











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Durante las últimas décadas, las poblaciones de la avifauna en América del Norte han experimentado un deceso significativo, y se estima que las aves de pastizal han sufrido una disminución de más del 50% de sus poblaciones (Rosenberg et al., 2019). Una característica notable es que alrededor del 80% de las aves de pastizal son migratorias; se reproducen en el norte del continente y migran al sur durante el invierno en busca de recursos para su supervivencia (Pool et al., 2014).
En este contexto, el gorrión de Baird (BAIS, Centronyx bairdii) es una especie de ave focal de pastizal que pasa el inverno en el Desierto Chihuahuense, presentando una preocupante tendencia en la disminución poblacional a una tasa anual de -2.93% (Sauer et al., 2020), por lo que ampliar el conocimiento del gorrión de Baird durante su estadía invernal puede favorecer al impulso de estrategias de conservación apropiadas y de largo plazo (Berlanga et al., 2010).
Esta disminución se atribuye principalmente a la pérdida y fragmentación del hábitat, provocadas por actividades antropogénicas como la agricultura, el sobrepastoreo, la conversión de la tierra y la urbanización (Pool et al., 2014). El estudio de esta especie es crucial porque se podría considerar como un indicador biológico de la salud del pastizal (Elliot & Johnson, 2018); así mismo, el gorrión de Baird depende de la condición de la estructura y la productividad de los pastizales naturales para su protección y alimentación. Por tanto, los cambios que provocan la fragmentación y pérdida de hábitat lo convierten en una especie vulnerable de ese ecosistema (Macías & Panjabi, 2013).
Se han llevado a cabo diversos estudios con el fin de comprender los procesos ecológicos que tienen lugar durante el periodo invernal del gorrión de Baird en el Desierto Chihuahuense. Estos estudios se han enfocado en investigar su distribución y abundancia (Martínez-Guerrero et al., 2011a), así como en analizar su supervivencia y los factores de depredación (Strasser & Panjabi, 2016). La estimación del tamaño del ámbito hogareño en las aves es fundamental para comprender el área que cada individuo requiere para obtener los recursos necesarios para su supervivencia (Burt, 1943). Durante la temporada invernal, estos recursos incluyen alimento, áreas de descanso y refugio contra los depredadores (Macías-Duarte et al., 2018).
En este contexto, las semillas de diferentes pastos y hierbas disponibles en el suelo, conocido como banco de semillas, representan la principal fuente de alimentación tanto para el gorrión de Baird como para otras aves de pastizal (Mendez-Gonzalez et al., 2010). La altura del pasto y la cobertura de hierba son atributos vegetales cruciales en la selección del hábitat de esta especie durante el invierno (Martínez-Guerrero et al., 2011b). El comprender estos factores es esencial para garantizar la supervivencia del gorrión de Baird y la salud de los pastizales, lo que, a su vez, contribuye al equilibrio ecológico y a la preservación de la biodiversidad en general.
Tomando en cuenta lo expuesto anteriormente, el objetivo principal de esta investigación fue analizar la relación entre el ámbito hogareño invernal, la densidad poblacional y la disponibilidad del banco de semillas en el suelo. Además, se buscó describir los atributos específicos de la vegetación en los sitios utilizados por el gorrión de Baird en los pastizales nativos del norte de Durango. Comprender las variables ecológicas del gorrión de Baird durante el invierno es fundamental para su conservación, ya que permite identificar y proteger hábitats clave, asegurar la disponibilidad de recursos esenciales, y anticipar y mitigar los efectos del cambio climático y otras amenazas. Todo esto contribuye a la supervivencia de la especie a largo plazo.
Materiales y métodos
Área de estudio
El estudio se llevó a cabo en tres sitios de pastizal natural ubicados al norte del municipio de Hidalgo, Durango, México (Figura 1). Estos sitios fueron identificados con acrónimos para facilitar su registro y manejo de información: "Doña Wencha" (DOWE), "La lagunita" (LALA) y "La potranca" (LAPO). Todos ellos se encuentran dentro del Área Prioritaria para la Conservación de Pastizales (GPCA, por sus siglas en inglés) (Commission for Environmental Cooperation & The Nature Conservancy [CEC & TNC], 2005; Pool & Panjabi, 2011) en Cuchillas de la Zarca (CUZA).
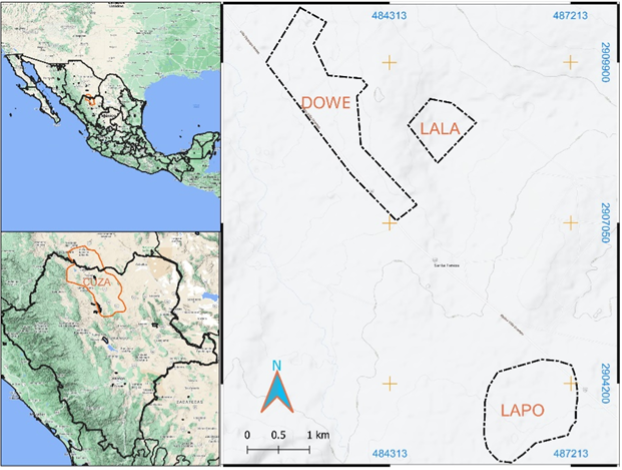
Fuente: Elaboración propia con imágenes de Google Earth en plataforma de QGIS.
Figura 1 Localización de los sitios de estudio DOWE, LALA y LAPO dentro del GPCA-CUZA.
Los sitios DOWE, LALA y LAPO (Tabla 1) se caracterizan por su estrato herbáceo, que incluye especies de gramíneas, herbáceas y arbustivas de géneros como Aristida, Bouteloua, Mulhenbergia, Panicum y Bothriochloa. Entre las hierbas dominantes se encuentran Aldama linearis y Dyssodia papposa, mientras que la comunidad arbustiva está compuesta mayormente por especies de los géneros Juniperus, Acacia y Prosopsis (Martínez-Guerrero et al., 2011a).
Tabla 1 Sitios de estudio utilizados por el gorrión de Baird en la región de Cuchillas de la Zarca.
| Nombre del sitio | Acrónimo | Altitud (m. s. n. m.) | Coordenadas | Superficie (ha) |
| Doña Wencha | DOWE | 1873 | 26° 17.7’ N; 105° 9.7’ O | 240 |
| La Laguna | LALA | 1866 | 26° 17.8’ N; 105° 8.8’ O | 45 |
| La Potranca | LAPO | 1920 | 26° 14.8’ N; 105° 7.6’ O | 109 |
Fuente: Elaboración propia.
Captura y procesamiento de aves
Para la captura y manipulación del gorrión de Baird, durante el invierno 2018-2019 se obtuvo el permiso de colecta SEMARNAT SPGA/DGVS/001783/18. Se utilizaron cuatro redes de niebla para la captura de aves, las cuales presentan dimensiones de 12 m de largo, 2.60 m de alto y una malla de 36 mm, suministrado por Avian Research Supplies (ARS). La captura se llevó a cabo durante tres días consecutivos (de 8 am a 12 pm y de 4 pm a 7 pm) con la colaboración de un equipo de ocho a diez personas, quienes implementaron el método de arreo de las aves hacia las redes (Beyer & Panjabi, 2010).
Tras la captura, a cada ejemplar se le colocó un anillo metálico talla número 1 suministrado por el United States Geological Survey (USGS) para su identificación, y se registraron sus medidas morfométricas. Posteriormente, se les colocó un transmisor modelo PicoPip 379 de LOTEK®, con un peso de 0.5 g y una duración de batería de aproximadamente 40 días. Este transmisor se sujetó al dorso del ave mediante un arnés elástico (Rappole & Tipton, 1991) con el propósito de realizar un seguimiento mediante la técnica de telemetría. Después de un periodo de 30 días, las aves fueron recapturadas para cambiar el transmisor.
Se llevó a cabo el seguimiento diario de las aves equipadas con transmisor (n = 50) utilizando un receptor de radio ATS de la marca Biotrackers®, junto con una antena Yagi de tres elementos portátil y auriculares (Modelo David Clark® modelo H10-00-4). La ubicación de las aves fue registrada con precisión utilizando un navegador GPS marca Garmin® modelo Vista.
El tamaño del ámbito hogareño invernal (AHI) de cada ave (n = 26) se determinó utilizando el método de Kernel fijo (K) al 95% de confianza, con validación cruzada de mínimos cuadrados (Powell, 2000; Seaman & Powell, 1996; Seaman et al., 1999). Este método requiere un mínimo de 30 detecciones por individuo. El análisis se realizó empleando el paquete "the adehabitat HR" del software R, versión 1.1.463 (R Development Core Team, 2018). Se eliminaron valores atípicos o movimientos exploratorios de las aves que podrían resultar en una sobreestimación del ámbito hogareño, y se procedió a eliminar manualmente las ubicaciones más alejadas del centro de gravedad de la nube (Börger et al., 2006; Getz et al., 2007)
Una vez calculados los AH individuales por sitio, se generó una lista de números aleatorios correspondientes a los registros de ubicaciones diarias de cada ave (punto central georreferenciado de las aves). Se seleccionaron las primeras 10 detecciones de esta lista para obtener el banco de semillas en el suelo y estimar la cobertura porcentual de pasto, hierba, arbustos, suelo desnudo y otras formas de cobertura, como vegetación muerta y excremento animal. Además, se tomaron medidas en centímetros de la altura del pasto, hierba y arbustos (Strasser & Panjabi, 2016).
Biomasa de semilla
Para la obtención del banco de semillas en el suelo (n = 210), se insertó un aro metálico de 8.8 cm de diámetro a una profundidad aproximada de 0.5 cm, recolectando así un volumen de 30 cm3 de suelo (Ginter & Desmond, 2005; Pulliam, 1980, 1986). Posteriormente, las muestras fueron llevadas al laboratorio de Posgrado de la Facultad de Medicina Veterinaria y Zootecnia de la Universidad Juárez del Estado de Durango, donde se tamizaron para eliminar residuos de mayor tamaño, como rocas, materia orgánica, tierra y basura.
Las semillas se separaron manualmente con la ayuda de un estereoscopio 10 × 40 de la marca Leica® modelo EZ4, y se pesaron en una balanza analítica (modelo CP224 S de Sartorius), con el fin de obtener la biomasa promedio del banco de semillas (g/m2), tal como se describe en el trabajo de Desmond et al. (2008).
Densidad de aves
Para estimar la densidad de gorriones de Baird se realizaron 17 transectos de 500 m de longitud (Strasser et al., 2018). Apoyados de un GPS (GARMIN® etrex 20) y los puntos previamente cargados, dos personas recorrieron a pie los transectos para cubrir un rango de visión más amplio (Panjabi et al., 2010a). Para el conteo e identificación de las aves, se usaron binoculares Bushnell® prime 15 × 50, y para medir la longitud entre las detecciones de aves y la línea del transecto se utilizó un distanciómetro láser (Yardage Pro scout).
Caracterización de la cobertura y estructura vegetal
Para la estimación de la cobertura y estructura vegetal (n = 210), se utilizó la metodología propuesta por Macías et al. (2018), que consiste en estimar visualmente la cobertura vegetal (%) de pasto, hierba, arbustos, suelo desnudo y otra cobertura (material leñoso, excremento animal, rocas mayores a un puño) en un radio de 5 m. Adicionalmente, se registró la especie de pasto dominante por cada punto de muestreo. Para la estimación de las alturas de pasto, hierba y arbustos, se utilizó un tubo Robel graduado en centímetros (Robel et al., 1970).
Análisis estadístico
Para las variables de AH, biomasa de semillas en el suelo (g/m2), cobertura de la vegetación (%) y altura de la vegetación (cm), se aplicó un análisis de varianza de una vía utilizando estadística no paramétrica (Kruskal-Walis, p ≤ 0.05). Con el programa DISTANCE® 7.3 (Thomas et al., 2010) se estimó la densidad de aves por sitio empleando el muestreo de distancias convencional (CDS) y el ajuste medio normal +coseno para modelar la función de detección. Para determinar el grado de relación entre AHI y las variables antes descritas, se realizó un análisis de correlación de Spearman (r s ). El análisis de los datos se realizó mediante el software R, versión 1.1.463 (R Development Core Team, 2018).).
Resultados y discusión
Ámbito hogareño invernal
Durante el invierno 2018-2019, se capturaron un total de 46 aves de la especie Centronyx bairdii, distribuidas de la siguiente manera: DOWE = 11, LALA = 7 y LAPO = 28. Sin embargo, sólo 26 aves proporcionaron las detecciones necesarias (>30) para la construcción del ámbito hogareño, generando así un total de 1752 puntos de detección. El tamaño promedio del ámbito hogareño invernal fue de 0.042 ha (IC 95% ± 0.006 ha) (Tabla 2). Este valor resulta menor en comparación con los hallazgos de Strasser et al. (2018), quienes estimaron un tamaño territorial de 1.57 ha para la misma especie en distintas regiones del Desierto Chihuahuense. De igual manera, Sierra-Franco et al. (2019) y Herrera-Vargas et al. (2021) reportaron 4.82 ha y 0.105 ha, respectivamente, para la misma región. Estos patrones de movimiento del gorrión de Baird son cambiantes e irregulares de un periodo a otro, debido a la variabilidad espacial y temporal de la distribución de las lluvias y semillas (Rechetelo et al., 2016).
Tabla 2 Ámbito hogareño invernal, biomasa de semillas en el suelo, densidad poblacional y cobertura y estructura vegetal (promedio ± E.E.) en tres sitios utilizados por el gorrión de Baird en la región de Cuchillas de la Zarca.
| Variable | Sitio | ||
| DOWE (N = 5) | LALA (N = 5) | LAPO (N = 16) | |
| AHI (ha) | 0.07 ± 0.07b | 0.43 ± 0.07a | 0.04 ± 0.04b |
| Semilla (g/m²) | 2.04 ± 1.45c | 12.45 ± 1.45a | 4.64 ± 0.81b |
| Densidad (ind/ha) | 0.95 ± 0.39b | 1.63 ± 0.31b | 2.65 ± 0.38a |
| Cobertura (%) | |||
| Hierba | 2.34 ± 0.45b | 34.4 ± 3.45a | 5.38 ± 1.07b |
| Arbusto | 7.06 ± 0.61a | 0.00 ± 0.61c | 2.00 ± 0.19b |
| Pasto | 52.6 ± 4.43b | 41.7 ± 4.43b | 72.92 ± 1.38a |
| Suelo desnudo | 34.3 ± 3.07a | 19.06 ± 3.07ab | 12.68 ± 0.96b |
| Otra cobertura | 3.7 ± 1.43b | 4.84 ± 1.43ab | 7.02 ± 0.44a |
| Altura (cm) | |||
| Hierba | 11.78 ± 4.14b | 31.58 ± 4.1 4a | 29.91 ± 1.29a |
| Arbusto | 68.9 ± 7.48a | 0.00 ± 7.48c | 47.51 ± 2.33b |
| Pasto | 31.54 ± 2.21ab | 16.06 ± 2.21b | 36.36 ± 0.69a |
Nota. Valores con diferente literal indican diferencia significativa p ≤ 0.05 K-W.
Fuente: Elaboración propia.
La variación del tamaño del AHI puede reflejar la respuesta de los individuos a variables ambientales como: cambios en la precipitación, que afecta directamente a la vegetación de los pastizales (Bengsen et al., 2016); la temperatura baja, que puede restringir movimientos y comportamiento en la búsqueda de alimento (Haase et al., 2019); el fotoperiodo durante el proceso de migración, que modifica sus hábitos alimentarios duplicando sus hábitos alimentarios (Emlen, 1975), entre otros factores (Börger et al., 2006). Estos cambios pueden manifestarse de manera gradual o abrupta en las condiciones del hábitat para las aves, lo cual puede influir en la dimensión del territorio ocupado, especialmente en especies locales o migratorias, donde dicha dimensión puede depender de la escala de tiempo (Börger et al., 2008) y las modificaciones que experimente el hábitat a lo largo del tiempo. Conocer el ámbito hogareño invernal de Centronyx bairdii es crucial, ya que permite identificar las áreas donde se encuentran y cómo utilizan estos espacios durante los meses de invierno. Esta información es vital para proteger las zonas de invernada, que son esenciales para su supervivencia y para asegurar que las aves dispongan de los recursos necesarios para mantenerse en buena condición de cara a la siguiente temporada de reproducción (Martínez-Guerrero et al., 2022).
El cambio climático puede alterar significativamente las áreas invernales de muchas especies, modificando la disponibilidad de recursos y las condiciones ambientales (Salazar-Borunda et al., 2022; The Nature Conservancy [TNC], 2020). Comprender el ámbito hogareño invernal de Centronyx bairdii permite anticipar cómo podrían verse afectadas y qué medidas de conservación serían necesarias para mitigar estos impactos.
Durante el invierno, las aves enfrentan nuevos desafíos, como la competencia por recursos limitados y las interacciones con otras especies. Estudiar su ámbito hogareño invernal ayuda a entender estas interacciones y su impacto en la ecología de la especie. La conservación efectiva de especies migratorias requiere un enfoque integral que abarque todo su ciclo de vida, incluyendo tanto las áreas de reproducción como las de invernada. Conocer el ámbito hogareño invernal de Centronyx bairdii es fundamental para diseñar estrategias de conservación completas y eficaces.
Banco de semilla en el suelo, densidad poblacional del gorrión de Baird y cobertura y estructura de la vegetación
Se recogieron y procesaron 10 muestras de semillas para cada uno de los AHI estimados (n = 26), lo que sumó un total de 260 muestras. Se determinó un promedio de 5.65 g/m2 de biomasa de semillas en el suelo que contrasta con los hallazgos de Cabanillas et al. (2016), quienes reportaron 0.89 g/m2, y los de Salazar-Sánchez et al. (2021), quienes encontraron 1.69 g/m2 de biomasa de semillas depositadas en el suelo para el mismo sitio de estudio. La cantidad y distribución de la lluvia durante la temporada de crecimiento de las plantas en el Desierto Chihuahuense es un factor que puede explicar las discrepancias entre los diferentes años para esta variable (Reichmann & Sala, 2014).
El gorrión de Baird, siendo una especie focal de pastizal, no posee la misma capacidad de adaptación en la selección de la dieta que otras aves granívoras que habitan en ambientes desérticos (Marone et al., 2017). Los estudios relacionados con la disponibilidad de semillas en el suelo y la dieta invernal de aves focales de pastizal son escasos (Mendez-Gonzalez, 2010; Titulaer 2016; Titulaer et al., 2017). Sin embargo, recientemente se han realizado trabajos para la región de Cuchillas de la Zarca con el gorrión chapulín (Ammodramus savannarum) y el gorrión de Baird (Centronyx bairdii) en la selección de hábitat, potencial energético de la semilla y relación del ámbito hogareño con las semillas (Herrera-Vargas et al., 2021; Martínez-Guerrero et al., 2022; Salazar-Sánchez et al., 2021).
Durante el presente estudio, se observó una variación significativa en la disponibilidad de semillas en el suelo en los sitios donde se detectó el gorrión de Baird (p ≤ 0.05) (Tabla 2). Se destacó especialmente el sitio LALA, que presentó una cobertura importante de hierba del 34.4% y una disponibilidad de semilla de 12.45 g/m2. Esta variable podría desempeñar un papel crucial como fuente de alimento durante la temporada no reproductiva para las aves invernales de pastizal (Brown & Long, 2007; Brown & Sherry, 2006). La cantidad de semillas encontradas en el sitio LAPO (4.64 g/m2) fue similar a la reportada por Mendez-Gonzalez (2010) en un área de pastizal en Estados Unidos, donde los valores oscilan entre 4.0 g/m2 y 14.0 g/m2. Sin embargo, estos valores fueron mayores que los estimados en este estudio para el sitio DOWE (2.04 g/m2).
Las comparaciones realizadas entre los sitios (DOWE, LALA y LAPO) revelaron diferencias estadísticas (p ≤ 0.05) (Tabla 2) en las variables de ámbito hogareño, densidad de semillas y estructura de la vegetación. Sin embargo, se detectó la presencia del gorrión de Baird en los tres sitios, lo que sugiere que éstos poseen las características requeridas para la estancia invernal de la especie (Martínez-Guerrero et al., 2011b; Panjabi et al., 2010b).
La densidad de aves entre los sitios varió significativamente (p ≤ 0.05), siendo el sitio LAPO el que presentó el valor más alto (2.65 ind/ha), en comparación con los otros dos sitios (DOWE y LALA). Es importante destacar que la densidad de aves en los tres sitios difiere de las estimaciones reportadas por Cabanillas et al. (2016) y Martínez-Guerrero et al. (2011a) para la misma región, donde se registraron 0.05 ind/ha y 0.08 ind/ha, respectivamente.
El sitio LAPO se caracterizó por tener una mayor cobertura de pasto (72.92%) y una menor área de suelo desnudo (12.68%). Estos hallazgos coinciden con investigaciones previas (Elliot & Johnson, 2018) que sugieren que ciertas especies de aves de pastizal pueden actuar como indicadores biológicos de la salud del pastizal. Se ha documentado que estas especies prefieren hábitats con una mayor cobertura de pasto y altura de hierba (Macías-Duarte et al., 2018; Sierra-Franco et al., 2019), los cuales proporcionan recursos alimenticios, protección y perchas durante el invierno (Martínez-Guerrero et al., 2011a).
Dentro de los territorios utilizados por los gorriones de Baird, se registraron ocho especies de pastos característicos del Desierto Chihuahuense (Brown & Makings, 2014) (Tabla 3). Destacan los pastos Navajita (Bouteloua gracilis) y espiga negra (Hilaria cenchroides) como las especies más abundantes, mientras que Bothriochloa barbinodis y Muhlenbergia spp. fueron los pastos más altos encontrados en estos territorios. Esta información es crucial puesto que las aves de pastizal prefieren semillas de ciertas especies de gramíneas nativas en sus sitios de invernada (Titulaer et al., 2017). El sitio LAPO se distinguió por albergar la mayoría de las especies de gramíneas registradas en los tres sitios, lo cual coincide con lo reportado por Arroyo-Arroyo et al. (2020) para la misma zona, quienes identificaron seis de las ocho especies de gramíneas presentes en esta investigación.
Tabla 3 Porcentaje de los principales pastos en los sitios utilizados por el gorrión de Baird en Cuchillas de la Zarca.
| Especies | Cobertura (%) | ||
| DOWE | LALA | LAPO | |
| Aristida spp. | 29.86 | 1.45 | 22.38 |
| Bothriochloa barbinodis | 0.68 | 0 | 1.17 |
| Bouteloua curtipendula | 18.07 | 0 | 14.92 |
| Bouteloua gracilis | 29.17 | 34.06 | 44.06 |
| Hilaria cenchroides | 22.22 | 42.03 | 5.13 |
| Muhlenbergia spp. | 0 | 17.39 | 11.87 |
| Sporobolus airoides | 0 | 5.07 | 0 |
| Pleurafis mutica | 0 | 0 | 0.47 |
Fuente: Elaboración propia.
La mayor densidad de aves en LAPO puede deberse a diversos factores ecológicos y ambientales. Entre ellos, la disponibilidad de semillas juega un papel crucial, ya que las áreas con alta producción de semillas, tanto de pastos nativos como de plantas anuales, tienden a atraer y sostener mayores densidades de aves (Martínez et al., 2020; Rechetelo et al., 2016). Además, los pastizales con una estructura vegetal adecuada, que incluya una mezcla de áreas abiertas y vegetación baja, proporcionan refugio y acceso al alimento. La presencia de pequeños parches de vegetación densa también es importante, ya que ofrece protección contra depredadores (Martínez et al., 2020).
Aunque las aves granívoras de pastizal no requieren grandes cuerpos de agua, la disponibilidad de fuentes de agua accesibles, como pequeños estanques o charcas, puede incrementar la densidad de aves, ya que es esencial para su hidratación (Bock, 2015). Las relaciones entre especies, como la competencia por recursos y la presencia de especies dominantes, también pueden influir en la densidad de aves granívoras. En algunos casos, las aves se benefician de la agregación, lo que puede aumentar la eficiencia en la localización de alimentos y reducir el riesgo de depredación (Huang et al., 2021). Estos factores interactúan de manera compleja, y su combinación determina la capacidad de un hábitat para sostener altas densidades de aves granívoras durante el invierno.
Relación entre ámbito hogareño, biomasa de semilla y características de la vegetación invernal (cobertura, estructura)
En los sitios DOWE y LALA se observaron correlaciones negativas significativas (Spearman p ≤ 0.05) entre el ámbito hogareño invernal y la biomasa de semilla en el suelo, lo cual concuerda con los informes de Mendez-Gonzalez (2010) y Cabanillas et al. (2016), quienes estimaron valores de 4.0 g/m² - 14.0 g/m² y 1.65 g/m², respectivamente. Ambos autores sugieren que el tamaño del territorio puede ser menor cuando hay una alta biomasa de semillas disponibles en el suelo. Sin embargo, algunos autores como Bechtoldt & Stouffer (2005) mencionan que los gorriones de Baird son especialistas del ecosistema pastizal, por lo que su densidad, ámbito hogareño y distribución espacial pueden correlacionarse positivamente con la biomasa de semillas depositadas en el suelo.
De igual manera, estudios realizados con aves granívoras de pastizal (Bechtoldt & Stouffer, 2005; Grzybowski, 1983; Macías-Duarte et al., 2018) han encontrado una relación positiva entre la densidad y la abundancia invernal con la disponibilidad de semillas en el suelo. Además, Pulliam & Dunning (1987) demostraron la asociación entre la abundancia de aves de pastizal y la disponibilidad de semillas durante el invierno, considerando a la vegetación como un elemento importante por su capacidad para producir alimento.
En el sitio LALA, se observó que el AHI y la cobertura de hierba fueron mayores en comparación con los otros dos sitios. En el sitio LALA, se observó que el AHI (0.43 ha) y la cobertura de hierba (34.4%) fueron mayores en comparación con los otros dos sitios. Esto podría indicar que el banco de semillas en el suelo tendría una mayor proporción de semillas de hierbas (variable no medida en este estudio). A partir de lo anterior, podemos asumir que éstas también constituyen una fuente importante de alimento para el gorrión de Baird, lo que les permite llevar a cabo sus actividades cotidianas como volar, buscar alimento y escapar de sus depredadores (Titulaer et al., 2017).
En este estudio, se encontró que las variables de la cobertura y estructura de la vegetación no mostraron relaciones significativas (Spearman p ≥ 0.05) con respecto al AHI. Sin embargo, investigaciones previas han revelado relaciones positivas de los gorriones de Baird a variables como la cobertura y altura de pasto, así como relaciones negativas con el suelo desnudo y otras formas de cobertura (De León-Mata et al., 2020; Martínez-Guerrero et al., 2011b; Pool et al., 2014; Rodríguez-Maturino et al., 2017).
Las principales amenazas son la pérdida de hábitat (debido a la conversión de pastizales naturales en tierras agrícolas o pastizales para alimentar el ganado, así como a la fragmentación del hábitat por la división de grandes extensiones de pastizales en pequeñas parcelas desconectadas, lo cual afecta la capacidad de las aves para encontrar refugio y alimento). El uso de plaguicidas y agroquímicos en la agricultura puede afectar directamente a las aves al intoxicarlas o, indirectamente, al reducir la disponibilidad de insectos (Rosenberg et al., 2019).
Algunas estrategias para la protección de los pastizales y las aves que en ellos habitan pueden ser la conservación y restauración de hábitat, el manejo sostenible de la tierra, el monitoreo e investigación, la educación y sensibilización y la aplicación de políticas y regulaciones para que los propietarios de las tierras adopten prácticas de manejo amigables.
Conclusiones
Conocer el ámbito hogareño invernal de Centronyx bairdii es esencial para identificar y proteger las áreas clave que son fundamentales para la supervivencia de la especie durante el invierno. La comprensión de estos territorios permite prever los efectos del cambio climático sobre la especie y tomar medidas de conservación adecuadas para mitigar los impactos negativos. Estudiar el ámbito hogareño invernal ayuda a comprender las interacciones con otras especies y cómo éstas afectan la ecología general de Centronyx bairdii. La información sobre el ámbito hogareño invernal es crucial para diseñar estrategias de conservación integrales, asegurando la protección de la especie a lo largo de todo su ciclo de vida. La protección y gestión adecuadas de los hábitats invernales son fundamentales para la conservación a largo plazo de Centronyx bairdii, contribuyendo a la estabilidad y viabilidad de sus poblaciones.
Se observó que el sitio LAPO presentó la mayor densidad de aves, lo cual sugiere que las características de este lugar proporcionaron condiciones favorables para la estancia invernal del Centronyx bairdii. Además, se encontró que los tres sitios de pastizal considerados en este estudio presentaron diferencias significativas en cuanto a la cobertura y estructura de la vegetación. Para futuras investigaciones, es crucial considerar variables adicionales no abordadas en este estudio, como la precipitación pluvial y la temperatura, con el fin de comprender mejor la interrelación entre las variables ambientales y su influencia sobre la selección del hábitat del gorrión de Baird durante su estancia invernal en México.
Conflictos de interés
Los autores no tienen conflicto de interés con la investigación realizada.

