











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCTION
During the last decades, Uruguay had a significant expansion of agricultural and forestry activity (Arriaga 2021, Boscana 2021) with a consequent increase in the importation and application of agrochemicals with high potential for environmental contamination (Lajmanovich et al. 2012, Daley et al. 2014, MGAP 2021). Studies carried out in different river basins in this country have recorded negative effects of insecticides on some non-target animal groups, such as fish and bees (Leites Cartagena 2009, Ríos et al. 2010, Colombo et al. 2011), as well as the presence of agrochemicals both in water and in sediments (Eguren et al. 2008, Nardo et al. 2015) that deteriorate water quality in limnic systems (Kruk et al. 2013).
The use of biological indicators not only helps to assess the quality of the environment, but also the responses of such organisms to environmental changes caused by pollution (Barbour et al. 1999, Hanlon and Relyea 2013, Pollo et al. 2019). In this sense, and due to their high sensitivity and biological characteristics, amphibians have been established as a good environmental bioindicators (Young et al. 2004, Hopkins 2007). Several amphibian species show population declines (IUCN 2022), as well as an increase in the frequency of malformations recorded in nature (Peltzer et al. 2011, Mónico et al. 2019); being exposure to chemical contaminants one of the main causes of these phenomena (Blaustein and Kiesecker 2002, Egea-Serrano et al. 2012). All these reasons could be the main cause that explain the increase in ecotoxicological studies with this zoological group, which is reflected in the literature (Curi et al. 2019, Pérez-Iglesias et al. 2019, Barreto et al. 2020, de Arcaute et al. 2020, Lajmanovich et al. 2022, Pereira et al. 2023).
Genetic biomarkers such as micronucleus (MN) frequency (Schmid 1975, Al-Sabti and Metcalfe 1995) and the formation of other nuclear anomalies or aberrations (AN) (Carrasco et al. 1990) are tools used for the evaluation of xenobiotic activity of contaminants. Among others, agrochemicals are of particular interest for ecologists and conservationists since their genotoxic activity reflects on DNA damage in the initial stages of exposure (William 2007, Fonseca et al. 2011, Eskandari et al. 2012). This damage is easily identified by microscope observation and can be quantified by the MN test (Bolognesi and Hayashi 2011). In this context, there is a growing need to carry out new research that provides knowledge about the risk of agrochemicals on amphibian populations. The objective of this study was to evaluate the lethality and genotoxic effects of Picloram (Tordon®24K) on Boana pulchella tadpoles.
MATERIALS AND METHODS
Chemical compound
Picloram (PIC) 4-amino-3,5,6-trichloropicolinic acid CAS No. 1918-02-01 (PAN 2019) is a rapidly absorbed, selective and systemic herbicide that acts as a growth regulator of woody plants, broadleaf weeds, both annuals and perennials (Corteva 2019). It is produced in many commercial formulations, and one of the most used in forest crops in Uruguay is Tordon® 24K (TOR) soluble concentrate (Dow AgriSciences, Argentina), with a PIC concentration of 278 g/L (240 g/L Eq.Ac.).
Test species
Boana pulchella (Duméril and Bibron 1841) is a Hylid frog that shows a wide distribution in the Pampa Biome, in both natural and urbanized environments (Agostini et al. 2009, Maneyro and Carreira 2012). It is considered a species of “least concern” both at global and at national scale (Kwet et al. 2004, Carreira and Maneyro 2015). Previous studies suggest that B. pulchella tadpoles are a suitable in vivo model to detect toxic effects induced by xenobiotics (Natale et al. 2018, Sansiñena et al. 2018, Barreto et al. 2020).
Test organisms
We collected three clutches of B. pulchella in seasonal ponds located in the western Montevideo department, Uruguay (34º47'21''S 56º18'58''W) where the use of agrochemicals is not established yet. The tadpoles were acclimatized in the laboratory and when they reached the GS25 and GS36 stages of development according to Gosner (1960), the acute and chronic toxicity experiments were started respectively. The collection, experimentation, and euthanasia methods were approved by experimental protocols number 474 and 684 according to the Ethics Commission for the use of Animals (CEUA) of the Facultad de Ciencias of the Universidad de la República.
Experimental design
Acute toxicity bioassay: LC50 determination
We performed experimental tests to determine the acute toxicity range of PIC. From these results, the nominal PIC concentrations used were 8.5 (T5), 10.5 (T4), 12.5 (T3), 14.5 (T2), and 16.5 (T1) mg/L, and we used negative controls consisted in filtered tap water dechlorinated by aeration for 48 hours. We prepared dilutions from a stock solution of 240 mg TOR/L. All test solutions and controls were carried out in duplicate, and we prepared them immediately before the experiment. We place eight tadpoles (N = 96) in glass containers (13 cm in diameter and 20 cm in height) with 1 L of solution. A static model without renewal standardized according to USEPA (USEPA 1975) was applied, with a photoperiod of 16:8 h light/dark and under a controlled temperature of 20±2 ºC. Tadpoles were exposed to different PIC solutions for 48 hours to estimate the mean lethal concentration (LC5048h), lowest observed effect concentration (LOEC), and no observed effect concentration (NOEC). According to USEPA (1975) the following physical and chemical parameters were controlled: dissolved oxygen (DO) 8.2±1.5 mg/L, conductivity 247.0±12.5 mS/cm3, and pH 7.5±0.05. We randomly choose the location of the containers and the individuals used in each treatment. The tadpoles were not fed, and mortality was assessed by visual observation every 24 hours. Organisms were considered dead when no movement was detected after gently moving them with a glass rod. Cleaning tasks and removal of dead organisms were carried out every 24 hours.
Chronic toxicity bioassay
We used a static model with standardized renewal according to estimate sublethal effects of genotoxicity (USEPA 1975) under the same conditions of temperature, photoperiod, and physical and chemical parameters as those used in the acute bioassay. Tadpoles at the GS36 (Pérez Iglesias 2018) were exposed to nominal PIC concentrations corresponding to NOEC (12.5 mg/L) and NOEC/1.5 (8.5 mg/L). For negative controls we used filtered tap water dechlorinated by aeration for 48 hours. Four tadpoles (N=36) were placed per treatment, where all test solutions and controls were carried out in triplicate. All solution media were prepared immediately before use and completely replaced every 48 hours. We fed tadpoles throughout the experiment using boiled lettuce (Lactuca sativa) and fish flake food (Spirulina). The endpoint evaluation was made at eight weeks of exposure, when at least one larva reached stage 45 (GS45), that is, the stage before the metamorphic climax (Gosner 1960). Once this stage was reached, the micronucleus test was applied to all the tadpoles.
Micronucleus (MN) test
We select tadpoles from the chronic toxicity test for the evaluation of genotoxicity at sublethal concentrations (recorded mortality = 0.72% - two tadpoles). Peripheral blood smears were made (in duplicate) for each individual (N = 34) on clean, air-dried slides fixed with absolute ethyl alcohol for 20 min, then stained with acridine orange (Sigma-Aldrich, CAS No. 10127-02-3) for 10 min. The slides were coded and scored blindly by an observer in a Zeiss Axioplan 2 epi-fluorescence microscope with 400X magnification according to the criteria of Krauter (1993), Lajmanovich et al. (2005) and Cabagna et al. (2006). MN were identified as chromatin fragments smaller than one-third of the main nucleus, showing the same staining intensity and with a border distinguishable from the main nucleus border (Schmid 1975, Fenech 2000, Zalacain et al. 2005). The frequency of MN was expressed as the total number of MN per-1000 peripheral mature erythrocyte (Candioti et al. 2010).
Following the same procedure, other nuclear abnormalities (AN) were evaluated according to the criteria established by Cavaş and Ergene-Gözükara (2003) and Carrasco et al. (1990). Briefly, cells with two nuclei were considered binucleate. Cells with a nucleus showing a relatively small invagination of the nuclear membrane were classified as kidney shaped. Nuclei with large evagination from the nuclear membrane, which could have several lobes, were considered lobed nuclei. Nuclei with vacuoles and appreciable depth into a nucleus without containing nuclear material were recorded as notched nuclei. Finally, fragmented nuclei partially joined by chromatin were classified as bridged nuclei. The value of the NA was the outcome of the sum of all types of observed aberrations (Lajmanovich et al. 2014).
Data processing and analysis
The determination of the LC50 concentration was done using the non-parametric Spearman-Karber test (α = 0.05). For genotoxicity evaluation, data for MN and AN frequencies were applied to the binomial proportion test (Margolin et al. 1983). Statistical analyses were performed with the BioEstat.5.0 software - (Ayres et al. 2007). A p-value < 0.05 was considered to correspond with statistical significance for all tests unless indicated otherwise.
RESULTS
Survival as a function of exposure time observed in the acute toxicity test is shown in table I. The LC50 at 48h value was 16.5 mg/L (confidence limits: 15.68 - 17.32, Spearman-Karber test). The NOEC value was 12.5 mg/L, and the LOEC value was 14.5 mg/L for PIC. Mature erythrocytes from B. pulchella tadpoles were oval with a single well-defined and visible central nucleus, allowing easy recognition of the cytoplasm (Fig. 1 A). The MN were observed and identified as spherical nuclear fragments detached from the main nucleus (Fig. 1 B, C, and D). Some of the erythrocytes presented NA of the kidney-shaped nucleus, lobulated, with bridges and binucleated cells (Fig. 1 B, C, D, E, F). The data obtained on the frequency of MN in erythrocytes of the tadpoles showed an increase in the concentrations of 8.5 and 12.5 mg/L with statistically significant differences with the control group (Fig. 2). AN were observed in all test treatments, mainly kidney-shaped nuclei and binucleated cells, revealing a significant increase in AN frequency compared to the negative control (Binomial, 8.5 mg/L p = 0.0057; 12.5 p = 0.0060; Fig. 3).
TABLE I SURVIVORSHIP OF Boana pulchella TADPOLES EXPOSED TO VARIOUS CONCENTRATIONS OF PICLORAM IN THE ACUTE TOXICITY TEST. THE FRACTIONS REPRESENT THE NUMBER OF DEAD INDIVIDUALS DIVIDED BY THE TOTAL NUMBER OF INDIVIDUALS AT THE BEGINNING OF THE EXPERIMENT FOR EACH TREATMENT (IN ALL CASES, VALUES INCLUDE THE ADDITION OF INDIVIDUALS FROM ALL REPLICATES).
| Time (hours) | Treatment | |||||
| Control | T1 | T2 | T3 | T4 | T5 | |
| 0 | 0/16 | 0/16 | 0/16 | 0/16 | 0/16 | 0/16 |
| 24 | 1/16 | 5/16 | 4/16 | 0/16 | 0/16 | 0/16 |
| 48 | 1/16 | 8/16 | 4/16 | 0/16 | 0/16 | 0/16 |
Control (0.0); T1 (16.5); T2 (14.5); T3 (12.5); T4 (10.5); T5 (8.5) mg/L de Picloram.
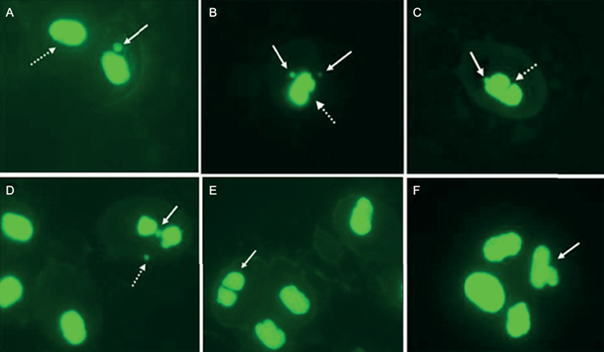
Fig. 1 Erythrocytes of Boana pulchella tadpoles (630X). A: normal cell (dotted arrow) and presence of micronucleus (MN, white arrow). B: Cell with two MN (white arrow) and nuclear aberration: kidney-shaped (dotted white arrow). C: MN (white arrow) and nuclear aberration: notched (dotted white arrow). D: Bridge (white arrow) and MN (dotted arrow). E: binucleate cell (white arrow) F: lobed nucleus (white arrow).
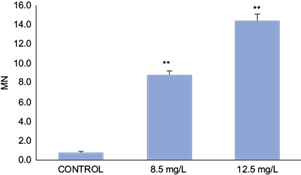
Fig. 2 Micronuclei (MN) frequency (per 1000 erythrocytes) in Boana pulchella tadpoles after 48 h of exposed to different concentrations of Picloram (Tordon 24K). Bars represent the mean ± SEM (standard error of the mean). The significant difference was **p ˂0.01 with respect to the control (binomial proportion test).
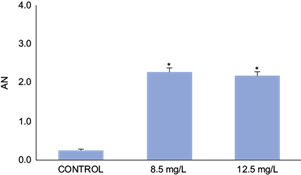
Fig. 3 Frequency of nuclear aberrations (AN) in erythrocytes (per 1000 red blood cells) in Boana pulchella tadpoles at 48 h of exposed to different concentrations of Picloram (Tordon 24K). Bars represent the mean ± SEM (standard error of the mean). The significant difference was *p ˂0.05 with respect to the control (binomial proportion test).
DISCUSSION
The value obtained for CL5048h of PIC in the present work (16.5 mg/L) is similar to that reported for different aquatic organisms; being moderately toxic as well as for fish (LC5096h 19.3 mg/L in rainbow trout, 14.5 mg/L in bluegill sunfish and 55 mg/L in fathead minnow) and invertebrates such us Daphnia magna, CL5048h 50 mg/L (EXTOXNET 2022). Studies carried out on different species of amphibians show ranges of toxicity that run from very toxic to slightly toxic. For neotropical species, the LC5048h determined for PIC in Rhinella arenarum tadpoles was 0.025 mg/L (Lajmanovich et al. 2013), while for Rhinella marina and Physalaemus centralis the LC5096h was 0.30 mg/L and 0.51 mg/L, respectively (Figueiredo and de Jesús Rodrigues 2014). Likewise, another study on PIC toxicity in Australian Limnodynastidae anurans suggests moderate toxicity for Adelotus brevis (123.0 mg/L) and Limnodynastes peronii (116.0 mg/L) tadpoles (Johnson 1976). In this regard, although the variability in the toxicity of pesticides has already been reported for different species of amphibians (Wojtaszek et al. 2004, Izaguirre et al. 2006, Relyea and Jones 2009, Agostini 2013) the different ranges of sensitivity are especially relevant when dealing with species that live in the same habitats and are therefore exposed to the same local pressures (Natale et al. 2018).
Another aspect to consider is the use of different commercial formulas. Substances present in commercial pesticide formulations that are not active ingredients (API) are considered a priori inert excipients (Cox and Surgan 2006) and fulfill a variety of functions, being able to act as solvents, surfactants, preservatives, and stabilizers, among others. Different studies have revealed that the excipients modify the ecotoxicity of pesticides, in terms of the toxicity and availability of the API, resulting many times more toxic than the active principle itself (Cox and Surgan 2006, Könen and Çavaş 2008, Peltzer et al. 2013). Taking these differences into account, it is important to use commercial pesticide formulations in ecotoxicological tests to avoid underestimating effects (García-Ortega et al. 2006, Lajmanovich et al. 2013). In particular, concerning B. pulchella, many of the previous evaluations in tadpoles have focused mainly on insecticide toxicity (Agostini et al. 2010, Pérez-Iglesias et al. 2014, Natale et al. 2018). Our results support the idea that this species presents a differential range of sensitivity for different exposure times and complement the information available about the toxicity of many of the most frequently used pesticides in the region.
Amphibian tadpoles are organisms susceptible to genetic damage even at low concentrations of xenobiotic contaminants and during short exposure times (Ralph et al. 1996, Clements et al. 1997). In this regard, previous studies have documented that B. pulchella can be affected by different pesticides used in agricultural environments at various stages of its development. Pérez-Iglesias et al. (2015), evidenced genotoxicity of the herbicide imazethapyr by observing an increase in the frequency of MN and the formation of AN in tadpoles exposed for 96 hours in concentrations ranging from 0.39 to 1.17 mg/L. Brodeur et al. (2012) evaluated the impacts of the herbicide glyphosate on adult specimens of B. pulchella, determining that, after its application, an intense drought is generated on the agricultural landscape that produces oxidative stress in males and females, inducing DNA damage, increasing the formation of MN. On the other hand, an increase in the frequency of nuclear anomalies or aberrations reflects surveillance mechanisms and reactions to eliminate cells with genetic damage (Cavaş and Ergene-Gözükara 2003). In this sense, these authors provide information about the cellular state since it has been reported that the frequency of lobulated cells or cells without a nucleus present is increased under stressful situations such as diet changes, pathologies, or metabolic disorders, among others (Lajmanovich et al. 2013). Likewise, pyknotic nuclei (with highly condensed chromatin) are associated with DNA damage and apoptosis, occurring at high levels in response to cell injury (Lajmanovich et al. 2013). The presence of kidney-type AN may be associated with a clastogenic effect (Brzuzan et al. 2006) and those of the binucleate type are indicators of cytotoxicity (Cavaş and Ergene-Gözükara 2005). Our results showed a significant increase in the frequency of AN in all the treatments tested at sublethal concentrations, which would indicate the clastogenic and cytotoxic of PIC by the presence of AN, mainly binucleate and kidney-shaped. Being consistent with the previous evaluations that demonstrate the genotoxicity caused by PIC.
In summary, this study represents the first evidence of the genotoxicity of the picloram herbicide in Boana pulchella tadpoles under controlled laboratory conditions. Considering that amphibian population declines can be caused by alterations in the habitat and by chemical pollution, being herbicides used in agricultural systems among the most relevant contaminants, the results agree with previous studies on the need to carry out ecotoxicological evaluations to identify and characterize the hazards associated with their use. In this sense, our results increase the information about picloram, which could be useful for its management.
Based on the results obtained and considering that Boana pulchella is not a threatened species, it has a wide distribution in the Uruguayan territory and in the Pampa Biome, it could be considered as a reference organism in environmental monitoring programs in the region.

