











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introduction
The species of the genus Apiospora Sacc. are a group of microscopic fungi of the family Apiosporaceae within the phylum Ascomycota. In Index Fungorum (2024), from 1875 to 2024, there are 182 records, which correspond to 157 recognized species in the genus.
Although the members of Apiospora have been reported to have a cosmopolitan distribution, occurring in terrestrial and marine environments (Klemke et al., 2004), these species are considered uncommon and occasionally understudied. In one study, for example, Kwon et al. (2022) found 242 isolates of nine Apiospora species in different bamboo organs in Korean forests. The lifestyle of Apiospora specimens is variable, mainly saprobes (Crous et al., 2021; Tian et al., 2021), including coprophiles (Alfatafta et al., 1994), but there are symbiotic (Wang et al., 2017) and endophytic species (Park et al., 2020; Liao et al., 2023), as well as plant (Liao et al., 2022, 2023; Agustí-Brisach et al., 2023) and human pathogens (Paul et al., 2024).
Studying a phytopathogenic alga from blackberry stems (Cortés Martínez et al., 2023), we isolated a fungus with colony morphology and spores similar to Apiospora or Arthrinium Kunze. Conidia and conidiophores are difficult to distinguish among the species of those genera (Pintos and Alvarado, 2021; Liao et al., 2023). Even Arthrinium was previously considered to be the sexual phase of Apiospora (Pintos et al., 2019). It was not until the study by Pintos and Alvarado (2021), with sequences from four molecular markers (ITS, 28S, TEF1-α, tub2) and phylogenetic analyses, that it was possible to define both genera as sister lineages.
In Mexico, to our knowledge, there are no reports of Apiospora species. The closest taxon is Arthrinium sp., which was detected in samples from glacier ice on the volcano Iztaccíhuatl at 5010 m a.s.l. by Calvillo-Medina et al. (2020).
The objective of this study was to identify the fungus that was isolated from the stem of a blackberry plant (Rubus 'Tupy') by means of phylogenetic analysis of the sequences of the internal transcribed spacer (ITS) and the translation elongation factor 1-alpha gene (TEF1-α), and determine whether it represents a pathogenic organism of blackberry.
Materials and Methods
Collection site and isolation of fungus
The fungus under study was isolated from the blackberry stems of Rubus 'Tupy' affected by orange blotch (Cortés Martínez et al., 2023). The material was collected on April 4, 2019, in the locality of Taretan, Michoacán, Mexico. The climate in the area is humid subtropical with summer rains, annual rainfall of 1560 millimeters and temperatures ranging from 14.4 to 29.7 ºC (INAFED, 2020).
Blackberry stem fragments (0.5 cm2), after three seconds of vortexing (Scientific Industries, G560 SI-0236, Vernon Hills, USA) in 96% alcohol, were rinsed with distilled water, vortexed with 3% chlorine for three seconds (Agrios, 2005), and rinsed three times with sterile distilled water. The material was then placed on potato dextrose agar (PDA, MCD Lab, Mexico) in darkness. The unique isolate obtained of this fungus was purified and labeled as INE47. Samples of the isolate were preserved in 30% glycerin and maintained at 4 oC in the “Colección de organismos benéficos y patógenos del Instituto de Ecología, A.C. (INECOL), Centro Regional del Bajío” in Pátzcuaro, Michoacán, Mexico.
DNA extraction and sequencing
For molecular identification, the total DNA was extracted from the fresh isolate, using the Wizard® Genomic DNA Purification Kit (Promega, USA) according to manufacturer´s instructions. The internal transcribed spacer 1 (ITS1), 5.8S rDNA and ITS2 were amplified by PCR with the primers ITS1F/ITS4 (White et al., 1990; Gardes and Bruns, 1993). Also, the translation elongation factor 1-alpha (TEF1-α) was amplified with primers EF1-688F/EF1-1251R (Alves et al., 2008). In both cases, ITS and TEF1-α, the 25 μl PCR mixture contained 2.5 μl of DNA, 2.5 μl of buffer 5X colorless, 1 μl of 50 mM MgCl2, 1 μl of 10 mM dNTPs, 1 μl of 10 μM forward primer, 1 μl of 10 μM reverse primer, 0.2 μl of 5U Taq polymerase (Promega, USA) and 15.8 µl of sterilized water. ITS PCR cycling parameters were as follows: one cycle at 94 °C for 3 min, followed by 30 cycles with a denaturation step at 94 °C for 30 sec, an annealing step at 50 °C for 30 sec, an extension step at 72 °C for 1.15 min, and a final extension at 72 °C for 7 min. The TEF1-α amplification program consisted of one cycle at 95 °C for 5 min, followed by 30 cycles with a denaturation step at 94 °C for 30 sec, an annealing step at 55 °C for 45 sec, an extension step at 72 °C for 90 sec, and a final extension at 72 °C for 10 min, as described by Phillips et al. (2005). Amplified PCR products were sent to Macrogen (Seoul, Korea) for purification and sequencing reactions. Fungal sequences were deposited in GenBank (2024).
Phylogenetic analyses
The fungal ITS and TEF1-α sequences were assembled using BioEdit v. 7.2.5 (Hall, 1999). A phylogenetic analysis was performed integrating an ITS rDNA matrix (ITS1-5,8S-ITS2 region) and also a second matrix for the TEF1-α marker, with related sequences of the Apiospora species referred in Crous et al. (2015), Kwon et al. (2021), Tian et al. (2021), and the most similar published species-sequences deposited in NCBI (2024) (Table 1), including IOM 325295 of Arthrinium sp. from the Iztaccíhuatl glacier (Calvillo-Medina et al., 2020) in order to know whether it is related to INE47. Sequences from Arthrinium were added and Nigrospora guangdongensis L.Y. Tian, a sister taxon of Apiospora and Arthrinium, was used to root the tree. Sequences in each matrix were aligned using the Mafft program v. 7 (Katoh et al., 2019). The molecular evolution that best fits each data set was determined using jModel test v. 0.1.1 (Posada, 2008). Phylogenetic inference analyses were performed by the Maximum Likelihood (ML) method with 1000 replicates of bootstrap in the program MEGA v. 11.0.10 (Tamura et al., 2021), and Bayesian inference (BI) using the program MrBayes v. 3.2.7a (Ronquist et al., 2012) for 1000000 replicates. The phylogenies from ML and BI analyses were displayed in MEGA v. 11.0.10 (Tamura et al., 2021) or FigTree v. 1.4.3 (Rambaut, 2016), respectively.
Table 1: Sequences and GenBank (2024) accession numbers of Apiospora Sacc. species used for phylogenetic analyses. Nigrospora Zimm. and Arthrinium Kunze sequences were used as outgroup. Samples added in this study are indicated in bold. (-) Information not available.
| Species | Strain/voucher | Country | Substrate/Host | ITS accession number | TEF1-α accession number |
| Apiospora aquatica (Luo, Hyde & Su) Pintos & Alvarado | CC.XQDXG234 | China | Neyraudia reynaudiana (Kunth) Keng ex Hitchc. | PP407937.1 | PP496624.1 |
| Apiospora arundinis (Corda) Pintos & Alvarado | MUCL 14429 | Japan | - | AB220280.1 | - |
| Apiospora bambusicola (Tang, Hyde & Kang) Tian & Tibpromma | MFLU 20-0528 | Thailand | Bamboo | NR_171983.1 | - |
| Apiospora bambusicola | MFLUCC 20-0144 | Thailand | Dead culms of bamboo | MW173030.1 | MW183262.1 |
| Apiospora chiangraiense Tian & Tibpromma | MFLU:21-0046 | Thailand | Bamboo and maize | MZ542520.1 | - |
| Apiospora chiangraiense | KUNCC 24-17543 | China | Poaceae sp. | PP584687.1 | PP933194.1 |
| Apiospora intestini Kajale, Sonawane & Roh. Sharma) Pintos & Alvarado | IRAN 4331C | Iran | Quercus brantii Lindl. | PP464101.1 | PP565758.1 |
| Apiospora intestini | CBS 135835 | India | Grasshopper | NR_154698.1 | KR011351.1 |
| Apiospora intestini | INE47 | Mexico | Blackberry | PQ459871 | PQ468997 |
| Apiospora italica (Pintos & Alvarado) Pintos & Alvarado | AP221017 | Italy | Arundo donax L. | MK014880.1 | MK017956.1 |
| Apiospora jatrophae (Sharma, Kulk. & Shouche) Pintos & Alvarado | CBS:134262 | India | Jatropha podagrica Hook. | NR 154675.1 | - |
| Apiospora jiangxiensis (Wang & Cai) Pintos & Alvarado | CC.HLG100.4 | China | An unidentified woody plan | PP407913.1 | PP496602.1 |
| Apiospora lageniformis Kwon & Kim | KUC21686 | South Korea | Phyllostachys nigra var. henonis (Mitford) Stapf ex Rendle | ON764022.1 | ON806626.1 |
| Apiospora montagnei Sacc. | AP101020 | Spain | Arundo donax | ON692409.1 | - |
| Apiospora montagnei | AP301120 | Spain | Arundo micrantha Lam. | ON692408.1 | ON677182.1 |
| Apiospora ovata (Crous) Pintos & Alvarado | CBS 115042 | Hong Kong | Arundinaria hindsii Munro | NR 121558.1 | KF145037.1 |
| Apiospora paraphaeosperma (Senan. & Hyde) Pintos & Alvarado | KUC21688 | South Korea | Bamboo | ON764025.1 | ON806629.1 |
| Apiospora pseudosinensis (Crous) Pintos & Alvarado | CBS:135459 | Netherlands | Bamboo | NR_121559.1 | KF145044.1 |
| Apiospora rasikravindrae (S.M. Singh, Yadav, P.N. Singh, Rah. Sharma & S.K. Singh) Pintos & Alvarado | MFLU:23-0237 | Thailand | Ananas comosus (L.) Merr. | OR438411.1 | OR500340.1 |
| Apiospora saccharicola (Stevens) Pintos & Alvarado | CBS:463.83 | Netherlands | Phragmites australis (Cav.) Trin. ex Steud. | KF144921.1 | KF145053.1 |
| Apiospora serenensis (Larrondo & Calvo) Pintos & Alvarado | SFC20220920-B248 | South Korea | Sea sand | OP597860.1 | - |
| Apiospora sphaerosperma Pers.) Pintos & P. Alvarado | CBS:114314 | Iran | Hordeum vulgare L. | KF144904.1 | KF145038.1 |
| Arthrinium caricicola Kunze & Schmidt | CBS:145903 | Germany | Carex ericetorum Pollich | NR 175149.1 | |
| Arthrinium crenatum Pintos, Alvarado & Gardiennet | CBS 146353 | France | Festuca burgundiana Auquier & Kerguélen | NR_175664.1 | MW221917.1 |
| Arthrinium pusillispermum Kwon, Jang & Kim | KCTC 46906 | South Korea | Seaweed | NR_177480.1 | - |
| Arthrinium pusillispermum | KUC21321 | South Korea | Seaweed | MH498533.2 | MN868930.1 |
| Arthrinium sp. | IOM 325295 | Mexico | Iztaccíhuatl Glacier | MH890621.1 | - |
| Nigrospora guangdongensis Tian | CFCC 53917 | China | - | NR_174814.1 | MT024493.1 |
Morphological characterization
The mycelial colony was characterized in PDA and in malt-extract agar media (MEA, MCD Lab, Mexico) after two weeks of being maintained at 25 oC in darkness. The colors of its front and rear in the Petri dish were registered using as a reference the color codes of the Methuen handbook (Kornerup and Wanscher, 1983). Semipermanent preparations were made with PVLG (polyvinyl alcohol-glycerol) and 3% KOH to observe the micromorphological structures with a Carl Zeiss optical microscope (Primostar 1, Göttingen, Germany). Size, shape, and color of the spores, conidiophores and conidiogenic cells were recorded in a Leica optical microscope (Serie Galen III, Buffalo, USA). The Zeiss Zen Model 2012 software and an AxioCam ERc 5s camera (Göttingen, Germany) were used to take pictures.
Pathogenicity test
To test the capacity of producing damage in the stems and leaves, fresh mycelia of INE47 were inoculated in detached healthy tissues of blackberry (Rubus 'Tupy'). The surface of stems and leaves were disinfested with 70% ethanol for 30 sec, then washed with sterilized water (da Silva et al., 2016). Leaves and stems were wounded with a puncture (1 mm diameter) made with a sterilized dissecting needle (1 mm). Then, 5 mm agar mycelium squares were inoculated on the puncture. In addition, an inoculum square was placed on an area of the tissue without lesion. Controls were inoculated just with an agar square without mycelia in a punctured and an unpunctured area. The test was performed in triplicate and maintained in a humidity chamber (80-100% humidity) for eight days at 25 °C, with a light period of 8 h. Next, if a lesion was observed on tissues, the disease severity was assessed by measuring the lengths of the spots. To confirm whether the lesion was caused for the inoculated organism, small fragments (1-2 mm) of the affected tissues were placed on PDA media. Once the fungus grew, fungal colony and conidia were compared with originally inoculated Apiospora to conclude the Koch's postulates.
Results
Molecular identification
Ensambled fungal ITS and TEF1-α sequences from INE47 were uploaded to GenBank (2024) under accession number PQ459871 (ITS) and PQ468997 (TEF1-α). Comparison of the ITS sequence with the BLAST (2024) tool in NCBI shows its high similarity with Arthrinium intestini Kajale, Sonawane & Roh. Sharma, whose accepted name is Apiospora intestini (Kajale, Sonawane & Roh. Sharma) Pintos & P. Alvarado (culture from holotype: CBS 135835, accession number NR_154698.1), with 99.63% of identity and 96% query coverage, as well as to Apiospora intestini (strain IRAN 4331C, accession number PP464101.1) with 99.81%, and 91% of query coverage. The second closest species is Arthrinium pusillispermum S.L. Kwon, S. Jang & J.J. Kim (holotype, KCTC 46906m, accession number NR_177480.1), with 99.43% of identity and 92% of query coverage. Whereas the TEF1-α sequence of INE47 is close to Apiospora intestini (holotype, CBS 135835, KR011351.1), with 99.44% of identity, this sequence comes from the grasshopper gut, isolated in India, while the second closest sequence, with 98.49% percentage identity, is Arthrinium intestini (MZ546406.1), whose host are dead culms of bamboo from Thailand.
In the phylogenetic analyses, a total of 47 sequences (28 ITS and 19 TEF1-α) were incorporated, including the two sequences obtained in this research. The data matrix corresponding to the ITS1+5.8S+ITS2 marker database consisted of 28 sequences, with a 595 bp length. On the other hand, 19 sequences were integrated in the TEF1-α database, with a total alignment length of 552 bp. For the ITS data, the best fitting evolutionary model was TIM3ef+I+G, while for the TEF1-α sequences it was TrNef+G.
Phylogenetic trees based on the TEF1-α sequence showed higher resolution in the Apiospora intestini clade than those based on ITS analysis. The concatenated sequence analysis based on ITS and TEF1-α sequences (1147 characters, including gaps) showed a similar topology in Maximum Likelihood and Bayesian analyses, with the latter showing higher resolution (Fig. 1).
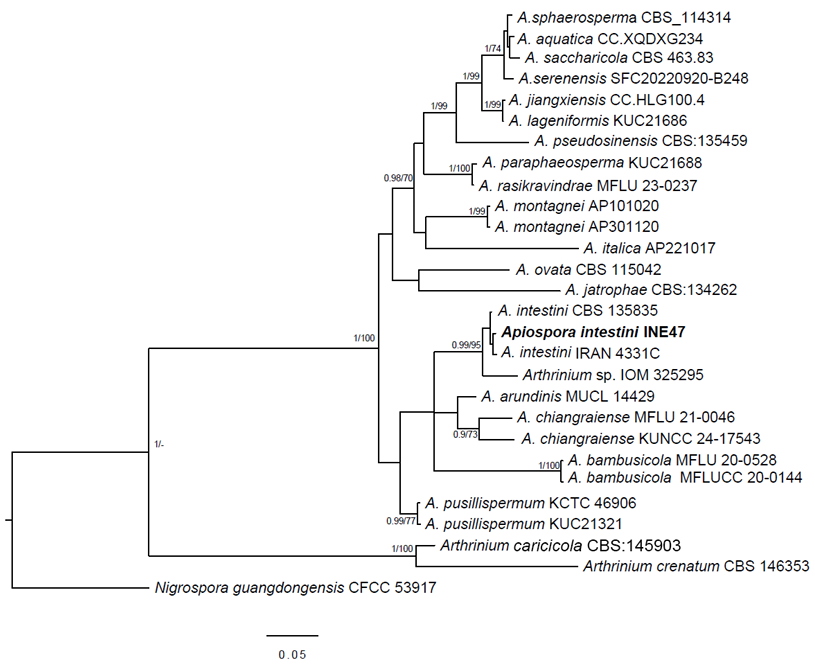
Figure 1: Multigene phylogenetic analysis constructed with Bayesian inference from a concatenated ITS and TEF1-α dataset. Nigrospora guangdongensis L.Y. Tian was used to root the tree. Bayesian posterior probability ≥0.95 and bootstrap ≥70 ML are indicated near the nodes. The Apiospora intestini (Kajale, Sonawane & Roh. Sharma) Pintos & P. Alvarado sequence from this study is indicated in bold letters.
Through the multigene analysis of the ITS and TEF1-α, INE47 was identified within the Apiospora genus. Arthrinium species were located in the phylogenetic analysis between Apiospora strains and Nigrospora guangdongensis (Fig. 1). INE 47 clustered with A. intestini of the holotype (CBS 135835) from India and the strain from IRAN (4331C) with significant statistical support (BI posterior probability=0.99, ML bootstrap=95). Apiospora arundinis (Corda) Pintos & P. Alvarado and A. chiangraiense X.G. Tian & Tibpromma were the sister clades of INE47.
The glacier sequence (IOM 325295) corresponds to Apiospora and is a sister clade of A. intestini. Additional molecular markers would clarify the species identity of this sequence.
Taxonomy
Apiospora intestini (Kajale, Sonawane & Roh. Sharma) Pintos & P. Alvarado, Fungal Systematics and Evolution 7: 206. 2021. Fig. 2.
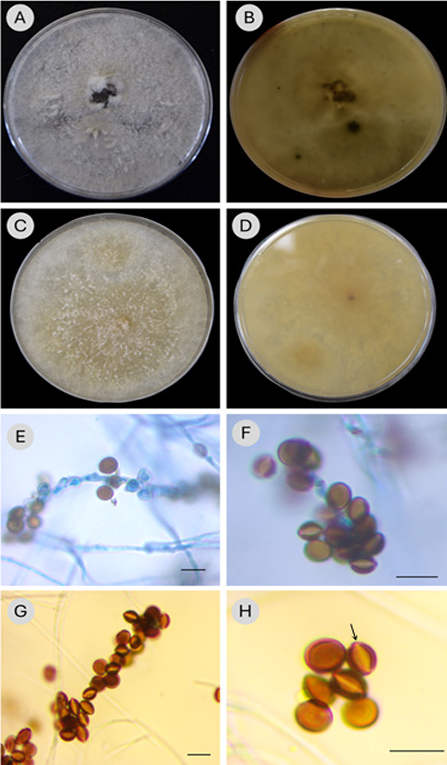
Figure 2: Morphological features of Apiospora intestini (Kajale, Sonawane & Roh. Sharma) Pintos & P. Alvarado (INE47). A-B. culture morphology of colony grown in PDA (A, obverse; B, reverse); C, D. in MEA (C, obverse; D, reverse); E, F. conidiophore and brown conidia, stained with trypan blue 0.05%; G, H. conidia brown, globose or citriform in side view with a scar and lighter slit in the middle part of the conidium (arrow). Scale bars=10 μm.
≡ Arthrinium intestini Kajale, Sonawane & Roh. Sharma (as ´gutiae´), Persoonia 35: 315. 2015.
TYPE: INDIA. Pune, Western Ghats, Maharashtra, gut of grasshopper, 10.I.2013, S. Kajale & M. S. Sonawane (holotype: MCC H1002, culture ex-type MCC 1077 = CBS 135835).
Culture: Mycelium grows fast, covering the entire Petri dish (9 mm) in 3-4 days on PDA and MEA at 25 oC; colony on PDA (Fig. 2A) white, cottony-flat texture, compact, margin irregular, with aerial hyphae forming erect mycelial clusters spreading in some parts of the Petri dish, sporulation scattered, blackish at maturity; reverse side (Fig. 2B) yellowish white to pale yellow (Methuen code: 3A2-3) with spots, concentric circles or olive-brown and greyish-brown rays (Methuen code: 4D3, 4F3 and 5D3); mycelium on MEA (Fig. 2C) beige, loose, scattered over the surface, slightly aerial when forming thin and shorts cords that rise above the surface; pale orange-brown pigments appear as the days go by (Methuen code: 5B2-3); spores forming black spots, aggregates; reverse side (Fig. 2D) yellowish-brown and greyish yellow, with scattered darker brown stains (Methuen code: Bb3-4, 4B3).
Description: Asexual morph: hyphae, thin-walled, smooth, hyaline, septate and branched; conidiophores (Fig. 2E-G) macronematous, hyaline, bunch-shaped, 26-50 × 2 µm, conidia clustered (Fig. 2F, GF); conidiogenous cells short cylindrical and hyaline (2 × (1-)2 µm); conidia aseptate, brown, amerospores, globose in surface view (5-7 × 5-6 µm), citriform in side view (6-7 × 4-5 µm) with a scar and pale slit in the middle part along the length of the conidium, thick wall (Fig. 2G, H). Sexual morph: not observed.
Material examined: MEXICO. Michoacán, municipality Taretan, 1170 m, 19°20'00"N, 101°55'00"W, isolated from blackberry stem affected by Cephaleuros virescens s.l., 4.IV.2019, J. Cortes Martínez and E. Garay Serrano INE47 (IEB).
Pathogenicity test
In the experiment of inoculating mycelium in blackberry stems and leaves (Figs. 3, 4), INE47 only induced disease in the punctured tissue, resulting in oval or rounded necrotic spots with a lesion diameter of 2-4 mm on leaves and 2-5 mm on stems after eight days post-inoculation. The same pathogen was re-isolated (100%) from the circular spots of the affected tissues, proving that A. intestini is the causative agent of these lesions. However, in the vegetal tissues without puncture, there was no damage or stain of disease induced by INE47.

Figure 3: Pathogenicity test with punction on blackberry stems. A-C. INE47 inoculated on punctured area (left side: p+INE47), and control with punctured area without inoculum (right side: cp); D. subepidermal lesion induced by inoculation of INE47; E. control: punctured subepidermal area without inoculum.

Figure 4: Pathogenicity test on blackberry leaves. A-B. INE47 inoculated on non-punctured leaf (np+INE47), showing the adaxial (A) and abaxial side of the same leaf (B), no lesion was induced for the fungus; C. on the left side of the leaf, without injury: control with punction (cp) and, non-punctured (np), while small spots on the right side because of the inoculation of INE47 in a punctured area (p+INE47); D, E. control areas just with punction; F, G. oval and round necrotic lesion caused by INE47 in a punctured area.
Discussion
INE47 detected in the sample of blackberry 'Tupy' was identified as Apiospora intestini, as it clustered with the holotype strain of the species (CBS 135835) and the strain (4331C) of Iran, with significant statistical support (BI/ML = 0.99/95). Morphological characteristics confirmed this identity, because of the conidia shape (brown, amerospores, subglobose in side view with a pale equatorial slit), and the disposition of conidia that form spore bunches, conidiophores macronematous with similar sizes (21.5-50 × 2-2.5 μm) in Crous et al. (2015) and 26-50 × 2 μm for INE47. Moreover, the diameter of conidia is almost the same, in INE47 5-6 μm in surface view, in comparison with the type of A. intestini (4.5-)5.5(-6) μm (Crous et al., 2015), while in lateral view it is thinner in the type ((2-)4(-6) μm) than in INE47 (4-5 μm).
The closest sister species in the phylogeny, Apiospora arundinis (Corda) Pintos & P. Alvarado has spores similar in size: (5-)6-7 μm, in surface view (Crous and Groenewald, 2013), but they are arranged in star-shaped conidiophores rather than in long clusters. However, it is the molecular analyses that mainly support the differentiation of these species (Pintos and Alvarado, 2021) as observed in figure 1, where A. intestini is a sister clade of A. arundinis.
Apiospora intestini was discovered in the gut of a grasshopper in India in 2015 and described as Arthrinium gutiae Kajale, Sonawane, Rohit Sharma (Crous et al., 2015). Subsequently, Pintos and Alvarado (2021), in the separation of Arthrinium and Apiospora species, recognized it under the current name (A. intestini). Since its discovery, few records of this species have been documented, with different lifestyles: as saprobic in dead culms of bamboo in Thailand (Tian et al., 2021), as pathogen on leaves and stems of Quercus brantii Lindl. seedlings, causing very weak symptoms (Bashiri and Abdollahzadeh, 2024), and in Quercus infectoria Oliv. (Ghobadi and Jamali, 2025), both in reports from Iran, and the latest record, as endophyte in the medicinal plant Maytenus emarginata (Willd.) Ding Hou (Shahim et al., 2024). In this study, the disease severity caused by A. intestini on blackberry is limited to a small area. Therefore, at least now, it does not represent an important plant pathogen for this host as it only induces localized necrotic spots in the inoculated areas (Figs. 3, 4). The lesion caused by INE47 is small, and the pathogen is able to establish in the host only through a wound in the stem. Therefore, INE47 could be considered as an opportunistic fungus (Bashiri and Abdollahzadeh, 2024).
There are few reports of Apiospora species in Rosaceae: Apiospora arundinis in rotten loquat fruit (Eriobotrya japonica (Thunb.) Lindl. (Thambugala et al., 2018), and Apiospora dichotomanthi (Wang & Cai) Pintos & Alvarado found on Dichotomanthes tristaniicarpa Kurz (Wang et al., 2018). Therefore, this report represents, to the best of our knowledge, the first record of A. intestini in a Rosaceae species.
The specimen of Apiospora from the Iztaccíhuatl glacier (Calvillo-Medina et al., 2020) and INE47 of this study represent the only two records of the genus in Mexico. The present research extends the worldwide Apiospora intestini distribution and contributes to the knowledge of Mexican fungal diversity, also broadening the understanding of pathogen lifestyle in a new host.

