











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los productos forestales se han convertido en un recurso estratégico del crecimiento económico de la humanidad, dado que son una fuente de madera para construcciones, pulpa para papel, madera aserrada, cajas y empaques (Food and Agriculture Organization of the United Nations [FAO], 2019). Las plantaciones forestales no solo pueden considerarse un mecanismo para satisfacer la necesidad mundial de madera, sino que también pueden valorarse como una táctica para la rehabilitación de terrenos degradados (Lanly, 2003). No obstante, la extracción de madera trae consigo desafíos ambientales como el llevar a cabo aprovechamiento forestal, permitiendo la conservación de remanentes de bosques y la rehabilitación gradual o total de los terrenos con aptitud forestal degradados (Pensado-Fernández et al., 2014).
Para el año 2021, Colombia produjo cerca de 2 238 981 m3 de madera proveniente de plantaciones forestales comerciales, en un área plantada de 528 855 ha (Ministerio de Agricultura y Desarrollo Rural, 2022). Dentro de las especies forestales más comerciales en el país se encuentran algunas introducidas como Acacia mangium Willd., Pinus patula Schl. et Cham., Pinus caribaea Morelet., Tectona grandis L. f., Eucalyptus pellita F. Muell., Pinus tecumanii Eguiluz & J.P. Perry., Gmelina arborea Roxb. ex Sm.; y algunas nativas como e Hevea brasiliensis Willd. ex A. Juss. Müll. Arg. Sin embargo, plantaciones de especies exóticas podrían no albergar la diversidad nativa de las diferentes zonas de vida del territorio colombiano. Por ejemplo, el P. patula se encuentra ampliamente distribuido por el territorio nacional, con alrededor de 59 204 ha, principalmente en la región del eje cafetero y el suroccidente (Ministerio de Agricultura y Desarrollo Rural, 2021). Aunque constituye una materia prima básica para muchos procesos industriales y múltiples subproductos (Ministerio de Agricultura y Desarrollo Rural, 2020), se presume que esta especie también ha causado impactos negativos sobre el suelo y la hidrología de los páramos, ocasionando variaciones en las condiciones microclimáticas y edáficas, las cuales a su vez pueden provocar cambios importantes en la estructura y diversidad de la vegetación (Ruiz, 2014).
El P. patula acumula una densa y gruesa capa de hojarasca de acículas ricas en lignina, la cual, en teoría, reduce o inhibe la germinación y el establecimiento de especies autóctonas (Tulande-Marín. et al., 2018). Asimismo, se ha sugerido que es posible que la presencia de un alto contenido de lignina en las acículas, así como de resinas y fenoles, pueden acidificar el suelo y afectar el establecimiento de otras especies (Ruiz, 2014) y, por tanto, el proceso de regeneración natural. La regeneración natural se concibe como un proceso gradual de recuperación de la estructura, la función y la composición del ecosistema, a las condiciones previas a la perturbación, con cambios en la vegetación que deberían estar acompañados de modificaciones en los microorganismos del suelo y la fauna (Chazdon y Guariguata, 2016). No obstante, si la composición química, física y microbiana del suelo cambia, podrían cambiar también las especies vegetales que lo ocupan debido a los niveles de tolerancia de las mismas (Edelaar et al. 2017), lo que podría verse reflejado en cambios de composición de las comunidades vegetales. Así, el estudio de especies indicadoras permitiría diferenciar los ensamblajes vegetales asociados a este tipo de cobertura, en comparación con los relictos de bosque nativo, facilitando la comprensión del impacto de esta especie exótica sobre la biodiversidad local.
Las especies indicadoras son organismos cuya presencia, abundancia o ausencia refleja condiciones ambientales específicas y permite evaluar el estado ecológico de un ecosistema (Dufrêne y Legendre, 1997). Así, la bioindicación (análisis de especies indicadoras) es fundamental en estudios de regeneración y conservación, ya que puede proporcionar información clave sobre los efectos de perturbaciones o cambios en el hábitat (González-Valdivia et al., 2011). No obstante, los estudios han centrado sus esfuerzos en comparar la diversidad en términos de riqueza de especies, prestando poca atención a los cambios en la composición y al recambio de especies en áreas donde P. patula está presente. Por ejemplo, Jadán et al. (2019) encontraron que la riqueza de especies fue mayor en los ecosistemas naturales, en comparación con las plantaciones de P. patula en Azuay, Ecuador. Sin embargo, la abundancia fue estadísticamente similar en los dos escenarios evaluados. Escenario contrario encontraron en Cotopaxi (Ecuador), donde los valores de diversidad fueron mayores, aunque no significativamente, para las zonas de páramo reforestadas con plantaciones de pino (Pinus radiata D. Don y P. patula), que en las zonas naturales (García-Macas, 2019).
En este contexto, el presente trabajo buscó responder las siguientes preguntas: ¿Cuál es la riqueza y los valores de diversidad verdadera (números de Hill) en el bosque nativo y la plantación de P. patula?; ¿Existen diferencias en la composición de especies (diversidad beta) entre el bosque nativo y la plantación de P. patula?, ¿Cuáles son las especies indicadoras en cada sitio?, ¿Cuál es la relación entre las variables ambientales (pH, cobertura del dosel, horizonte O) con los patrones de diversidad hallados en los dos relictos? Estas preguntas fueron basadas en la hipótesis de que las modificaciones en el hábitat, generadas principalmente por la acumulación de acículas del P. patula, reducen la diversidad de especies respecto a un relicto en regeneración natural, debido a la exclusión de aquellas especies no tolerantes a dichas condiciones.
Objetivos
El objetivo de este trabajo fue comparar la diversidad vegetal de especies vasculares arbóreas y arbustivas entre un relicto de bosque y una plantación de P. patula bajo regeneración natural de 30 años sin intervención. Además de evaluar la riqueza de especies (diversidad alfa), analizar el recambio de especies y los cambios en la composición (diversidad beta); y estimar la relación entre la estructura de la vegetación y las condiciones edáficas, incluyendo pH, hojarasca y cobertura del dosel, con el fin de comprender cómo las modificaciones generadas por la presencia de P. patula pueden influir en la regeneración y recuperación de especies nativas.
Materiales y métodos
Sitio de estudio
El estudio se llevó a cabo en las instalaciones del Centro de Investigación La Selva AGROSAVIA ubicado en el municipio de Rionegro, Antioquia, Colombia (06° 07’ 946” N y 075° 24’ 986” W), en un relicto de conservación de seis hectáreas, con altitud media de 2137 m, temperatura media de 17 °C, precipitación anual de 2525 mm y humedad relativa media de 75%, lo cual corresponde a la zona de vida bosque muy húmedo Montano Bajo (bmh-MB) (Holdridge, 1967). Se seleccionaron dos lotes como área de estudio: 1) una plantación abandonada de P. patula (en adelante, pino) de 25 598 m2, que tenía probablemente un objetivo comercial, pero que, por cambios en la vocación del predio, se dejó sin intervención de manejo por al menos 30 años; 2) un bosque nativo de 35 635 m2 (en adelante, nativo), ambos pertenecientes al “relicto de conservación de La Selva”.
Establecimiento de parcelas e identificación taxonómica
Las parcelas se delimitaron en campo, empleando la metodología de Gentry (1995)); cada parcela se subdividió en 10 subparcelas de 50 m × 2 m, separadas 2 m entre ellas, y por lo menos a un metro de los senderos de tránsito. Las subparcelas del bosque nativo fueron marcadas del 1 al 10 y las de la plantación de pino del número 11 al 20. Posteriormente, se midió la altura de los árboles seleccionados con una regla grabable manual, según la ecuación 1.
donde:
H : altura de árbol, por conocer
h : altura del observador en el instrumento
D : distancia conocida del árbol hasta el observador
d : distancia del instrumento hasta el ojo del observador
En la parcela se contabilizaron los individuos con un diámetro a la altura del pecho (DAP; diámetro a 1,3 m de altura del fuste con respecto al nivel del suelo) mayor o igual a 3 cm y/o con una altura total mayor o igual a 2 m. El DAP se calculó a partir de la circunferencia del árbol (cm), medida con cinta métrica, idealizando el fuste como un círculo perfecto.
Adicionalmente, se recolectaron muestras de herbario para su posterior identificación por comparación en el herbario de la Universidad Católica de Oriente (HUCO) y/o por clave taxonómica.
Caracterización ambiental de las parcelas
Se realizaron dos muestreos de suelos, uno en la parcela pino y otro en la parcela nativo, siguiendo la metodología descrita en Mendoza y Espinoza (2017). Con la ayuda de una pala y un machete, se despejó la zona donde se recolectó la submuestra de suelo; para ello se realizó una calicata de 25 cm × 25 cm y se separó la capa orgánica “Horizonte O” del suelo. Lo obtenido con la pala, se pesó en una báscula digital de 1 g de resolución y se introdujo en una bolsa sellable para su posterior transporte al laboratorio. Se recolectaron 10 submuestras de suelo, cada una con 100 g, para un equivalente aproximado de 1000 g de suelo por parcela. Al finalizar la recolección de cada una de las parcelas, se mezcló y rotuló cada una de las muestras con su respectiva información. Las muestras de suelo fueron enviadas a un laboratorio comercial de la zona para su posterior análisis, incluyendo conductividad eléctrica, capacidad de intercambio catiónico y elementos mayores y menores.
En campo, se midieron las variables: horizonte (O), pH y cobertura de dosel en cada una de las subparcelas, de cada parcela. Para el muestreo del horizonte, se hicieron cuatro calicatas distribuidas de manera uniforme, equidistantes entre sí y de los bordes del lote, en cada una de las subparcelas. Para ello se siguió la metodología para la descripción de suelos de la FAO (2009). En cada una de las subparcelas se generó una calicata de aproximadamente 30 cm de profundidad, que permitiera determinar la profundidad del horizonte, el cual fue medido con un flexómetro manual de 1 mm de resolución. Para la medición de pH del suelo en cada subparcela, se obtuvo una muestra de suelo usando un barreno holandés. Cada muestra tomada por parcela fue debidamente almacenada en bolsas sellables y rotulada con el número de la subparcela; posteriormente, en el laboratorio de la Universidad Católica de Oriente, se determinó el valor del pH de cada alícuota; para ello se utilizó agua destilada y un medidor de pH, siguiendo la metodología de Gómez-Giraldo (2013).
Cobertura del dosel
Para la medición de la cobertura de dosel, se empleó una lente de pescado ubicada sobre la cámara de un teléfono celular, con ajuste de nivel, colocada sobre el suelo y en modo profesional para conservar las configuraciones de apertura. Se tomaron cuatro fotos en cada una de las subparcelas de pino y nativo. Las fotografías se tomaron en cuatro puntos diferentes de la subparcela (uno al inicio, dos cercanas a la mitad de la subparcela según el mapa topográfico y la última al finalizar la subparcela); las fotografías se tomaron a la altura de dos metros del suelo, siempre con la lente hacia arriba; fueron nombradas con el código de la subparcela, siguiendo la metodología descrita en Garrido et al. (2017). Posteriormente, las fotografías se analizaron en la aplicación HabitApp© versión 1.1 (MacDonald y MacDonald, 2016), para calcular el porcentaje de cobertura del dosel. Los valores se promediaron para asignar el valor de cobertura del dosel a cada subparcela.
Análisis de datos
Para determinar riqueza, diversidad y abundancia de especies, se midió inicialmente la diversidad gamma para el sitio de estudio, la cual se considera como la riqueza de especies dentro de varios tipos de cobertura (Calderón-Patrón y Moreno, 2019). Posteriormente, se calculó la diversidad alfa para los dos lotes (pino y nativo), y se estimó el número esperado de especies a través de una curva de rarefacción y extrapolación-interpolación con base en la cobertura del muestreo, utilizando el método descrito por Colwell et al. (2012). Dichos análisis fueron implementados en la plataforma iNEXT, (Chao et al., 2016). Adicionalmente, se realizaron comparaciones de la riqueza y la abundancia, mediante la prueba no paramétrica MannWhitney.
Se calcularon los índices de diversidad en términos de números equivalentes (números de Hill): la diversidad de orden 0 (q0) o riqueza de especies; la diversidad de orden 1 (q1), que es el exponencial de la entropía del índice de Shannon; y la diversidad de orden 2 (q2), que es el inverso del índice de Simpson. Se considera que q1 hace referencia a la diversidad ponderada por la abundancia de las especies típicas, y que q2 incluye la diversidad únicamente de aquellas especies más importantes o abundantes (Cultid-Medina y Escobar, 2019).
Se calculó la diversidad beta siguiendo el método POD, propuesto por Podani y Schmera (2011) y seguido por Carvalho et al. (2012), en el cual la disimilitud de Jaccard es calculada mediante la ecuación 2.
donde:
a = número de especies compartidas entre dos sitios: A (nativo) y B (pino)
b y c =números de especies exclusivas para cada sitio
También se calculó la diversidad beta particionada, para comprender la disimilitud debida al recambio o al anidamiento, con la ecuación 3 (Calderón-Patrón y Moreno, 2019).
donde:
β cc = disimilitud total
β -3= disimilitud debida al reemplazo y
β rich = disimilitud debida a las diferencias en riqueza (anidamiento)
Adicionalmente, mediante la ecuación 4, se calcularon los valores de beta para datos de abundancia, siguiendo los índices propuestos por Baselga (2013).
donde:
beta.Bray-Curtis =disimilitud de Bray-Curtis
beta.Bray.Bal = componente de variación balanceada de la abundancia
beta.Bray.Gra = componente de gradiente de abundancia
Finalmente, se realizaron análisis de agrupación para estimar la diferencia entre las subparcelas, basados en los índices de Bray Curtis y Jaccard, donde la similitud entre dos comunidades se mide, respectivamente, a partir de la abundancia de las especies o de su presencia/ausencia.
Con el fin de establecer si existen especies indicadoras de cada ensamblaje, se llevó a cabo un análisis de especies indicadoras (IndVal%) para determinar qué especies podrían ser indicadoras de cada uno de los sitios. Esta metodología resulta clave para identificar a aquellos taxones asociados con mayor significancia a las distintas unidades del paisaje. Entre estos, destacan los vinculados al bosque (área no perturbada), el cual actúa como ecosistema de referencia en la zona (Dufrêne y Legendre, 1997). Las especies consideradas como características (indicadoras) de una condición de hábitat fueron aquellas con InVal≥50% (González-Valdivia et al., 2011).
Adicionalmente, se realizó un análisis de covarianza (Ancova) para evaluar, de manera indirecta, el efecto de las parcelas sobre el crecimiento de los árboles, en términos de su altura y DAP. En la Ancova, se consideró la altura (m) de los individuos como variable respuesta, y el DAP (cm) como variable explicativa; y la parcela (pino o nativo), como variable categórica o factor (Badii et al., 2008). Posteriormente, se realizó un análisis de ordenamiento multidimensional no métrico (NMDS, por sus siglas en inglés), basado en el índice de Bray-Curtis, para definir las diferencias de la composición de especies entre las parcelas con las variables ambientales (pH, Horizonte O y cobertura de dosel). Los valores medios para las parcelas de dichas variables ambientales se compararon a partir de pruebas Tukey (Tukey, 1977), empleando los datos de las subparcelas. Para la comparación del pH, fue necesario transformar los datos a su forma lineal (H+=10-pH ), antes de realizar el análisis.
Todos los análisis fueron llevados a cabo en el programa Past 4.16 (Hammer et al., 2001).
Resultados
Caracterización fisicoquímica de las parcelas
Los resultados de los análisis de suelos de las parcelas sugieren que el pH del suelo en la parcela pino es más bajo que el de nativo; asimismo, la presencia de aluminio se manifiesta solo en la parcela pino, pero no en nativo. Para el caso de los macronutrientes (P, K, Ca) y algunos micronutrientes (Mg, Co, Mn, Zn), se presentan en mayor cantidad en el bosque nativo. Por el contrario, Fe, Bo y S, se presentan en mayor cantidad en la parcela pino (Tabla 1).
TABLA 1 Comparación de variables químicas del suelo de una plantación de P. patula sin manejo y de un área de vegetación secundaria (bosque nativo) en Antioquia, Colombia.
| Determinación analítica | Pino | Nativo | Unidad |
| pH. | 4,57 | 5,50 | pH |
| Conductividad eléctrica | 0,28 | 0,05 | dS/m |
| Capacidad de intercambio catiónico | 7,05 | 2,54 | Cmol (+) /Kg |
| Fosforo | < 7,05 | 12,37 | mg/kg |
| Azufre | 6,28 | 5,90 | mg/kg |
| Boro | 6,22 | 0,02 | mg/kg |
| Aluminio intercambiable | 4,63 | 0,00 | Cmol (+) /kg |
| Calcio | < 0,59 | 1,49 | Cmol (+) /kg |
| Magnesio | 0,28 | 0,72 | Cmol (+) /kg |
| Potasio | 0,15 | 0,34 | Cmol (+) /kg |
| Sodio | < 0,14 | 0,00 | Cmol (+) /kg |
| Hierro | 206,44 | 73,6 | mg/kg |
| Cobre | 2,07 | 4,00 | mg/kg |
| Manganeso | 3,48 | 13,30 | mg/kg |
| Zinc | 2,56 | 7,50 | mg/kg |
Los valores para las condiciones físicas (conductividad eléctrica, capacidad de intercambio catiónico, pH y elementos mayores y menores), encontradas en el estudio, se resumen en la tabla 2. Entre las parcelas no se encontraron diferencias significativas entre el porcentaje del dosel (t = 1,2762; P = 0,21812), pero sí se encontraron diferencias significativas para la profundidad del horizonte O (t = 6,0115; P = 1,10E-05) y pH (t = 6,8995; P = 1,8844E-06).
TABLA 2 Condiciones físicas y de pH para las parcelas de bosque de una plantación de P. patula sin manejo y de un área de vegetación secundaria (bosque nativo) en Antioquia, Colombia.
| Variable ambiental | Estadístico | Nativo | Pino |
| pH | Min | 4,57 | 4,10 |
| Max | 5,03 | 4,47 | |
| Mediana | 4,77 | 4,30 | |
| Horizonte O | Min | 4,45 | 2,15 |
| Max | 8,28 | 5,08 | |
| Media | 6,65 | 3,58 | |
| Desv. est. | 1,22 | 1,06 | |
| Dosel % | Min | 71,50 | 64,25 |
| Max | 80,75 | 81,25 | |
| Media | 77,43 | 75,03 | |
| Desv. est. | 2,85 | 5,22 |
Riqueza y composición florística
En total, se encontraron 304 individuos (165 individuos en nativo y 139 en pino), pertenecientes a 15 familias y 19 especies, de las cuales se registraron 16 en el bosque nativo y 12 en la plantación de pino. Del total de especies encontradas, nueve se comparten (a) entre las dos parcelas, siete solo se encuentran en nativo (b), y tres en la plantación de pino (c) (Tabla 3). Al comparar los valores de diversidad por parcelas, se sugieren mayores valores promedio tanto para la riqueza (Fig. 1A), como para la abundancia (Fig. 1 B), pero sin diferencias significativas en las mismas (Riqueza: z =1,5187; P= 0,12884 Abundancia: z = 1,7112; P= 0,087051).
TABLA 3 Comparación de especies arbóreas y arbustivas registradas en parcelas de una plantación de P. patula sin manejo y de un área de vegetación secundaria (bosque nativo) en Antioquia, Colombia.
| Especie | Nativo | Pino | Biondicación | IndVal% | Valor p |
| Alchornea grandiflora Müll.Arg. | X | X | Pino | 57,44 | 0,0382 |
| Alchornea triplinervia (Spreng.) Müll.Arg. | X | Nativo | 50 | 0,0153 | |
| Bejaria aestuans L. | X | ||||
| Cecropia peltata L. | X | ||||
| Clidemia hirta (L.) D. Don. | X | ||||
| Clusia multiflora Kunth. | X | X | Nativo | 65,91 | 0,009 |
| Cyathea caracasana (Klotzsch) Domin. | X | X | Nativo | 74,36 | 0,0047 |
| Eugenia sp. | X | ||||
| Guadua angustifolia Kunth. | X | Pino | 60 | 0,0061 | |
| Hedyosmum racemosum (Ruiz & Pav.) G. Don. | X | X | |||
| Ladenbergia macrocarpa (Vahl) Klotzsch. | X | X | |||
| Miconia notabilis Triana. | X | ||||
| Nectandra acutifolia (Ruiz & Pav.) Mez. | X | Nativo | 90 | 0,0001 | |
| Persea caerulea (Ruiz & Pav.) Mez | X | X | |||
| Piper daniel-gonzalezii Trel. | X | X | |||
| Psidium cattleianum Sabine. | X | ||||
| Vasconcellea sp. | X | X | |||
| Vismia baccifera (L.) Planch. & Triana. | X | ||||
| Zyzigium jambos (L.) Alston. | X | X |
La presencia de cada especie en los distintos ecosistemas se indica con una "X". Además, se presentan los valores de bioindicación significativos, expresados como porcentaje de IndVal (%), junto con su respectivo valor de significancia estadística (Valor p).
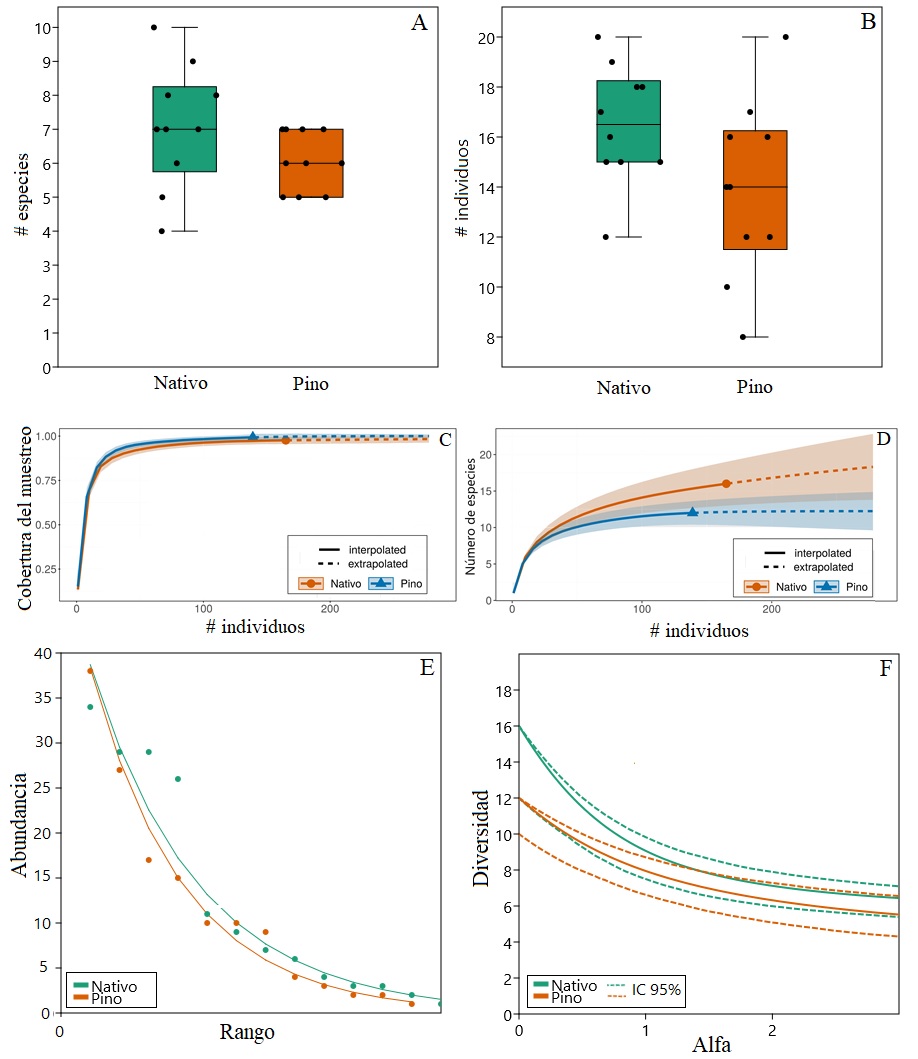
A: riqueza por subparcelas; B: abundancia vegetal por subparcelas. C: curva de extrapolación para cobertura del muestreo. D: curva de extrapolación para riqueza. E: curva rango abundancia. F: perfiles de diversidad basados en números de Hill (línea continua); líneas discontinuas denotan intervalos de confianza de 95%.
FIGURA 1 Comparación de la diversidad vegetal en parcelas de una plantación de P. patula sin manejo y de un área de vegetación secundaria (bosque nativo) en Antioquia, Colombia.
El muestreo realizado fue representativo, con una cobertura estimada de 99 % para el sitio pino y 98% para nativo, y muestra la misma tendencia asintótica ante una extrapolación hecha al doble de los individuos de la parcela menos abundante (pino) (Fig. 1C). De acuerdo con la estimación de especies, mediante la extrapolación, se sugiere un máximo de 18 especies (± 5,94, para intervalos de confianza de 95%), es decir, dos especies más que las encontradas. Para pino no se estima mayor número de especies si se incrementara al doble el esfuerzo de muestreo (± 2,79, para intervalos de confianza de 95%) (Fig. 1D).
Las comunidades de ambas parcelas mostraron patrones de dominancia similares (Fig. 1E), siguiendo un patrón de modelo geométrico. Asimismo, el perfil de diversidad (números de Hill) muestra un comportamiento similar; se observa que la riqueza es levemente mayor (q0), aunque sin diferencias significativas, para el bosque nativo, la diversidad ponderada por la abundancia relativa (q1) y la diversidad ponderada por las especies más abundantes (q2) fueron similares (Fig. 1F).
Composición
El análisis de composición sugiere una disimilitud total (βcc) de 52%, de la cual, 31% corresponde al reemplazo de especies (β-3) y 21% está dado por las diferencias en riqueza (βrich) entre las parcelas bosque y pino. Para los datos de abundancia, la disimilitud total (beta.Bray-Curtis) fue de 86,02%, representada por 82,53% de variación balanceada (beta.Bray.Bal) y tan solo 3,49% por el gradiente de abundancia (beta.Bray.Gra). El mismo patrón se vio reflejado en los análisis de agrupamiento, los cuales muestran una gran similitud en composición entre subparcelas evaluadas al interior de las parcelas, tanto para las distancias basadas en la abundancia (Bray-Curtis, Fig. 2A), como para las distancias basadas en presencia/ausencia (Jaccard, Fig. 2B).
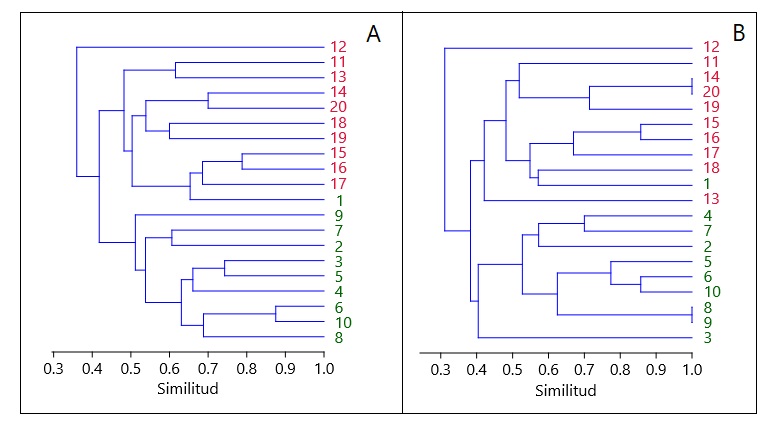
A: agrupamiento calculado por distancias de Bray-Curtis, basadas en abundancias. B: agrupamiento calculado por distancias de Jaccard para datos basados en presencia y ausencia.
FIGURA 2 Análisis de agrupamiento para las subparcelas evaluadas al interior de parcelas de bosque nativo (verde) y plantación de P. patula sin manejo (rojo) en Antioquia, Colombia.
El análisis de especies indicadoras sugiere significancia para dos especies en pino: Alchornea grandiflora (IndVal% = 57,44; P = 0,0382) y Guadua angustifolia (IndVal% = 60,00; P = 0,0061); y para cuatro de nativo: Alchornea triplinervia (IndVal% = 50,00; P = 0,0153), Clusia multiflora (IndVal% = 65,91; P = 0,009), Cyathea caracasana (IndVal% = 74,36; p = 0,0047) y Nectandra acutifolia (IndVal% = 90,00; P = 0,0001) (Fig. 3).
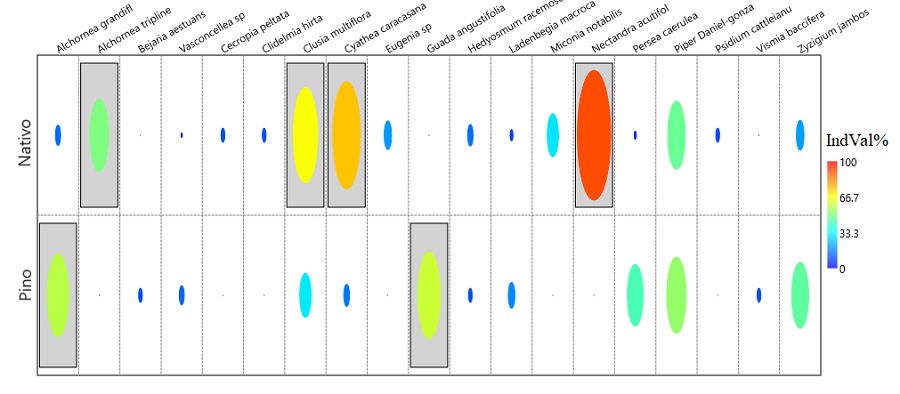
Relación de las especies y las variables ambientales
En el NMDS (Fig. 4A), se muestra la ordenación de las especies según su abundancia (Bray-Curtis), con un valor de estrés de 0,17 (Fig. 4B). La relación entre el pH y el Horizonte O fue positiva y significativa (r = 0,68; P = 0,00085) (Fig. 4C).
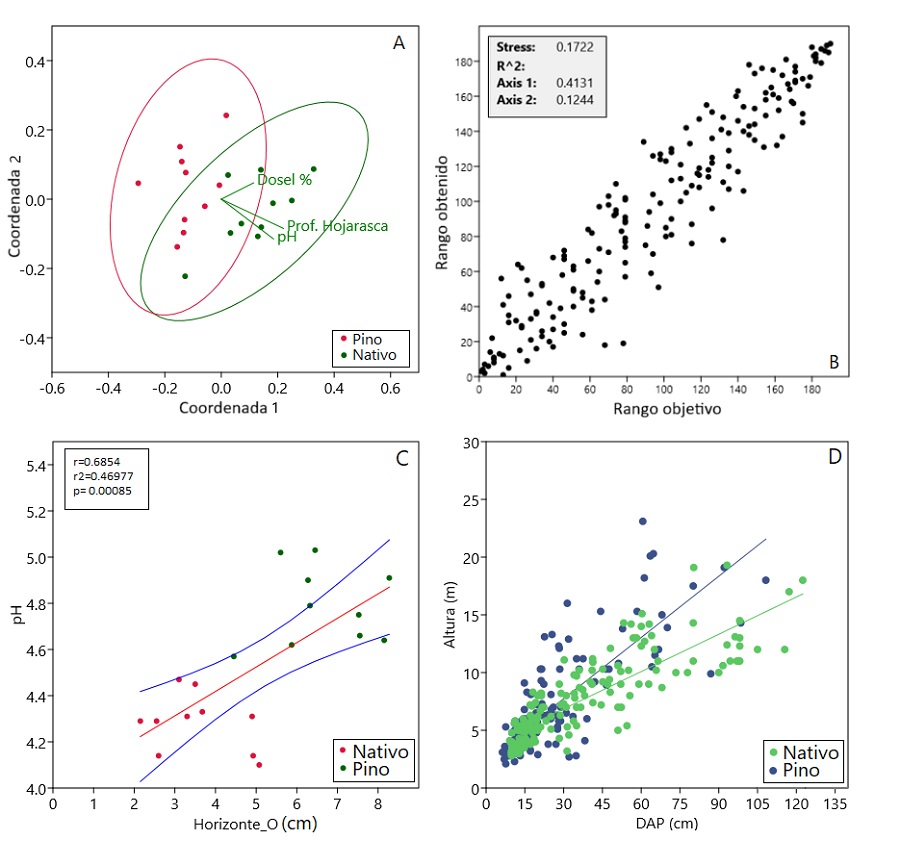
A: Análisis de ordenamiento multidimensional no métrico (NMDS) por distancias de Bray Curtis. B: gráfico de dispersión para el desempeño del modelo asociado a los ejes de ordenación del NMDS. C: Relación entre la profundidad del Horizonte O y el pH de las parcelas de pino. (intervalos de confianza del 95% en líneas azules). D: Análisis ANCOVA para la relación del DAP (cm) y la altura (m).
FIGURA 4 Análisis de variables ambientales en las subparcelas de una plantación de P. patula sin manejo y de un área de vegetación secundaria (bosque nativo) en Antioquia, Colombia.
Al comparar la relación entre el DAP y la altura de los individuos registrados, se observó una tendencia positiva en ambas parcelas, donde el incremento en el DAP se asoció con un aumento en la altura (Fig. 4D). Sin embargo, los análisis revelaron diferencias significativas entre las parcelas (F = 7,906; p = 0,005), donde la plantación de pino presentó una altura media superior y una mayor proporción altura-DAP (pendiente = 0,555) en comparación con el bosque nativo (pendiente = 0,338)
Discusión
Los productos forestales han sido un recurso estratégico en el desarrollo económico de la humanidad (FAO, 2019). Sin embargo, las plantaciones forestales de especies exóticas pueden traer consecuencias graves en la conservación de la diversidad nativa en los ecosistemas (Pensado-Fernández et al., 2014). En este trabajo se comparó la diversidad vegetal de una parcela de bosque nativo y la de una plantación de pino sin mantenimiento y en proceso de regeneración natural, para evaluar los cambios en la comunidad y algunas variables ambientales. Los resultados sugieren que, aunque la riqueza de especies no difiere estadísticamente entre ambas parcelas, sí se presentan diferencias importantes en cuanto a su composición florística. Esto no solo se observa en las especies (reemplazo, β-3), sino también en el balance de las especies más abundantes en cada parcela (beta.Bray.Bal); es decir, las especies más abundantes en la parcela pino no son las más abundantes en la parcela de bosque nativo. Lo anterior, fue soportado también por las especies indicadoras asociadas a cada parcela.
En teoría, los ecosistemas naturales presentan una mayor riqueza y diversidad que las plantaciones de pino (Jadán et al., 2019). No obstante, el abandono del manejo silvicultural en la plantación forestal podría haber favorrecido la colonización y establecimiento de especies con tolerancia diferencial a las condiciones de cobertura modificadas por el pino, lo cual se vería reflejado en los cambios de composición con respecto a la parcela de bosque nativo. Por ejemplo, las especies con mayores valores de bioindicación en la parcela pino fueron A. grandiflora (57,44 %) y G. angustifolia (60,00%), las cuales son reconocidas por adaptarse a las condiciones medioambientales y edáficas que proporciona la plantación de pino. A. grandiflora, según Guariguata et al. (2002), es pionera dentro de una regeneración natural y podría contribuir a la conformación de un bosque primario. Adicionalmente, también se conoce que esta especie se adapta muy bien en bosques de galería y bosques secundarios maduros (Vargas, 2002). Asimismo, G. angustifolia se adapta muy bien a suelos con pH entre 5,5 y 6; sin embargo, puede establecerse en suelos con valores de pH inferiores, aunque su buen desarrollo puede verse afectado. No obstante, debido a su uso en plantaciones comerciales, es altamente probable que su presencia en este sitio no sea resultado de un proceso de dispersión natural (Casanova-Alvino et al., 2019).
Para el bosque nativo, las especies indicadoras fueron A. triplinervia, C. multiflora, C. caracasana y N. acutifolia. A. triplinervia es una especie arbórea que ha sido registrada en los inventarios florísticos como una especie dominante; se conoce por su capacidad de regeneración en bosques degradados, y sus frutos sirven de alimento para algunos animales (Granato-Souza et al., 2018). También ha sido registrada para el departamento de Antioquia en ambas cordilleras, mostrando su adaptabilidad principalmente entre 1700 m y 2700 m s.n.m., creciendo en bosque premontano (b-PM) y bosque húmedo montano (bh-M) (García-Ruenes y Álzate-Guarín, 2021). De igual forma, el helecho arbóreo C. multiflora, ha sido señalado en otras investigaciones como una de las especies con mayor representación en bosques nativos (Cortés-Ballén et al., 2020) y se encuentra distribuida entre los 1300 m y 3400 m s.n.m.; en Colombia se ha registrado mayormente en los Andes centrales antioqueños y crece sobre puntos o abrigos monoclimáticos húmedos (Bonilla, 2002). Respecto a C. caracasana, se sabe que la luz es un factor importante para la germinación de sus esporas, dado que estas se dispersan en áreas abiertas y pueden germinar en condiciones nubladas y lluviosas (Hiendlmeyer y Randi, 2007). No obstante, es probable que no sobrevivan a los días soleados con sol directo, en condiciones de menor dosel como la parcela pino. Es de considerar que los helechos son un importante elemento en la biodiversidad de los trópicos, cumplen funciones ecológicas importantes en la dinámica de ecosistemas estratégicos y la recuperación de áreas degradadas, actuando como pioneras en la sucesión vegetal (Giraldo y Mejía, 2003). Por otro lado, N. acutifolia es una especie arbórea ampliamente distribuida en Antioquia, desde el nivel del mar hasta los 2500 m s.n.m. (Callejas y Idárraga, 2013); ha sido registrada como una de las especies con mayor peso ecológico en un estudio realizado en plantaciones forestales de Pinus caribaea y Eucalyptus pellita, establecidas en Villanueva, Casanare, Colombia (Fernández et al., 2012). N. acutifolia crece en pendientes y valles andinos, se distribuye desde Colombia hasta Bolivia, está presente en bosques maduros y bosques secundarios (Toro-Murillo, 2021).
Desde el punto de vista ambiental, variables como pH y horizonte O son factores ambientales que podrían verse afectados por especies del género Pinus; en teoría, estas especies liberan acículas que se acumulan en forma de “mulching” ocasionando baja retención de agua y modificaciones en el pH (Ilek et al., 2024). Asimismo, se ha sugerido que la materia seca residual de P. patula en plantaciones de restauración en Colombia tiende a ser significativa, y su descomposición puede ser más lenta en comparación con la de otras especies debido a su alto contenido de lignina y polifenoles (León y Osorio, 2014). Dichas modificaciones podrían impactar negativamente el establecimiento de especies vegetales nativas, debido a su poca adaptación a dichos ambientes (Fernández-Núñez, 2008). En este trabajo, las variables ambientales pH, cobertura del dosel y horizonte O, fueron mayores en el bosque nativo y, probablemente, están asociadas con la menor tolerancia de sus especies indicadoras a estas condiciones. Por otra parte, Moretti et al. (2019) demostraron cómo la radiación es una variable clave para la supervivencia y crecimiento de las plantas, y cómo los niveles de cobertura del dosel regulan la cantidad de luz que llega al sotobosque, afectando de forma directa el crecimiento y desarrollo de muchas especies, siendo así obligadas a tener que competir por espacio y luminosidad. No obstante, en este estudio no se presentó diferencia significativa respecto al dosel entre parcelas. Asimismo, al realizar la comparación de la altura con el DAP de cada uno de los individuos registrados, para ambas parcelas se encontró que a medida que incrementa el DAP aumenta la altura del árbol, pero es mayor la altura en la plantación de pino. Esto podría obedecer al comportamiento fisiológico de la especie, que es introducida y que, por las condiciones del trópico, favorece eficientemente su desarrollo (García et al., 1999). Finalmente es interesante anotar que, según los análisis de suelos, la conductividad eléctrica de la parcela pino fue notablemente mayor (0,28 dS/m) que la de nativo (0,05 dS/m). Sin embargo, Mancheno (2011), en un estudio realizado en el Parque Nacional Cajas, al sur de Ecuador, no encontró diferencias significativas en la conductividad eléctrica ni en la calidad de los suelos al comparar plantaciones de pino con Polylepis reticulata Hieron. Cabe señalar que la falta de réplicas en este estudio para estas variables limita el alcance de las conclusiones al respecto. Esto resalta la necesidad de ampliar la recolección de datos sobre las características fisicoquímicas del suelo en las plantaciones de P. patula, para comprender mejor las variaciones en la conductividad eléctrica y su impacto en los ecosistemas y las plantas nativas de los Andes tropicales.
Los resultados aquí obtenidos sugieren que la plantación de pino, a través del abandono de tareas de manejo silvicultural, ha permitido la colonización de especies adaptadas a las condiciones modificadas por el pino; mientras que en el relicto natural, las condiciones ambientales parecieran favorecer la presencia y abundancia de especies indicadoras, típicas de bosques nativos. Las variaciones en el pH, el horizonte O y la conductividad eléctrica del suelo sugieren que las plantaciones de pino pueden alterar significativamente las condiciones edáficas, lo que podría afectar la regeneración y conservación de la biodiversidad nativa no tolerante a dichas modificaciones. Estos hallazgos sugieren la necesidad de continuar investigando el recambio de especies vegetales nativas en áreas cuyo historial forestal afectó las condiciones del suelo.
Conclusiones
Se evidenció que entre la parcela pino y la parcela nativo no existen diferencias respecto a la riqueza de especies, pero sí respecto a su composición, con dos especies fuertemente asociadas a la parcela pino (A. grandiflora y G. angustifolia) y cuatro especies a la parcela nativo (A. triplinervia, C. multiflora, C. caracasana y N. acutifolia). Por otra parte, los valores para las variables ambientales como pH y horizonte O fueron menores en la parcela pino, sugiriendo que dichos son factores se ven afectados por la especie dominante, P. patula, lo que afecta la fertilidad del suelo. Finalmente, los niveles de cobertura del dosel parecen no tener influencia de forma directa el crecimiento y desarrollo de las diferentes especies en cada parcela. Así, dadas las interacciones tróficas promovidas por el recambio en la diversidad vegetal en ambientes modificados con el P. patula, es necesario ampliar los estudios de diversidad a otros niveles tróficos en dichos ecosistemas. Esto permitirá comprender, de mejor manera, la dinámica de las comunidades en los Andes tropicales y las implicaciones a largo plazo de las plantaciones forestales, que incluyen especies exóticas en estos ecosistemas.

