hinchadas después de la antesis” carácter genérico de Hovenia y “un engrosamiento similar se ha visto algunas veces en C. macrocarpa (Cav.) G. Don del centro de México pero este engrosamiento es en los ovarios, los cuales son aparentemente infectados por insectos y desarrollan agallas”. La autora menciona que las láminas foliares tienen de 3 a 5 nervaduras laterales y la ausencia de glándulas y reconoce la presencia en Mesoamérica de Colubrina celtidifolia.
En la descripción de Nesom (2023b, p.5) cita para Hovenia celtidifolia el tipo de venación broquidódroma con tres venas primarias laterales y con una glándula en cada diente. Al leer la descripción del género Hovenia en la Flora de China (Chen y Schirarend, 2007) se menciona la venación trinervadas desde la base, vena primaria con 4 a 8 pares de venas secundarias y no se cita la presencia de glándulas en la lámina foliar.
En lo que respecta a la arquitectura foliar de C. celtidifolia en este trabajo se encontraron tres venas primarias que parten de la base (acródroma basal imperfecta según Dilcher, 1974, p. 33 o también conocida como actinódroma Ash et al., 1999, p. 29) y las ramificaciones de las venas de primera categoría son las que forman los arcos, no es una venación broquidódroma ya que esto implica que se presente una vena de primera categoría pinnada y los arcos se formen de las venas secundarias (Dilcher, 1974, p. 30). La presencia de glándulas en C. celtifolia solo se mencionan por Nesom (2023b, p. 5.), no se apreciaron por Pool (2014, p. 60) ni se observaron en este trabajo.
Debido a que la arquitectura foliar y la ausencia de glándulas coincide con varias especies conocidas de Colubrina de la sección Serrataria y además, la presencia de tricomas de base radial con células epidérmicas modificadas, hasta ahora descritos dentro de las Rhamnaceae para el género Colubrina y al no contar con datos de arquitectura foliar y superficies epidérmicas de Houvenia se decidió seguir utilizando el nombre de C. celtidifolia.
En el trabajo de Nesom (2023b) se describen cinco especies nuevas de Colubrina de las cuales sería interesante en un futuro poder contar con algunas hojas para poder corroborar las diferencias entre especies cercanas que el mismo autor cita, por ejemplo: Colubrina lanceolata Nesom (sp. nov.) es parecida a C. triflora difiere en el grosor de la hoja, glauca, se encuentra en BT deciduo y C. triflora en matorral, la nueva especie prospera solo en Aguascalientes. Colubrina mezquitalensis Nesom (sp. nov.) similar a Colubrina macrocarpa en sus hojas bicoloras, densamente lanosas abaxialmente, márgenes foliares con pequeños dientes. Colubrina neoviridis Nesom (sp. nov.) similar a Colubrina viridis en su hábito arbustivo, ramificado con brotes cortos, hojas pequeñas y apretadas con márgenes enteros y pecíolos filiformes. Colubrina subsessilis Nesom (sp. nov.) similar a Colubrina triflora por sus márgenes fuertemente trinervados y poco dentados-aserrados, hojas escasamente pubescentes con ápice agudo-acuminado, hojas consistentemente más pequeñas, ovadas con base subcordada y pecíolos tomentosos de 2 a 5 mm de largo. Colubrina tequila Nesom (sp. nov.) similar a Colubrina ehrenbergii por las hojas gruesas y ovaladas con relativamente pocos dientes con puntas glandulares, así también es similar a C. triflora en la forma de las hojas y textura, tricomas persistente, conocida solo de la localidad tipo y C. villarrealii Nesom que parece estar ligada con Colubrina texensis var. pedunculata M.C. Johnston.
Así también, en el trabajo de Nesom (2023b, p. 2) se cita a Colubrina asiatica (L.) Brongn. que no fue considerada en este trabajo por ser es una planta introducida.
Conclusiones
Se logró separar las dieciséis especies de Colubrina presentes en México. Es necesario revisar el subgénero Serrataria, debido a que la característica principal es la presencia de glándulas en los dientes del margen de las hojas; sin embargo, ninguna de las especies las presentó. El dendrograma muestra claramente las relaciones de afinidad entre las especies de Colubrina, destacando la separación entre las secciones Cowaiana y Barcena del subgénero Colubrina, así como la distinción del subgénero Serrataria. Dentro de este último subgénero, se evidencian dos agrupamientos entre las especies incluidas. Finalmente consideramos que es crucial revisar Colubrina arborescens y determinar si los ejemplares recolectados en el sureste de México y en Guatemala son taxones distintos a los de las Antillas.

