











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Los bosques de encino, o bosques de Quercus L., son comunidades vegetales propias de las zonas montañosas y, junto con los bosques de coníferas, son representativos de áreas de clima templado (Kappelle, 2006). De acuerdo con Rzedowski (1978), los bosques de encino se distribuyen en zonas montañosas del país, en un rango entre 1200 y 2800 m s.n.m., temperatura media anual entre 12 a 20 °C y una precipitación de 600 a 1200 mm. Prospera de manera general en ambientes templados y subhúmedos. En el estado de Puebla, los encinares cubren una extensión de 167,423 ha (4.9% del territorio político estatal) y se distribuye de norte a sur de la entidad, principalmente en las provincias fisiográficas Sierra Madre Oriental, Eje Neovolcánico Transversal y Sierra Madre del Sur (Yanes, 2011).
Entre las Áreas Naturales Protegidas decretadas en el estado de Puebla, la Reserva Estatal Sierra del Tentzo (REST) conserva la mayor superficie de bosque de Quercus. La REST abarca 57,815.28 ha y está integrada parcialmente por 13 municipios. Dentro de la REST, el bosque de Quercus cubre la mayor parte, con 32,570.28 ha (53.5%), lo que representa casi 20% de dicha cubierta forestal en el estado de Puebla (SMADSOT, s.f.).
Respecto a elementos florísticos, la REST ha sido poco explorada a pesar de su cercanía a vías de comunicación. Por ejemplo, Martínez (1998) y Hernández (2002) estudiaron parte de la Sierra del Tentzo, ubicada al sureste de la ciudad de Puebla y parte alta de la cuenca alta del río Atoyac. En estas obras se reporta la presencia de bosque tropical caducifolio, bosque de Quercus, matorral xerófilo, palmar, bosque de galería y asociaciones secundarias. También hay estudios que incluyen parte de elementos de la REST, ya sea mediante el estudio de una familia en particular (Flores, 2023) o mediante el descubrimiento de nuevas especies para la ciencia (Alvarado-Cárdenas et al., 2024). Por lo tanto, resulta relevante efectuar un mayor esfuerzo de recolecta que permita un mejor conocimiento de la riqueza florística de esta Área Natural Protegida. Esto con el fin de conocer aquellas especies que están catalogadas en alguna categoría de riesgo, endémicas o nativas del territorio nacional; o bien, pueden ser cruciales para el diseño de estrategias de conservación.
Los objetivos de este trabajo fueron 1) contribuir al conocimiento de la flora del estado de Puebla mediante un inventario florístico de un área localizada al este de la REST, 2) estimar su riqueza florística y 3) evaluar sus afinidades florísticas con otras áreas del país que permitan apreciar el valor biológico para su conservación.
Materiales y Métodos
Área de estudio
El área de estudio delimitada tiene 927.6 ha, con un intervalo elevacional de 1922 a 2350 m. Se ubica en la cordillera Tentzo (Castillo y Martínez, 2014), dentro del municipio Tzicatlacoyan, que forma parte del polígono del Área Natural Protegida Reserva Estatal Sierra del Tentzo (Fig. 1), donde se desarrollan elementos de bosque templado en su parte más alta. El área estudiada ha sido poco explorada florísticamente en la región. En este sentido el inventario florístico puede ser un referente para mejores prácticas en el manejo de sus recursos bióticos.
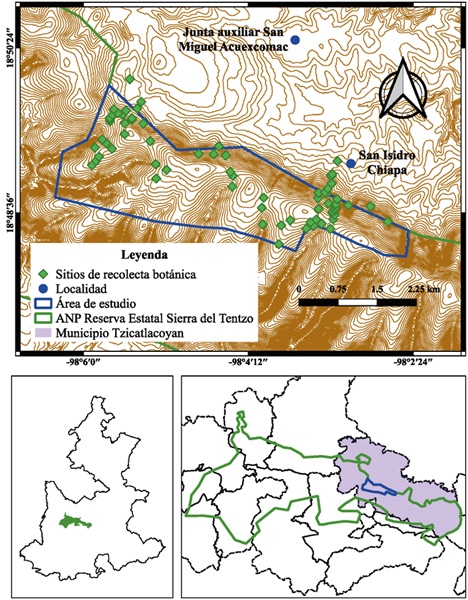
Figura 1: Área de estudio localizada en la Cordillera Tentzo, Área Natural Protegida Reserva Estatal Sierra del Tentzo (REST), Tzicatlacoyan, Puebla, México. Elaboración: José Andrés Cruz-Martínez.
El área estudiada se sitúa en la cordillera Tentzo (Fig. 2), en los límites de las provincias biogeográficas Cuenca del Balsas y Faja Volcánica Transmexicana (Morrone et al., 2017) y tiene su origen en afloramiento recifal que se remonta hacia el Cretácico Inferior (Barremiano-Albiano). Superficialmente está constituido por rocas calizas de la Formación Morelos (Morón et al., 2000; Zepeda, 2013; Castillo y Martínez, 2014).

Figura 2: Cordillera Tentzo, Tzicatlacoyan, Puebla, México. A. vista panorámica general de la Cordillera Tentzo; B. ladera norte con presencia de Quercus L., Juniperus L., Malacomeles (Decne.) Decne., Agave L. y Yucca L.; C. ladera norte con presencia de Agave L. y Brahea Mart. Fotografías: José Andrés Cruz-Martínez.
El clima que predomina es templado subhúmedo (C(w1)), y en menor medida semicálido subhúmedo del grupo C ((A)C(w1)) (García y CONABIO, 1998; García, 2004). La temperatura media anual es de 16.1 °C y la precipitación media anual es de 757.7 mm (SMN, 2025). El tipo de suelo mejor representado donde se desarrolla el bosque de Quercus es el leptosol, seguido del vertisol (INEGI, 2024; Ortiz y Villaseñor, 2025). Existe un uso ganadero del suelo y un área de 860 ha está destinado al programa de Pago por Servicios Ambientales (PSA) por medio de la Comisión Nacional Forestal (CONAFOR).
Riqueza florística
Se efectúo una búsqueda exhaustiva de las especies vegetales recolectadas en la REST a través de colecciones digitalizadas del Instituto de Biología de la Universidad Nacional Autónoma de México (Murguía-Romero et al., 2024), del Sistema Nacional de Información sobre Biodiversidad (CONABIO, 2025) y de la Red de Herbarios Mexicanos (RHM, 2025), además de contribuciones florísticas previas (Martínez, 1998; Hernández, 2002; Flores, 2023; Alvarado-Cárdenas et al., 2024).
Se realizaron 29 salidas al campo entre septiembre de 2023 y enero del 2025 para la recolecta de ejemplares botánicos. El mayor esfuerzo de muestreo se llevó a cabo en la época de lluvias. Se procuró recolectar cuatro o cinco duplicados preferentemente con estructuras reproductoras. Los materiales se procesaron de acuerdo con las técnicas indicadas por Lot y Chiang (1986). Se tomaron datos ecológicos, geográficos y características de los materiales recolectados.
El material fue determinado por medio de literatura especializada: Flora de Guerrero (Diego-Pérez, 2004; Castro-Mendoza y Fonseca, 2012; Fonseca y Medina-Lemos, 2012; Diego-Escobar et al., 2013; Velázquez, 2018), Flora de Tehuacán-Cuicatlán (Alvarado-Cárdenas, 2004; Medina-Lemos y Fonseca, 2009; Calónico-Soto, 2011; Martínez-Gordillo et al., 2019; García-Mendoza, 2024), Flora del Bajío y de Regiones Adyacentes (Rzedowski y Calderón de Rzedowski, 1997, 1999; Carranza, 2007; Romero et al., 2014; Bedolla et al., 2024), Flora de Veracruz (Zanoni, 1982; Espejo-Serna y López-Ferrari, 2003, 2005; Villarreal y Villaseñor, 2004; Martínez-Bernal et al., 2008), Flora Fanerogámica del Valle de México (Rzedowski y Calderón de Rzedowski, 2005), Árboles y arbustos de México (Standley, 1920), así como artículos y tesis que abordaran algún tratamiento taxonómico (Vázquez, 1992; Jimeno, 2008; Flores, 2013; Valencia-A. et al., 2015; Soto, 2020; Murillo-Pérez y Rodríguez, 2021). Para el caso de plantas que no pudieron ser determinadas taxonómicamente se recurrió al apoyo de especialistas en ciertas familias.
Los materiales determinados se cotejaron con las colecciones digitalizadas del Instituto de Biología de la Universidad Nacional Autónoma de México (MEXU) y Red de Herbarios Mexicanos (HUAP, IBUG, RSA, entre otros) (acrónimos según Thiers, 2025). Confirmada la determinación, se corroboró la correcta escritura de la especie y nombre de la autoridad en Plants of the World Online (POWO, 2025). Para el caso de Lycopodiopsida y Polypodiopsida se consultó la página Pteridoportal (PCC, 2025). El material recolectado será depositado en los herbarios FEZA, HUAP y MEXU.
El inventario florístico se ordenó con base en PPG I (2016) para Lycopodiopsida y Polypodiopsida, Christenhusz et al. (2011) para gimnospermas y APG IV (2016) para Angiospermas. Para complementar el inventario, se añadió la siguiente información: hábito, distribución geográfica y tipos de vegetación en los que se encuentran. El hábito se tomó con base en Moreno (1984), excepto las plantas con hojas agrupadas en rosetas y las cactáceas, que se consideraron como otras formas de crecimiento (Gutiérrez y Solano, 2014). La distribución geográfica se consideró a partir de la propuesta de Rzedowski (1991a): México, Megaméxico I, Megaméxico II, Megaméxico III, así como distribución continental y amplia distribución. Su hábitat se consideró de acuerdo con el tipo de vegetación (Rzedowski, 1978) en donde se les reporta, tanto de literatura especializada como de ejemplares de herbarios. Además, el listado florístico se cotejó con la Norma Oficial Mexicana NOM-059-SEMARNAT-2010 (SEMARNAT, 2010) para obtener aquellas especies con estatus de conservación.
Para estimar la riqueza florística y calcular cuántas especies faltarían por recolectarse en el área de estudio, se construyó una matriz de presencia-ausencia con base en los eventos de recolecta. La matriz obtenida fue analizada con el programa EstimateS v. 9.1 (Colwell, 2025) y los estimadores no paramétricos Incidence-based Coverage Estimator (ICE, por sus siglas en inglés) (Chao y Chiu, 2016) y Chao2 (Chao, 1984). El porcentaje de la riqueza conocida respecto a la estimada se obtuvo por medio de la fórmula: (elementos observados/elementos estimados) ×100, para cada índice no paramétrico.
Se ejecutó un análisis de similitud florística a nivel genérico, se comparó el área de estudio con 14 trabajos florísticos (Hernández, 2002; Castillo-Argüero et al., 2004; Cabrera-Luna y Gómez-Sánchez, 2005; Encina et al., 2009; Díaz-Cruz et al., 2011; León de la Luz et al., 2012; Trejo-Díaz y Tejero-Díez, 2017; Morales, 2019; Rojas-Martínez y Flores-Olvera, 2019; Cerros-Tlatilpa et al., 2020; Aragón-Parada et al., 2021; Gutiérrez et al., 2021; Harker et al., 2021; Valentín-Martínez et al., 2024), realizados en el país. Se consideraron estudios con reporte de bosque de encino, matorral xerófilo y bosque tropical caducifolio. Se construyó una matriz básica de datos de presencia-ausencia con 949 géneros y 15 áreas geográficas (Unidades Geográficas Operativas). La matriz fue analizada con el programa de cómputo NTSYS v. 2.02 (Rohlf, 1998). Con base en el coeficiente de similitud de Jaccard, se calculó la matriz de similitud y con esta última las áreas se agruparon por medio del método Media Aritmética no Ponderada (UPGMA, por sus siglas en inglés) para la construcción del fenograma (Gutiérrez y Solano, 2014).
Resultados
Riqueza florística
El área de estudio localizada en la REST está representada por 339 especies, 226 géneros y 76 familias (Apéndice 1, Figs. 3, 4, 5, 6). Las Angiospermas resultaron ser el grupo con la mayor riqueza florística: las Eudicotiledóneas registran 269 especies (79.4%), seguidas por las Monocotiledóneas con 58 (17.1%) y Magnólidas con 2 (0.6%). En menor proporción están las Polypodiopsida, Lycopodiopsida y Pinidae (Cuadro 1).

Figura 3: Algunas especies presentes en el área de estudio, Puebla, México. A. Selaginella pallescens (C. Presl) Spring (Selaginellaceae); B. Pleopeltis thyssanolepis (A. Braun ex Klotzsch) E.G. Andrews & Windham (Polypodiaceae); C. Argyrochosma formosa (Liebm.) Windham; D. Bommeria ehrenbergiana (Kl.) Underw.; E. Pellaea atropurpurea (L.) Link; F. Pellaea ovata (Desv.) Weath. (Pteridaceae); G. Juniperus flaccida Schltdl. (Cupressaceae); H. Peperomia bracteata A.W. Hill (Piperaceae); I. Nothoscordum bivalve (L.) Britton var. bivalve (Amaryllidaceae); J. Brahea dulcis (Kunth) Mart. (Arecaceae); K. Agave salmiana Otto ex Salm-Dyck; L. Dasylirion acrotrichum (Schiede) Zucc.; M. Echeandia flavescens (Schult. & Schult.f.) Cruden; N. Echeandia graminea M. Martens & Galeotti; O. Echeandia nana (Baker) Cruden; P. Echeandia reflexa (Cav.) Rose (Asparagaceae). Fotografías: José Andrés Cruz-Martínez.

Figura 4: Algunas especies presentes en el área de estudio. A. Echeandia vestita (Baker) Cruden; B. Yucca periculosa Baker (Asparagaceae); C. Thyrsanthemum floribundum (M. Martens & Galeotti) Pichon (Commelinaceae); D. Hypoxis mexicana Schult. & Schult.f. (Hypoxidaceae); E. Tigridia illecebrosa Cruden (Iridaceae); F. Deiregyne tenorioi Soto Arenas & Salazar; G. Habenaria strictissima Rchb.f.; H. Laelia albida Bateman ex Lindl.; I. Laelia autumnalis (Lex.) Lindl.; J. Malaxis javesiae (Rchb.f.) Ames (Orchidaceae); K. Heteranthera rotundifolia (Kunth) Griseb. (Pontederiaceae); L. Carlowrightia pringlei B.L. Rob. & Greenm.; M. Elytraria imbricata (Vahl) Pers.; N. Pseuderanthemum praecox (Benth.) Leonard; O. Ruellia lactea Cav.; P. Stenandrium dulce (Cav.) Nees (Acanthaceae). Fotografías: José Andrés Cruz-Martínez.

Figura 5: Algunas especies presentes en el área de estudio. A. Rhus standleyi F.A. Barkley (Anacardiaceae); B. Asclepias glaucescens Kunth; C. Asclepias linaria Cav.; D. Asclepias mexicana Cav.; E. Cascabela thevetioides (Kunth) Lippold; F. Funastrum elegans (Decne.) Schltr. (Apocynaceae); G. Dahlia coccinea Cav.; H. Grindelia inuloides Willd. var. inuloides; I. Tagetes lucida Cav. (Asteraceae); J. Berberis moranensis Hebenstr. & Ludw. ex Schult. & Schult.f. (Berberidaceae); K. Wimmeria microphylla Radlk. (Celastraceae); L. Ipomoea murucoides Roem. & Schult.; M. Ipomoea stans Cav. (Convolvulaceae); N. Echeveria heterosepala Rose (Crassulaceae); O. Machaonia hahniana Baill. (Rubiaceae); P. Sideroxylon salicifolium (L.) Lam. (Sapotaceae). Fotografías: José Andrés Cruz-Martínez.

Figura 6: Algunas especies presentes en el área de estudio de los cuales solo se cuenta con registro fotográfico. A. Agave lechuguilla Torr.; B. Agave potatorum Zucc. (Asparagaceae); C. Asclepias lynchiana Fishbein; D. Cynanchum foetidum (Cav.) Kunth (Apocynaceae); E. Coryphantha pallida Britton & Rose; F. Ferocactus latispinus (Haw.) Britton & Rose; G. Mammillaria haageana Pfeiff.; H. Opuntia pubescens H.L. Wendl. ex Pfeiff. (Cactaceae). Fotografías: José Andrés Cruz-Martínez.
Cuadro 1: Riqueza florística catalogada en el área de estudio localizado en la Reserva Estatal Sierra del Tentzo (REST), Tzicatlacoyan, Puebla, México. Valor entre paréntesis=porcentaje.
| Grupo taxonómico | Familias | Géneros | Especies |
| Lycopodiopsida | 1(1.3) | 1(0.4) | 1(0.3) |
| Polypodiopsida | 2(2.6) | 6(2.7) | 8(2.4) |
| Pinidae | 1(1.3) | 1(0.4) | 1(0.3) |
| Angiospermas | |||
| Magnólidas | 2(2.6) | 2(0.9) | 2(0.6) |
| Monocotiledóneas | 12(15.8) | 41(18.1) | 58(17.1) |
| Eudicotiledóneas | 58(76.3) | 175(77.4) | 2669(79.4) |
| Total | 76 | 226 | 339 |
Las diez familias con mayor riqueza de especies agrupan 51.9% de lo reconocido en el área de estudio (Cuadro 2). Destacan las familias Asteraceae (46 especies), Fabaceae (37), Lamiaceae (13), Poaceae (12), Apocynaceae (12), Euphorbiaceae (12), Orchidaceae (11), Asparagaceae (11), Convolvulaceae (11) y Solanaceae (11) (Cuadro 2). Por otro lado, 13 géneros agrupan 20.4% de las especies presentes en el área de estudio (Cuadro 3). Los géneros más diversos fueron Salvia L. (10 especies), Ipomoea L. (7), Tillandsia L. (6), Echeandia Ortega (5), Asclepias L. (5), Euphorbia L. (5), Senna Mill. (5), Quercus L. (5), Solanum L. (5), Stevia Cav. (4), Vachellia Wight & Arn. (4), Phoradendron Nutt. (4) y Physalis L. (4).
Cuadro 2: Familias con mayor cantidad de géneros y especies reportadas en el área de estudio localizado en la Reserva Estatal Sierra del Tentzo (REST), Tzicatlacoyan, Puebla, México.
| Familia | Géneros | No. de especies | % |
| Asteraceae | 33 | 46 | 13.6 |
| Fabaceae | 20 | 37 | 10.9 |
| Lamiaceae | 4 | 13 | 3.8 |
| Poaceae | 10 | 12 | 3.5 |
| Apcynaceae | 8 | 12 | 3.5 |
| Euphorbiaceae | 7 | 12 | 3.5 |
| Orchidaceae | 9 | 11 | 3.2 |
| Asparagaceae | 5 | 11 | 3.2 |
| Convolvulaceae | 4 | 11 | 3.2 |
| Solanaceae | 4 | 11 | 3.2 |
| Total | 104 | 176 | 51.9 |
Cuadro 3: Géneros con mayor riqueza de especies en el área de estudio localizado en la Reserva Estatal Sierra del Tentzo (REST), Tzicatlacoyan, Puebla, México. Valor entre paréntesis=porcentaje.
| Géneros | No. de especies (%) |
| Salvia L. | 10(2.9) |
| Ipomoea L. | 7(2.1) |
| Tillandsia L. | 6(1.8) |
| Echeandia Ortega | 5(1.5) |
| Asclepias L. | 5(1.5) |
| Euphorbia L. | 5(1.5) |
| Senna Mill. | 5(1.5) |
| Quercus L. | 5(1.5) |
| Solanum L. | 5(1.5) |
| Stevia Cav. | 4(1.2) |
| Total | 57(17) |
De los elementos reconocidos, el hábito dominante fue el herbáceo (208 especies), seguido por arbustos (87), árboles (32) y 12 especies se consideraron como otras formas de crecimiento. La distribución geográfica de las especies reconocidas reveló que 139 de estas son endémicas de México, 17 corresponden a Megaméxico I, 40 a Megaméxico II y 16 a Megaméxico III. Ciento dos especies tienen una distribución en gran parte de América y 12 son de amplia distribución. Ocho especies se registraron como exóticas: Digitaria ciliaris (Retz.) Koeler, Diplotaxis muralis (L.) DC., Erodium cicutarium (L.) L’Hér., Lysimachia arvensis (L.) U. Manns & Anderb., Malva nicaeensis All., Melinis repens (Willd.) Zizka, Oxalis corniculata L. y Portulaca oleracea L.
Seis especies se incluyen en alguna categoría de riesgo según la NOM-059-SEMARNAT-2010 (SEMARNAT, 2010): Mammillaria haageana Pfeiff. se cataloga en peligro de extinción, tres especies son consideradas como amenazadas (Bouvardia erecta (DC.) Standl., Dasylirion acrotrichum (Schiede) Zucc. y Hesperalbizia occidentalis (Brandegee) Barneby & J.W. Grimes), mientras que Brahea dulcis (Kunth) Mart. y Laelia autumnalis (Lex.) Lindl. están sujetas a protección especial.
La mayoría de las especies se reportan en más de un tipo de vegetación. Para el bosque de Quercus se reportan 299 especies, 290 en matorral xerófilo y 243 en bosque tropical caducifolio (Apéndice 1, Fig. 7). Un número reducido de especies solo se han reportado para un tipo de vegetación: Brongniartia vicioides M. Martens & Galeotti, Cladocolea tehuacanensis (Oliv.) Tiegh. y Echeveria heterosepala Rose en matorral xerófilo y Nissolia leiogyne Sandwith en bosque tropical caducifolio.
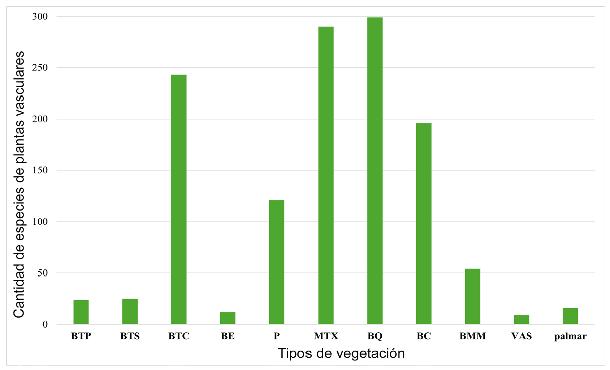
Figura 7: Especies de plantas vasculares presentes en el área de estudio reportados en los tipos de vegetación propuestos por Rzedowski (1978). Tipo de vegetación. BTP=bosque tropical perennifolio, BTS=bosque tropical subcaducifolio, BTC=bosque tropical caducifolio, BE=bosque espinoso, P=pastizal, MTX=matorral xerófilo, BQ=bosque de encino, BC=bosque de coníferas, BMM=bosque mesófilo de montaña, VAS=vegetación acuática y subacuática, otro tipo de vegetación=palmar.
Curva de acumulación
Los estimadores no paramétricos ICE y Chao2 revelaron que hasta el momento se ha recolectado entre 73.0 y 74.6% de las especies que conforman la flora vascular del área de estudio (Fig. 8). Faltarían por recolectarse entre 115 y 125 especies, por lo que el listado completo podría ascender a una cantidad entre 453 y 463 especies. Tres nuevos registros se reportan para el estado de Puebla: Asclepias elata Benth., Govenia lagenophora Lindl. y Sarcoglottis assurgens (Rchb.f.) Schltr. (Fig. 9).
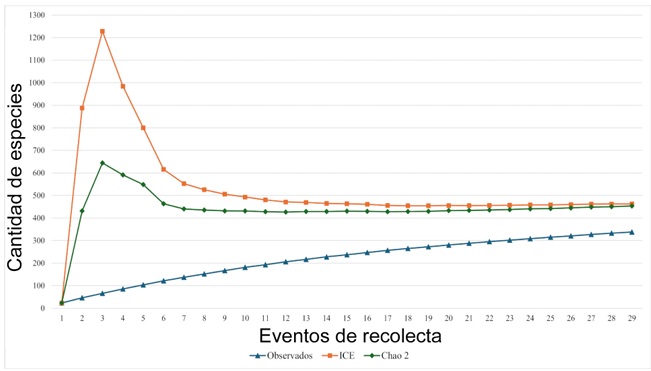
Figura 8: Estimación de la riqueza florística del área de estudio por medio de dos estimadores no paramétricos, ICE y Chao2.

Similitud florística
La flora de la Cordillera Tentzo se comparó a nivel genérico con 14 localidades florísticas del país (Apéndice 2). El análisis de agrupamiento mostró dos grupos: el grupo I integró dos localidades florísticas con presencia de elementos de bosque de encino templado; estas fueron la Sierra de Zapalinamé, Coahuila (Encina et al., 2009) y Sierra de las Ánimas, Estado de México (Trejo-Díaz y Tejero-Díez, 2017) (Fig. 10). Estas dos zonas comparten una similitud de 51% y presentan una similitud de 33.5 (76 géneros) y 33.9% (76 géneros respectivamente) con respecto a la cordillera Tentzo (Apéndice 2). La Sierra de Zapalinamé y Sierra de las Ánimas comparten 30 géneros; entre estos se encuentran Dalea L., Phoradendron Nutt., Quercus, Salvia, Selaginella P. Beauv., Tagetes L. y Tillandsia. De manera general, estas dos localidades florísticas se caracterizan, además del tipo de vegetación, por climas con régimen de lluvias en verano y temperatura media anual entre 12 y 18 °C (García, 2004) (Cuadro 4). El grupo II integró diversas localidades florísticas, donde se reconocieron tres subgrupos (Fig. 10). El subgrupo “a” se integró con Nocupétaro, Michoacán (Valentín-Martínez et al., 2024) y La Salada, Colima (Harker et al., 2021). Estas dos localidades presentan una similitud de 41.5% en comparación con el área de estudio; son similares a nivel genérico en 29.8 y 35.4%, respectivamente (Apéndice 2). Por otro lado, las localidades que se incluyeron en el subgrupo “b” tienen una similitud de 44% y fueron (en orden ascendente en la similitud genérica con el área de estudio) La Cañada, Querétaro (Cabrera-Luna y Gómez-Sánchez, 2005) (42.2%), Cerro El Pelado, Puebla (Rojas-Martínez y Flores-Olvera, 2019) (44%), Santo Domingo Huehuetlán, Puebla (Hernández, 2002) (50.4%), Península de Baja California Sur (León de la Luz et al., 2012) (52.2%) y Cerros La Cantera y Delgado, Morelos (Cerros-Tlatilpa et al., 2020) (52.4%) (Apéndice 2). Las localidades florísticas que se integraron en estos dos subgrupos tienen en común climas con temperatura media anual mayor a 18 °C, régimen de lluvias en verano y sequía invernal marcada frecuente en ecosistemas de clima cálidos en donde se distribuye vegetación de tropical, bajo cociente P/T (seco o semiseco) (García, 2004), en gradientes menores a 2000 m s.n.m., así como la presencia de suelos leptosoles y regosoles, aspectos que permiten la distribución del bosque tropical caducifolio (Cuadro 4). Las siete localidades tienen en común la ocurrencia de 15 géneros, entre estos Bursera Jacq. ex L., Ipomoea, Mimosa L., Opuntia Mill., Porophyllum Guett., Salvia y Tillandsia.
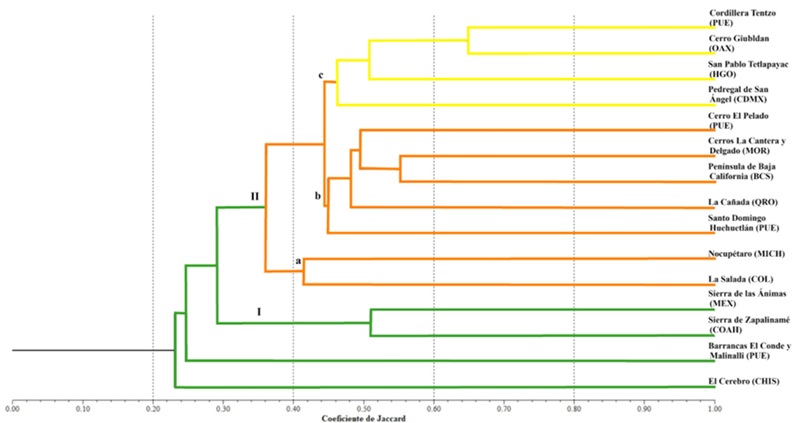
Figura 10: Fenograma de similitud florística entre el área estudiada (Cordillera Tentzo) y otras 14 localidades florísticas presentes en México. Entidades federativas: BCS=Baja California Sur, CDMX=Ciudad de México, CHIS=Chiapas, COAH=Coahuila, COL=Colima, HGO=Hidalgo, MEX=Estado de México, MICH=Michoacán, MOR=Morelos, OAX=Oaxaca, PUE=Puebla, QRO=Querétaro.
Cuadro 4: Comparación del área de estudio (Cordillera Tentzo) con respecto a otras comunidades florísticas en México con base en la similitud florística efectuada a nivel genérico. Provincia fisiográfica (PF): EN=Eje Neovolcánico Transversal, PBC=Península de Baja California, SCyG=Sierras de Chiapas y Guatemala, SMOr=Sierra Madre Oriental, SMS=Sierra Madre del Sur. Provincia biogeográfica (PB): BC=Baja California, CB=Cuenca del Balsas, DC=Desierto Chihuahuense, FVT=Faja Volcánica Transmexicana, SMOr=Sierra Madre Oriental, TBP=Tierras Bajas del Pacífico, V=Veracruzana. Tipo(s) de vegetación (TV): BQ=bosque de Quercus, BTC=bosque tropical caducifolio, MTX=matorral xerófilo.
| Localidades | Área | PF | PB | Clima | Tipo de suelo | TV | Elevación | Taxa | % | ||
| (km2) | (m) | Familias | Géneros | Especies | similitud | ||||||
| Cordillera Tentzo | 9.2 | EN | CB, FVT | C(w1), (A)C(w1) | leptosol, vertisol | Q, MTX, TC | 1922-2350 | 76 | 224 | 337 | 100.0 |
| Cerro Giubldan (Aragón-Parada et al., 2021) | 21.0 | SMS | CB | BShw, Cw | leptosol, luvisol, phaeozem | Q, MTX | 1600-2930 | 100 | 306 | 535 | 64.9 |
| Cerros La Cantera y Delgado (Cerros-Tlatilpa et al., 2020) | 0.4 | EN | CB | Aw0 | leptosol, regosol | TC | 1390-1530 | 85 | 286 | 434 | 52.4 |
| Península de Baja California (León de la Luz et al., 2012) | 3.3 | PBC | C | BS0, BS1 | regosol | TC | 500-1500 | 101 | 359 | 645 | 52.2 |
| San Pablo Tetlapayac (Morales, 2019) | 10.0 | SMOr | SMOr | BS0hw | leptosol, regosol | MTX | 1000-2012 | 57 | 219 | 296 | 50.9 |
| Santo Domingo Huehuetlán (Hernández, 2002) | 54.0 | EN | CB, FVT | BS1 | leptosol | TC | 1400-1800 | 78 | 234 | 300 | 50.4 |
| Pedregal de San Ángel (Castillo-Argüero et al., 2004) | 1.77 | EN | FVT | Cb(w1)(w) | leptosol | MTX | 2200-2277 | 74 | 198 | 337 | 48.0 |
| Cerro El Pelado (Rojas-Martínez y Flores-Olvera, 2019) | 18.7 | EN, SMS | CB | BS1(h')w | leptosol | TC | 1187-1631 | 67 | 225 | 339 | 44.0 |
| La Cañada (Cabrera-Luna y Gómez-Sánchez, 2005) | 0.2 | EN | DC | BS1k | leptosol | MTX, TC | 1850-2030 | 76 | 187 | 264 | 42.2 |
| La Salada (Harker et al., 2021) | 2.9 | SMS | TBP | Aw0 | regosol, vertisol | TC | 370-600 | 78 | 245 | 408 | 35.4 |
| Sierra de las Ánimas (Trejo-Díaz y Tejero-Díez, 2017) | 6.0 | EN | FVT | Cb(w1)(w) | andosol, leptosol, luvisol, phaeozem, vertisol | Q | 3010-3070 | 75 | 176 | 271 | 33.9 |
| Sierra de Zapalinamé (Encina et al., 2009) | 29.0 | SMOr | SMOr | BSkw, C(w0) | leptosol, phaeozem, xerosol | Q | 1590-3140 | 67 | 178 | 259 | 33.5 |
| Nocupétaro (Valentín-Martínez et al., 2024) | 0.1 | SMS | CB | BS(h')(w) | luvisol, regosol | TC | 680-759 | 56 | 134 | 173 | 29.8 |
| Barrancas El Conde y Malinalli (Gutiérrez et al., 2021) | ----- | EN | FVT | C(w1), C(w2) | durisol, luvisol | Q | 2060-2680 | 40 | 80 | 105 | 21.9 |
| El Cerebro (Díaz-Cruz et al., 2011) | 21.7 | SCG | V | Aw2 | litosol | Q | 860 | 54 | 120 | 150 | 20.1 |
Por último, el subgrupo “c” (Fig. 10) se conformó por el Pedregal de San Ángel, Ciudad de México (Castillo-Argüero et al., 2004), San Pablo Tetlapayac, Hidalgo (Morales, 2019), Cerro Giubldan, Oaxaca (Aragón-Parada et al., 2021) y la Cordillera Tentzo, Puebla, con una similitud de 48%. Estas comunidades tienen en común que su flora se desarrolla en clima árido sobre suelos leptosoles, condiciones que permiten la presencia de elementos florísticos de matorral xerófilo (Cuadro 4). Las comunidades que integran este último subgrupo tienen en común la presencia de 57 géneros, entre otros Asclepias, Calcicola W.R. Anderson & C. Davis, Echeandia, Echeveria DC., Euphorbia, Montanoa Cerv. y Villadia Rose. La flora genérica del Cerro Giubldan fue la de mayor similitud con la Cordillera Tentzo (64.9%) (Apéndice 2), al compartir 145 géneros.
Discusión
Riqueza florística
Este trabajo florístico permite un acercamiento al conocimiento de la flora del municipio Tzicatlacoyan, y un complemento para la REST, en conjunto con las contribuciones florísticas de Martínez (1998), Hernández (2002), Flores (2023) y Alvarado-Cárdenas et al. (2024), quienes reportan elementos de la REST de los municipios Huehuetlán el Grande y Puebla. La riqueza florística aquí reportada representa 4.96% de la flora del estado de Puebla (Villaseñor y Ortiz, 2025). Así mismo, con base en una revisión exhaustiva de colecciones digitalizadas y disponibles en el repositorio del Sistema Nacional de Información sobre Biodiversidad (SNIB), la Red de Herbarios Mexicanos (RHM), y contribuciones como la de Martínez (1998) y Alvarado-Cárdenas et al. (2024), se documentan un total de 669 especies de plantas vasculares, incluidas en 396 géneros y 95 familias para la REST.
Con base en nuestros resultados, 145 especies son registros nuevos para la REST, situación que eleva su riqueza florística a 814 especies. En este mismo sentido las adiciones a nivel de familia para la Reserva Estatal fueron Araliaceae, Berberidaceae, Brassicaceae, Dioscoreaceae, Iridaceae, Linaceae, Loganiaceae, Oxalidaceae, Pontederiaceae, Portulacaceae y Violaceae, además de 54 nuevos registros a nivel genérico, como Aralia L., Berberis L., Dioscorea Plum. ex L., Hechtia Klotzsch, Laelia Lindl., Malaxis Sol. ex Sw., Peperomia Ruiz & Pav., Stillingia L., Tigridia Juss. y Villadia.
Las diez familias con más especies de plantas vasculares (Cuadro 2) no presentan en lo general el mismo patrón de distribución a nivel nacional (Villaseñor y Ortiz, 2025). Asteraceae es la familia con mayor riqueza florística y de amplitud geográfica en México y en el mundo (Redonda-Martínez, 2022). Esta particularidad puede explicarse por su exitosa capacidad de dispersión (Villaseñor, 2018). Euphorbiaceae y Apocynaceae presentan distribución subcosmopolita principalmente en regiones tropicales y subtropicales del mundo; Euphorbiacae con menor ocurrencia en zonas templadas de ambos hemisferios (Steinmann, 2002), y Apocynaceae en México tiene uno de los centros de mayor diversidad (Alvarado-Cárdenas et al., 2020). La presencia de ambas familias en la Cordillera Tentzo posiblemente obedece a su influencia por el ambiente con clima cálido y permite prosperar bosque tropical caducifolio. En contraste, Poaceae habita en prácticamente todos los ecosistemas del mundo; en México no es la excepción (Sánchez-Ken, 2019).
A nivel genérico, los mejor representados en el área de estudio son Salvia, Ipomoea, Tillandsia, Euphorbia, Echeandia, Asclepias L., Senna Mill., Solanum, Stevia Cav. y Vachellia Wight & Arn., Phoradendron y Physalis. Según Hedge (1992) y Ramamoorthy y Elliott (1998), Salvia tiene mayor presencia en áreas de influencia laurásica o neártica. En México Salvia es considerado el género con mayor riqueza de especies (330; Villaseñor y Ortiz, 2025). Su distribución se presenta principalmente en bosques templados y menos frecuentemente en bosques tropicales caducifolios y subcaducifolios, así como matorrales xerófilos (Ramamoorthy y Elliot, 1998). Con excepción de los bosques tropicales subcaducifolios los otros ecosistemas se presentan en la REST. Ipomoea está bien representado en la región pantropical, posiblemente por su buena adaptación a distintas condiciones climáticas, edafológicas y topográficas en diversos tipos de vegetación (Carranza, 2004; Alcántar-Mejía et al., 2012). Echeandia es endémico del continente americano y en México se presenta un centro de diversificación (Rodríguez y Ortiz-Catedral, 2013; Rodríguez y Ortiz-Brunel, 2019), con amplia presencia en la Zona de Transición Mexicana (Ortiz-Brunel et al., 2021). La presencia de cinco especies en el área de estudio posiblemente responde a la posición geográfica cercana a esta zona biogeográfica.
Con respecto al hábito, la REST está representada con 61.7% de las especies herbáceas. Este resultado coincide con el patrón reportado para México por Villaseñor y Ortiz (2014), quienes señalaron que este hábito es el más común en la flora mexicana, seguido por el arbustivo y el arbóreo. Respecto al endemismo, 62.5% de las especies son exclusivas del territorio nacional. Esta cifra es congruente en proporción con lo reportado para el nivel nacional, donde cerca de 50% de las especies son endémicas (Villaseñor, 2016). Sin embargo, el número puede aumentar a medida de un mejor conocimiento de la flora (Rzedowski, 1991a). La conformación de la vegetación actual de México y sus endemismos tienen diversas interpretaciones; entre estas destacan las fluctuaciones climáticas durante el Terciario, la posición geográfica del país, procesos orogénicos y el vulcanismo (González-Medrano, 1996, 2012; González, 2012; Villarreal-Quintanilla et al., 2017). Bajo estas condiciones ambientales y fisiográficas la parte septentrional del país funcionó como un centro de diversificación de gran número de plantas vasculares adaptadas a la aridez, que más tarde se expandieron hacia el sur y hacia el norte del territorio nacional (Rzedowski, 1962; 1991b).
La vegetación del área de estudio revela una composición heterogénea, donde la mayoría de las especies (299) se ha documentado para bosque de Quercus. Sin embargo, diversos elementos florísticos se reportan para otros ecosistemas descritos por Rzedowski (1978), como matorrales xerófilos (290) y bosques tropicales caducifolios (243). Morón et al. (2000) señalaron que la vegetación del Tentzo está conformada por una transición entre bosque de Quercus, matorral xerófilo y bosque tropical caducifolio. Sin embargo, la composición florística refleja una heterogeneidad que sugiere sea definida como un área de transición (ecotono), donde convergen elementos florísticos característicos de tres tipos de vegetación.
Curva de acumulación
Con base en los estimadores no paramétricos ICE y Chao2, se observa que se ha generado 73 y 74.6% del conocimiento florístico, respectivamente. En este sentido se considera que el área de estudio ha sido bien muestreada como lo sugieren Rojas-Parra et al. (2003), una vez que las curvas generadas son próximas entre sí y muestran un comportamiento asintótico. Sin embargo, se sugiere continuar con la exploración botánica, actividad que podría detectar más especies para las que no se había reportado su distribución en el estado de Puebla (Fig. 9), como ocurre en este estudio con Asclepias elata Benth., Govenia lagenophora Lindl. y Sarcoglottis assurgens (Rchb.f.) Schltr., o bien, en la descripción de nuevas especies para la flora mexicana, como Gonolobus volcanicus Nuñez Oberg, L.O. Alvarado et S. Islas (Apocynaceae) (Alvarado-Cárdenas et al., 2024), conocida únicamente en la REST.
Similitud florística
Este análisis reveló la afinidad entre las áreas que presentan ecosistemas con similitudes ecológicas, reconociéndose dos grandes grupos. El grupo I incluye dos comunidades con presencia de géneros con distribución típica de clima templado (Fig. 10). Existen ciertas excepciones, como Desmodium Desv., que presenta distribución ecológica más amplia en vegetación tropical (Rzedowski y Calderón de Rzedowski, 2016) y Phoradendron, género americano, con presencia en regiones templadas y tropicales (Alvarado-Cárdenas, 2010). Quercus, elemento boreal con presencia principal en regiones montañosas de México (Romero et al., 2014), donde forma comunidades homogéneas o heterogéneas con pinos, u otros elementos florísticos presentes en bosques tropicales caducifolios y matorrales xerófilos (Vázquez-Villagrán, 2000). Un elemento epífito común en ambas comunidades es Tillandsia, cuya presencia en las regiones tropicales y subtropicales en México es frecuente (Diego-Escobar et al., 2013).
El grupo II integró la mayoría de las comunidades consideradas en este estudio, donde los subgrupos “a” y “b” se caracterizan por presentar ecosistemas de bosque tropical caducifolio. Los dos subgrupos comparten 19 géneros. Entre estos se encuentran Bursera, elemento común de este tipo vegetación, con pocas especies presentes en matorrales xerófilos (Medina-Lemos, 2008), Jatropha L., que tiene su centro de diversidad en regiones tropicales, semiáridas y áridas de América (Martínez-Gordillo et al., 2014) y Tetramerium Nees, en regiones subtropicales y tropicales del hemisferio occidental (Daniel, 2024).
El subgrupo “c” conjuntó cuatro localidades, entre las cuales está el área aquí estudiada. Este subgrupo se distingue por concentrar elementos florísticos característicos de matorral xerófilo, donde resaltan 57 géneros. Entre estos destacan Mammillaria Haw., con presencia de la mayoría de sus especies en zonas semiáridas de México (Bravo-Hollis y Sánchez-Mejorada, 1991), Agave L., con la mayoría de sus especies endémicas de México (García-Mendoza, 2011) y Mimosa, que concentra en México 20% de la riqueza del género, con distribución en zonas tropicales, áridas y semiáridas (Grether et al., 2006; Martínez-Bernal et al, 2008).
La comunidad de Santo Domingo Huehuetlán (Hernández, 2002), a pesar de su cercanía con el área de estudio (Cordillera Tentzo) y localizada en parte en la REST, difiere florísticamente, al relacionarse en mayor proporción con las comunidades del subgrupo “b”, que con excepción de la Península de Baja California, Baja California Sur son más cercanas geográficamente, grupo II (Fig. 10) (Apéndice 2). Además, en la porción norte de Santo Domingo Huehuetlán se expone la Formación Morelos con origen en el Cretácico Inferior y presencia de roca caliza, mientras que en la porción centro y sur aflora el Complejo Acatlán, con origen en el Paleozoico Inferior (Hernández, 2002). El Complejo Acatlán es considerado el basamento del Terreno Mixteco, compuesto por rocas metamórficas (Castillo y Martínez, 2014), y es en este tipo de sustrato rocoso donde los bosques tropicales caducifolios generalmente se establecen (Rzedowski, 1978).
El análisis de similitud también reflejó que la composición florística a nivel genérico de la cordillera Tentzo tuvo mayor relación con áreas de matorral xerófilo. Esta similitud posiblemente se debe a las condiciones edáficas y climáticas semejantes con las comunidades del subgrupo “c”. Comparte con el cerro Giubldan, Oaxaca (Aragón-Parada et al., 2021), San Pablo Tetlapayac, Hidalgo (Morales, 2019) y el Pedregal de San Ángel, Ciudad de México (Castillo-Argüero et al., 2004), suelos de tipo leptosol, caracterizados por ser someros, pedregosos y poco desarrollados, así como la presencia de roca caliza. Además, se asocian a sitios de compleja orografía, frecuente en México y que explica la amplia distribución de los matorrales xerófilos (Solleiro y Gama, 2011). Bautista et al. (2009) mencionaron que los suelos de tipo leptosol son comunes en zonas áridas y en ellos se desarrolla el matorral xerófilo. Estos suelos originados de afloramientos rocosos se relacionan con el desarrollo de plantas que toleran condiciones de aridez, asimismo, bosques tropicales caducifolios y en menor proporción bosques de Quercus (INEGI, 2015). Estas características edáficas posiblemente permiten el desarrollo de una vegetación que favorece la similitud entre el Cerro Giubldan, Oaxaca, y la Cordillera Tentzo, Puebla (64.9%, 146 géneros). El cerro Giubldan presenta bosque de Quercus y matorral xerófilo, con predominancia del primero de estos. Aragón-Parada et al. (2021) reconocieron para esta comunidad el bosque de encino perennifolio y bosque de encino caducifolio. Este último coincide con el área de estudio, al presentar un dosel abierto, dominado por especies caducifolias de Quercus que desprenden su follaje en los meses de enero a mayo. Algunas especies compartidas en ambas comunidades fueron Aralia humilis Cav., Bouvardia multiflora (Cav.) Schult. & Schult.f., Bursera fagaroides (Kunth) Engl., Brongniartia vicioides M. Martens & Galeotti, Clinopodium mexicanum (Benth.) Govaerts, Ipomoea murucoides Roem. & Schult., Q. deserticola Trel., Juniperus flaccida Schltdl., Salvia pubescens Benth., Trichostema purpusii Brandegee y Varronia curassavica Jacq. (Aragón Parada, 2015).
El matorral xerófilo está bien representado principalmente en el altiplano mexicano y se desarrolla en áreas geológica y geográficamente complejas. La escasez de precipitación donde se desarrolla este tipo de vegetación obedece, entre diversos factores, al efecto de continentalidad y el surgimiento de las Sierra Madre Occidental y Oriental que funcionan como barreras que impiden el paso de humedad hacia el interior de territorio mexicano (González-Medrano, 2012). En la Sierra del Tentzo la presencia de matorral xerófilo posiblemente es resultado de transformaciones geológicas y movimientos orogénicos ocurridas a finales del Cuaternario, dadas por eventos de erosión remontante (Brunet, 1967) que formaron las principales sierras de la cuenca del Balsas. Por otro lado, la aridez presente en la Cordillera Tentzo es una respuesta a la sombra orográfica dada por la sierra de Zongolica como ocurre con el Valle de Tehuacán-Cuicatlán (Miguel-Talonia et al., 2014).
En la cordillera Tentzo la presencia de elementos de bosque tropical caducifolio al parecer se remonta a un poco más de 20 Ma, cuando se desarrolló la flora del Terciario de Norteamérica, como lo sugieren Pérez-García et al. (2012). La presencia de elementos típicos de bosque tropical caducifolio en el área estudiada presenta una fuerte influencia florística típica del Reino Neotropical (Rzedowski, 1991a). Esta característica fortalece la hipótesis de una comunidad heterogénea al presentarse elementos de clima seco y templado, que aquí denominamos ecotono (Bader et al., 2007). Estas áreas por lo común presentan mayor riqueza al combinar especies provenientes de distintas comunidades vecinas y con mayor homogeneidad florística (Odum, 1971; Bader et al., 2007).
En el área de estudio hay elementos que resultaron característicos por su abundancia, entre ellos, Quercus deserticola Trel. que se asocia con Yucca L. (Romero et al., 2014), en este estudio con Y. periculosa Baker. Por otro lado, Quercus grahamii Oerst. ex Trel., frecuente en ecotonos de bosques tropicales caducifolios, asociado a Brahea dulcis (Kunth) Mart. y en suelos someros derivados de roca caliza (Valencia-Á. et al., 2015), condiciones presentes en el área estudiada. Situación similar ocurre con Rhus standleyi F.A. Barkley, con presencia en matorrales xerófilos (Rzedowski y Calderón de Rzedowski, 1999) y bosques de Quercus (Medina-Lemos y Fonseca, 2009).
Conclusión
Se propone que la vegetación del área estudiada debe ser reconocida como un ecotono. Los nuevos reportes florísticos para el estado de Puebla enriquecen el conocimiento de la flora para esta entidad federativa, y fortalecen las bases para el manejo y conservación de los recursos vegetales de la región.

