











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCTION
Northwest of Los Reyes Metzontla, located to the south of the Tehuacán, Zapotitlán, and San Juan Raya sites, is a famous area for the abundance and good preservation of the invertebrate fossils found there. Nyst and Galeotti (1840) carried out a first paleontological study on invertebrate (mollusks and echinoderms) of the Jurassic in Tehuacán. This dating has subsequently been considered as erroneous, and some authors attributed these fossils to the Cretaceous such as d’Orbigny (1850) who defined them as Senonian; other authors considered them as belonging to the Aptian (Early Cretaceous) as Müllerried (1933). The echinoderms were analyzed, by Desor (1858- 1859), who assigned them a Jurassic age. De Loriol (1876) studied some echinoderms collected in San Juan Raya. Later, Cotteau (1890) reviewed the echinoderms from the same locality, and identified species which were like those found in the Aptian of France, as well as other species described for the first time “et qui paraissent appartenir également á l’étage Aptien”. After, Buitrón (1970) carried out the taxonomic study of fourteen species of echinoids of Lower Cretaceous from San Juan Raya-Zapotitlán (Puebla).
The ostreids were reported by Coquand (1869). Hernández Ocaña et al. (2015) made a taphonomic and paleoecologic study of the ostreids from San Juan Raya Formation. A gastropod fauna from Zapotitlán was described by White (1881). Felix and Lenk (1891) described mollusks, corals, and echinoderms from San Antonio de las Salinas.
A systematic analysis of the invertebrate fauna was conducted by Alencáster (1956) from the San Juan Raya region. González-Arreola (1974) reported the ammonite Phylloceras from this same locality.
Studies on marine invertebrate fauna, mainly bivalves, have been reported by Alencáster and Hernández Láscares (1991) in the Reyes Metzontla region.
Cantú Chapa (1993) described a new ammonite, Neocomitidae, dating it as Hauterivian, which was collected in the Santiago Coatepec area. Recently, Alencáster et al. (2009) recorded the bivalve Trichites pueblaensis in the same area.
With respect to the presence of microfossils, Lozo (1943) registered an assemblage composed of foraminifera from the upper Aptian, represented by some species of the genus Pseudocyclammina? as well as Choffatella, Flabelammina, Ammobaculites, Nodosaria, Dentalina, and Textularia, associated with Ostracoda from the Phylloceras rioi Ammonite Zone. Maync (1950) cited the presence of Choffatella decipiens in the upper Aptian of the San Juan Raya Formation. A Kimmeridgian larger foraminifera and algae association study from the Santiago Coatepec region was made by Omaña and González Arreola (2008).
The objective of the present study is to document the presence of a Late Jurassic larger foraminiferal assemblage identified along a rock section located NW of Los Reyes Metzontla, as a contribution to the knowledge of this group in Mexico, in addition to determining the age and the environmental deposit of these rocks, as well as the paleobiogeographic distribution.
GEOGRAPHICAL AND GEOLOGICAL SETTING
The studied section is located NW of Los Reyes Metzontla in southeast Puebla state. This area belongs geographically to the Sierra Madre del Sur Province, characterized by a tectonic evolution with folding, normal, and thrust faults, as well as intrusive and volcanic events, all included within a complex structure. According to Centeno-García et al. (2009), upon the old schist basement (Acatlán-Oaxaca Complex), the first sedimentary unit is the continental Matzizi Formation named informally by Calderón García (1956), which consists of sandstone interstratified with dark shale, containing scarce conglomerate lenses and coal layers with a high content of fossil plant remains of late Carboniferous and Permian age (Silva-Pineda, 1970; Carrillo and Martinez Hernández, 1981; Weber, 1997; Velasco de León et al., 2015; Flores-Barragán et al., 2019).
Above the Matzitzi Formation, a rock succession of red layers rests unconformably, which were informally referred to by the same author as “red beds.” This is constituted by conglomerates and reddish feldspathic quartz sandstone. The succession was deposited in a fluvial environment, apparently as an alluvial fan, with incipient stratification, and hummocky-like crosses observed in the sandstone horizons. The age of the “red beds” is uncertain, but because of their stratigraphic position, it has been dated as Jurassic (Morán Zenteno et al. 1993).
According to Centeno-García et al. (2009) “the red beds lying conformably on Cretaceous marine rocks (Zapotitlán and San Juan Raya Formations) consist of quartz sandstone, feldspars and lithics, with a calcareous matrix, interstratified with conglomeratic lenses formed by clasts of schist, gneiss and granite. The succession also includes intercalations of calcareous strata with fossils of gastropods, pelecypods and corals, and horizons of marls; this succession was dated as Barremian-Aptian (Alencaster, 1956; Buitrón and Barceló-Duarte, 1980; Hernández Láscares and Buitrón-Sánchez, 1992)”.
MATERIAL AND METHODS
The studied samples were collected from a section located NW of Reyes Metzontla town (18°14'46.7'' N, 97°30'41.7'' W), southeast Puebla state (Figure 1).
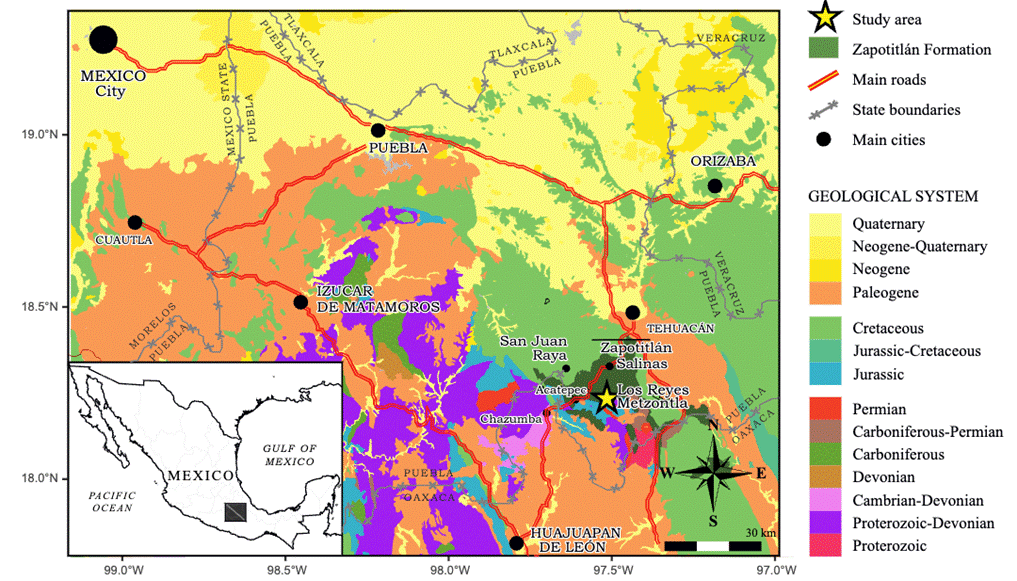
The sedimentary succession in the studied section mostly consists of limestone with intercalations of sandstone composed of quartz, feldspar and components lithics into a calcareous matrix (Figure 2).
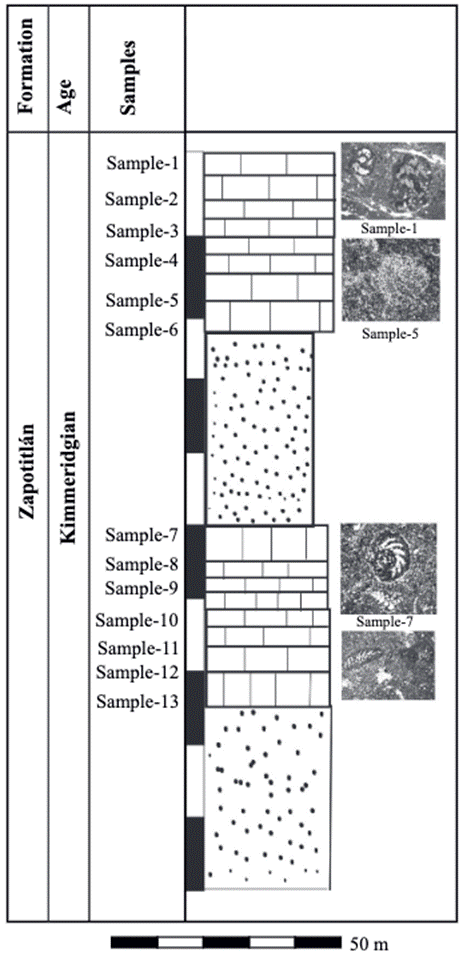
Figure 2 Simplified lithostratigraphic column of the Los Reyes Metzontla, section with position of samples and distribution of benthic foraminifera.
The limestone samples were processed and analyzed by means of thin sections. The larger benthic foraminifera was identified for establishing the age (Hottinger,1967; Pop and Bucur 2001; Bucur and Săsăran, 2005; Hughes, 2004 a), as well as a tool for interpreting the paleoenvironment (Hohenegger, 1999; BouDagher-Fadel, 2008). The paleobiographic distribution of these fossils was also reviewed.
RESULTS
Foraminiferal assemblage
The analyzed samples contain an association of larger foraminifera composed of Pseudospirocyclina mauretanica Hottinger, Choffatella tingitana Hottinger, Pseudocyclammina lituus (Yokoyama), Everticyclammina virguliana (Koechlin), E. praekelleri Banner and Highton, Rectocyclammina chouberti Hottinger, Mesoendothyra croatica Gušić, Alveosepta jaccardi (Schrodt, 1894), Nautiloculina oolitica Mohler, Siphovalvulina variabilis Septfontaine, and Coscinoconus alpinus (Leopold) (Figures 3 and 4). The macroinvertebrate fauna consists of bivalves, such as Trichites sp., and and the stromatoporid Cladoropsis mirabilis Felix.
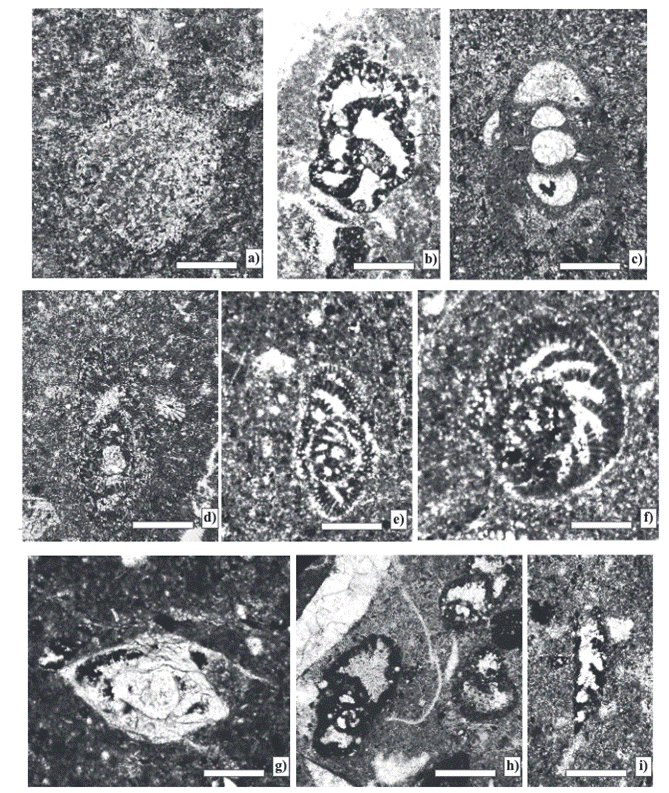
Figure 3 Larger foraminifera from the Zapotitlan Formation. a) Pseudospirocyclina mauretanica Hottinger. Equatorial section: test planispiral flabelliform with an alveolar hypodermis. (Sample-5c). b) Everticyclammina virguliana (Koechlin). Equatorial section: test planispiral with an alveolar wall (Sample-9). c) Nautiloculina oolitica Mohler. Axial section: test planispiral with septa doubled by addition of a second wall layer (Sample-5c). d) Pseudocyclammina lituus (Yokoyama). Axial section: test planispiral, wall coarsely alveolar and labyrinthic (Sample-5c). e) Alveosepta jaccardi (Schrodt). Subequatorial section: test planispiral with a streptospiral early stage. Wall finely and complex alveolar (Sample-7). f) Alveosepta jaccardi (Schrodt). Equatorial section: test planispiral, with wall complex alveolar (Sample-6). g) Lenticulina sp. Axial section: test planispiral, involute and biconvex with a keel, calcareous wall (Sample-8). h) Mesoendothyra croatica Gušić. Axial section: test coiled, early stage streptospiral, later becoming planispiral asymmetrical (Sample-5c). i) Rectocyclammina chouberti Hottinger. Subaxial section: test elongate, early stage planispiral, later uncoiling and rectilinear (Sample-5c). Scale bar 0.5 mm.
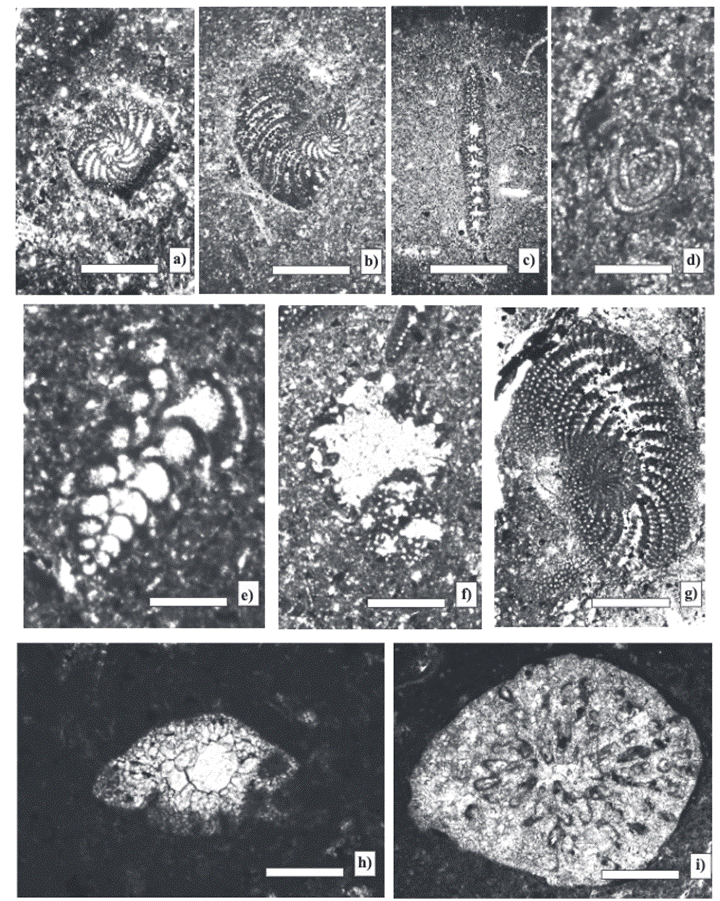
Figure 4 Larger foraminifera from Zapotitlán Formation. a) Chofatella tingitana Hottinger. Equatorial section: test planispiral, chambers numerous broad, and subepidermal network (Sample-2). b) Chofatella tingitana Hottinger. Subequatorial section (Sample-1). c) Chofatella tingitana Hottinger. Axial section (Sample-1). d) Glomospirella sp. Test: early part strongly twisted in a glomospiral shape upon a plane with intercalated planispiral whorls (Sample-5). e) Siphovalvulina variabilis Septfontaine. Tangential section: test triserial with connections between the chambers and canaliculate wall (Sample-7). f) Everticyclammina praekelleri Banner and Highton. Equatorial section: test planispiral with an alveolar wall (Sample-7). g) Chofatella cf. Ch. decipiens Schlumberger. Subequatorial section: test planispiral with a wall finely and complexly alveolar (Sample-4). h) Coscinoconus jurassicus (Henson). Axial section: test large with base conic pustular, and completely infilled umbilicus (Sample-13). i) Cladoropsis mirabilis Felix. Transversal section: skeletal elements and a differentiated cortex (Sample-12). Scale bar 0.5 mm.
Age
The benthic larger foraminifera are biostratigraphically significant in shallow water environments, where they are the most important fossils, and where other markers such as ammonites and calpionellids are not present.
The stratigraphic value of the larger foraminifera in the peri- Tethyan basins, as well as the comparison with the biostratigraphic ranges based on them, enable the foraminiferal associations contained in the studied samples to be dated. The most important age indicators will be examined below, suggesting a Kimmeridgian age to the analyzed succession.
Pseudospirocyclina mauretanica was described by Hottinger (1967, p. 73) from the upper Kimmeridgian-Portlandian of east flank of the Kerker Range, topographical chart Tistoutine Morocco. Later, this species was reported in Spain in the Asturias region (González Fernández et al., 2014).
Choffatella tingitana. This species was reported for the first time by Hottinger (1967, p. 65) from South Gebel Ahmar Lakdar, northern part of Marocco from levels considered of Kimmeridgian-Portlandian age. This foraminifer has been recorded from Spain as Ch. cf. Ch. tingitana in Asturias region by Gonzalez Fernández et al. (2014) and in the Teruel province of Tithonian age (Campos-Soto et al., 2016). Adefris et al. (2021) recorded Chofatella tingitana in the Kimmeridgian-Tithonian Hagere Selan section of the Bekele Basin in Ethiopia.
At Santiago Coatepec, Puebla (Mexico) it was identified in the Kimmeridgian (Omaña and González Arreola, 2008).
Pseudocyclammina lituus was described for the first time by Yoko- yama (1890, p. 26) as Cyclammina lituus from “Sehr häufig Kompira auch selten zu Torinosu” in Japan.
Yabe and Hanzawa (1926) made a revision of material collected near the Torinosu Limestone and proposed the new genus Pseudo- cyclammina based on thin sections of this species. The illustrations of Maync (1952, 1959) agreed with the reported form by Kobayashi and Vuks (2006) and Kobayashi and Wernli (2013) recordering the species in the Tithonian-Berriasian from the same locality in Japan.
Hottinger (1967) documented the occurrence of P. lituus in the Kimmeridgian-Portlandian interval in Morocco. Fourcade and Neumann (1966) and Fourcade (1971) reported it from the Kimmeridgian of the Albacete province (Spain).
In Croatia, this fossil occurs in an association dated as latest Oxfordian to earliest Kimmeridgian (Velić et al., 2002). it has been also reported in the Oxfordian-Berriasian Cieszyn Beds in the Polish Outer Carpathians (Olsewska, 2005), in the upper Tithonian Plassen carbonate platform in the northern Calcareous Alps (Schlagintweit et al., 2005), in the upper Kimmeridgian-Valanginian of southwestern Bulgaria (Ivanova et al., 2008), and in the Oxfordian-Berriasian of the southern part of the Crimea Mountains in southern Ukraine (Krajewski and Olszewska, 2007).
Velić (2007) reported Pseudocyclammina lituus associated with Everticyclammina virguliana, Alveosepta jaccardi, Siphovalvulina variabilis and Trocholina alpina from the Kimmeridgian.
Alveosepta jaccardi was first named as Cyclammina jaccardi by Schrodt (1894, p.734) of the Astartien from Col des Roches in Neuchatel (Switzerland) locality. It was later re-described by Maync (1958) as Pseudocyclammina jaccardi Schrodt=Pseudocyclammina, Chofatella sequana.
This species is widely distributed in the Tethys region, and therefore has a significant use for correlation. It has been reported by Pélissié and Peybernès (1982, p. 111), from the Oxfordian-Lower Kimmeridgian beds in Causses du Limogne, Quercy, NE Aquitaine, France; later Pelissié et al. (1984) considered it as lower Kimmeridgian age. Pop and Bucur (2001) regarded this species as late Oxfordian- early Kimmeridgian from the Valcăn Mountains (south Carpathians). Septfontaine (1988) considered this species as upper Oxfordian- Kimmeridgian. Clark and Boudagher-Fadel (2001) recorded it from Oxfordian-middle Kimmeridgian beds in Lebanon. Bucur and Săsăran (2005, p. 29) and Bucur et al. (2010) recorded deposits from Trascău Mountains (Romania), containing Alveosepta jaccardi of late Oxfor- dian-early Kimmeridgian age. Márques and Oloriz (1989) reported this species in Kimeridgian beds of the Algarve region in Portugal and in Turkey (Altiner, 1991; Tasli, 1993) and Morocco (Hottinger, 1967). In the Agadir Basin in Morocco, Bouaouda et al. (2004) situated the first occurrence A. jaccardi in the middle Oxfordian, extending up to the Kimmeridgian.
Alveosepta jacardi, Pseudocyclammina lituus, and Redmondoides lugeoni are present in the Hanifa Formation (Saudi Arabia) dated as mid-late Oxfordian to early Kimmeridgian age. The upper Jubaila Formation (Saudi Arabia) contains Alveosepta jaccardi, Kurnubia pa- lastiniensis, Trocholina palastiniensis, Coscinoconus alpina, suggesting a late Kimmeridgian (Hughes, 2004 a, 2009, 2018).
Alveosepta jaccardi has been recorded from deposits of the Sarv Abad section, spanning the Oxfordian to Early Kimmeridgian age by Sarfi and Yazdi-Moghadam (2016). Later Sarfi et al., (2023) indicated that the Alveosepta jaccardi Zone from the Mozduran Formation is da- ted as late Oxfordian to Kimmeridgian both studies carried out in Iran. Everticyclammina virguliana, identified in our material, was des- cribed by Koechlin (1942) from the Kimmeridgian; the type specimens come from the Berner Jura. Hottinger (1967) reported this species in the Kimmeridgian of eastern Morocco. This species is recorded in Portugal where its first occurrence is at the top of the Oxfordian (Ramalho, 1971). Banner and Highton (1990) dated it ranging from Kimmeridgian to “Portlandian” (Tithonian).
Everticyclammina praekelleri was described by Banner and High- ton (1990) in Broumana, near Beirut, Lebanon, from limestones dated by macrofossils to be Kimmeridgian-Tithonian in age (Dubertret, 1963). These authors considered that Everticyclammina praekelleri is of the Late Jurassic (Tithonian and possible Kimmeridgian) and a direct ancestor of the E. kelleri from earliest Cretaceous (Berriasian- Valanginian). Krajewski and Olzewaska (2007) recorded it in the Kim- meridgian and Tithonian in southern Crimea. This species has been identified by Pleş et al. (2015) in the Kimmeridgian-Tithonian from the central part of the Metaliferi Mountains (W Romania); Mircescu et al. (2016) indicated that E. praekelleri has a stratigraphic distribution from the Lower Kimmeridgian to upper Tithonian on the Getic Platform in the southern Carpathians (Romania).
Rectocyclammina chouberti was described for the first time by Hottinger (1967, p. 55) of lower and middle Kimmeridgian from Beni Bou Yahi topographical chart Tistoutine (Eastern Morocco). It has been recorded from different localities; for instance, in the Bikfaya Forma- tion from Lebanon (Clark and Boudagher-Fadel, 2001). Ivanova and Koleva-Rekalova (2004) recorded this species in the Kimmeridgian Slivnitsa Formation in southwestern Bulgaria. Later, Ivanova et al. (2008) registered this species from the Oxfordian-Kimmeridgian in the Velinovo section, in the same region.
Mesoendothyra croatica.Gušić (1969, p. 65) described this species, in the northern part of the Velika Kapela Mountain (Croatia). Later, Pélissié and Peybèrnes (1982) recognized two forms, a small one that appears in the strata between the Bajocian and Bathonian, and another larger one (equatorial diameter 1 mm), which is frequent in the Callovian-Oxfordian; however, Tasli (1993) reported this species in the Alveosepta jaccardi Biozone (upper Oxfordian-lower Kimmeridgian). Siphovalvulina variabilis was identified from the Callovian of Zuckerspitz in Switzerland (Septfontaine, 1980). Gale et al. (2018) stated that “the stratigraphic range of Siphovalvulina variabilis is uncertain due to the problems of nomenclature.” Tasli (2001) reported Siphovalvulina sp. from the Oxfordian-Kimmeridgian of the Central Taurides. Clark and BouDagher-Fadel (2001) recorded it from Callovian to Oxfordian strata of Lebanon. Schlagintweit et al. (2005) reported this species from the Kimmeridgian-Tithonian of Saltzkammergut, Austria; its stratigraphic range would be from the middle Jurassic to Tithonian (Krajewski and Olszewska, 2007). Adefris et al. (2021) registered this species from the Kimmeridgian to Tithonian at Mekelle Basin, Northern Ethiopia. Gale et al. (2018) concluded that the stratigraphic distribution ranged from Sinemurian to the Tithonian.
Paleoenvironment
In the studied samples, the occurrence of the larger foraminiferal assemblages has been very useful for interpreting the paleoenvironment as well as the limestone texture.
Among the factors that allow this group to flourish is water depth, which is probably the most significant factor that influenced the distribution of these fossils, as has been shown in various studies from Recent environments (Hallock and Glenn, 1986; Hottinger, 1997; Hohenegger, 2000, 2004). Water depth is associated with physical factors such as light penetration, temperature, energy, turbidity, nutrient availability, and substrate consistency, which are controlling the structure of these communities, so this information is very useful for inferring the paleoenvironment, supposing that these same factors affected the larger foraminiferal fossil communities (BouDagher-Fadel, 2008).
The later Jurassic was long considered to have been a warm greenhouse period (Valdes and Sellwood, 1992; Price and Sellwood, 1997; Sellwood et al., 2000; Sellwood and Valdes, 2006); however, recently an ice house cool event has been postulated, related to the short-term influx of cooler boreal water masses, based on oxygen isotope values from calcareous shell materials of marine organisms such as belemnite rostra and bivalve shells such as Trichites at the Subboreal Saxony Basin (Zuo et al., 2019).
Much of the Jurassic sediment consists of warm shallow-water carbonate successions, which are mostly of biogenic origin and contain larger foraminifera, algae, corals and stromatoporids as Cladoropsis mirabilis Felix.
With respect to the larger foraminifera identified in our material as Alveosepta and Choffatella, Banner and Whittaker (1991) indicated that present narrow alveolus and frequently labyrinthic hypodermis. These forms seem to have inhabited deeper paleoenvironments with reduced illumination and tolerated water rich in sediments in suspension, as in samples studied, while their contemporaneous forms with larger alveoles and irregularly labyrinthic hypodermis such as Everticyclammina and Pseudocyclammina are found in clear limestones, mostly free input of terrigenous particles. However, previously, Bassoullet et al. (1985) had already made an important observation about Pseudocyclammina, they noticed that this wall structure was a form adapted to detrital environments.
Later, Hughes (2004 b) indicated that the occurrence of Pseudocyclammina lituus and Alveosepta powersi is associated with deep-water foraminifera in the Hanifa Formation, which could indicate a slightly deep open marine environment. Hughes (2009) suggests open marine, moderately deep conditions based on the presence of Alveosepta jaccardi, Pseudocyclammina lituus, Kurnubia palastiniensis, and Nautiloculina oolithica in part of the Jubaila Formation, the two studies were conducted in Saudi Arabia.
The observations from other localities agree with the association identified in the locality under study. In addition, the thin-section petrographic analysis of the samples, that is based on the textural features allowing to define the microfacies as a bioclastic-wackestone with peloids, grains of quartz, that contains a larger benthonic foraminiferal assemblage, moreover bivalves, gastropods, echinoderms which could correspond to open platforms SMF 8 of Flügel (2010) or FZ 7 of Wilson (1975): open marine with shallow-water larger foraminifera and algae.
Paleobiogeography
The fragmentation of Pangea during the Late Jurassic- Early Cretaceous resulted in an intense tectonic activity, with palaeoceanographic changes including the continued expansion of the Central Atlantic and Western Tethys opening. Shallow seaways covered the continents, and marine and marginal marine sediments were deposited, preserving a diverse assemblage of fossils as the larger foraminifera (Boudhager-Fadel, 2008). The colonization of the American margins took place from the Callovian or Oxfordian (Fourcade and Michaud, 1987).
During the Kimmeridgian, climate warming permitted the Tethyan foraminifera to reach the most northerly paleolatitudes, which are a little further north than 35° N (Bassoullet et al., 1985).
The larger foraminifera identified in our material as Alveosepta jaccardi, Everticyclammina virguliana, Pseudocyclammina lituus, and Nautiloculina oolitica have been recorded in the platform of the Atlantic coast (Lisbon and Algarve, Ramalho 1971), in eastern Morocco (Hottinger 1967), in southern Morocco in the Tarfaya Province near the Cansado Port (Viotti, 1965), and northern Morocco in the Essaouira basin (Jaffrezo et al., 1985; Peybernès et al., 1987). The Mekele Basin in northern Ethiopa is the farthest south area where this association has been recorded (Adefris et al., 2021).
Alveosepta jaccardi has been identified for time in middle Oxfor- dian beds of the Theniet EtTemar Formation from the central Saharan Atlas in Algeria (Mahboubi et al., 2023). On the Senegal coast, in the oolitic limestone only two species, Alveosepta jaccardi and Everti- cyclammina virguliana, have been recorded (Castellain, 1965). This place is the most southern location that contains larger foraminifera in the Atlantic of Oxfordian-Kimmeridgian age, however toward the north, Alveosepta jaccardi and Kurnubia palastiniensis were recorded in Baltimore Canyon and the Nova Scotia shelf (Ascoli, 1976, Ascoli et al., 1984)), as well as the SW of the Grand Bank (Gradstein, 1978). Cladoropsis mirabilis (Felix) has been identified in our material.
It is found worldwide in the upper Oxfordian-Kimeridgian, especially on the southern edge of the Tethys in Spain, Italy, Yugoslavia, Greece, Algeria, Lebanon, and Japan (Termier et al., 1977), as well as in Saudi Arabia (Hughes, 2004 b) and Turkish (Tasli, 2001).
DISCUSSION
The breakup of Pangea during the Jurassic resulted in the gradual opening of the Tethys into a dominant marine seaway of the Mesozoic. A great amount of warm water flowed westward between the continents and connected the major oceans, extending around the earth between approximately 30° N and the equator, producing a significant effect on the Earth’s climate (Scotese et al., 1999; Tabor et al., 2019).
The earliest oceanic crust formed during the middle Jurassic, marks the opening of the Western (or Mediterranean) Tethys. This branch of the neo-Tethys Ocean persisted until the Hispanic corridor to the Caribbean area and it was closed by the northward movement of Africa; in the Late Jurassic, on the northern Tethys margin, were developed carbonate platforms from the Cuba-Bahamas-Florida area extending to Tibet (Forda and Golonka, 2003).
The development of the new foraminiferal biota in the Jurassic occurred against the background of significant events in the Earth’s history: the termination of the Early Cimmerian orogeny, and the opening of the North Atlantic (Boudhager-Fadel, 2008).
In Mexico, there are few reports about the presence of the Jurassic larger foraminifera, for instance, only has been reported from the San Ricardo Formation at Chiapas (Michaud, 1987; Ornelas and Hottinger, 2006), and in the marine wells drilled in the Campeche Sound (Gulf of Mexico) by Ornelas et al. (1993).
The Jurassic marine strata have not been reported at the Reyes Metzontla locality in the Zapotitlán basin. According to Centeno- García et al. (2009), a Mesozoic sedimentary succession composed of red beds had been recorded (García Calderon,1956), considered to be of Jurassic age due to the stratigraphic position and its lithological similarity to the rocks that abound in the nearby localities of Huajuapan de León and Santo Domingo Tianguistengo (Morán Zenteno et al., 1993). The marine rocks of the Zapotitlán and San Juan Raya Formations of Cretaceous age were deposited overlying the red beds. However, other authors had already studied the region; for instance, Pérez-Ibargüengoitia et al. (1965, p. 14) proposed a lithostratigraphic unit named the Mapache Formation, located at the Tocomazuchil stream (5.2 km. from the Petlalcingo, south Puebla) containing a variety of invertebrate fossils, which they dated as Late Jurassic (Kimmerigdian-Portlandian). This locality is situated SW of
the Los Reyes Metzontla.
Buitrón and Barceló Duarte (1980. p. 47) indicated that the Zapotitlán Formation rests discordantly on the Mapache Formation in the Tehuacán-San Juan Raya area.
Omaña and González Arreola (2008) recorded an algae and larger foraminiferal association at Santiago Coatepec of the Kimmeridgian. This locality is situated SW of the Los Reyes Metzontla, it contains a greater diversity of the fossils of this age. Thus, our research is a significant contribution to knowledge of the stratigraphy of the region, recording for the first time a Late Jurassic (Kimmeridgian) deposit in the Los Reyes Metzontla locality, mostly based on the occurrence and biostratigraphic significance of the larger foraminiferal assemblage composed of Pseudospirocyclina mauretanica, Chofatella tingitana, Pseudocyclammina lituus, Everticylamina virguliana, among others.
Additionally, this finding also enables a better understanding of the paleoenvironment, indicating a shallow-water deposit that was inhabited by the larger foraminifers as well as mollusks such as Trichites sp.
The known paleogeographic distribution of the larger foraminiferal assemblage present in our material indicates that they were cosmopolitan forms mostly associated with the Tethyan province during the Late Jurassic (Pelissie et al., 1982; Basoullet et al., 1985).
CONCLUSION
The proposed age of the studied sequence is principally based on the study of the larger foraminiferal assemblage. The paleoecological and palaeobiogeographical distribution is examined.
The stratigraphic distribution of the larger benthic foraminifera enabled us to suggest a Kimmeridgian age for the studied succession. The information furnished by these microfossils as well as the micro facies analysis, which consists of a wackestone with peloids and quartz grains with scarce presence of gastropods, bivalves and echinoderm plates, moreover of the larger benthic foraminifera. All components are poorly sorted and embedded within a dark-colored micritic matrix suggests an environment of warm shallow water with a low to medium energy regime. Larger benthic foraminifera identified are characteristics of the Tethyan Realm.

