











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
La introducción de grandes herbívoros domesticados en América significó, en términos ecológicos, la incorporación de nuevos elementos biológicos a los ecosistemas americanos. Este fenómeno fue particularmente relevante en el caso de las especies manejadas en forma de libre pastoreo. En las regiones áridas y semiáridas de América, estos neo-herbívoros han sido introducidos extensamente y su presencia se ha relacionado con cambios en la estructura y dinámica de poblaciones, comunidades y ecosistemas (Bucher, 1982; Fleischner, 1994; Gibbens et al., 2005; Martorell y Peters, 2005; Metzger et al., 2005; Tadey, 2006; Gaitán et al., 2018; Maza-Villalobos et al., 2022).
Uno de los componentes más significativos y característicos de los ecosistemas de las regiones áridas de América son las cactáceas, una familia de plantas suculentas con variadas formas de vida (Bravo-Hollis y Sánchez-Mejorada, 1978, 1991a, b; Hernández-Hernández et al., 2014). Se ha observado que los herbívoros introducidos en regiones áridas y semiáridas afectan la dinámica poblacional de estas plantas, aunque esta influencia es compleja y dependiente de las especies, formas de vida y condiciones locales. Por ejemplo, en el caso de algunas cactáceas globosas de pequeño o mediano tamaño, la presencia de pastoreo no parece estar correlacionada con su densidad (Sclerocactus wrightiae L.D. Benson (Bates et al., 2022; Lariviere et al., 2023)), o puede estarlo positivamente bajo cierto grado de disturbio (p. ej. Mammillaria pectinifera F.A.C. Weber (Martorell y Peters, 2005)), Echinocereus ferreirianus subsp. lindsayi (J. Meyrán) N.P. Taylor (Martorell et al., 2012) y Coryphantha werdermannii Boed. (Martorell et al., 2015).
En las especies de biznagas y cactus columnares el ganado puede causar daños a los individuos por consumo directo (p. ej., Echinocactus platyacanthus Link & Otto (Jiménez-Sierra y Eguiarte, 2010) y Echinopsis terscheckii (Pfeiff.) H. Friedrich & G.D. Rowley (Malo et al., 2011)) y reducir su desempeño reproductivo (E. terscheckii (Peco et al., 2011)), o retrasar su edad de madurez reproductiva (Cephalocereus columna-trajani (Karw.) K. Schum. (Zavala-Hurtado y Díaz-Solís, 1995)).
El reclutamiento de grandes cactáceas puede disminuir también porque la presencia de ganado reduce la abundancia de sus especies nodrizas. Esto parece haber ocurrido con Ferocactus cylindraceus (Engelm.) Orcutt y su especie nodriza Ambrosia dumosa (A. Gray) W. W. Payne, en el Parque Nacional del Gran Cañón, Arizona, EUA (Bowers, 1997; Abella, 2008), o con la interacción entre la especie columnar Carnegiea gigantea (Engelm.) Britton & Rose y sus especies nodrizas Prosopis velutina Wooton, y Parkinsonia microphylla Torr., en el Parque Nacional Saguaro, Arizona, EUA (Conver et al., 2017).
Entre los mecanismos de facilitación propuestos de las especies nodriza sobre las especies de grandes cactus asociadas se encuentran el mejoramiento de variables abióticas, como la modificación del contenido hídrico o nutrientes del suelo, modificación de la temperatura (Valiente-Banuet y Ezcurra, 1991; Carrillo-García et al., 1999) y la reducción de la presión de los consumidores sobre semillas, plántulas o juveniles de las especies facilitadas (McAuliffe, 1984; Suzan et al., 1996). El cardón (Pachycereus pringlei Britton & Rose) es, junto con el saguaro (Carnegiea gigantea), uno de los dos grandes cactus columnares del Desierto Sonorense y el cactus representativo de la península de Baja California, donde se distribuye a lo largo de la mayor parte de su región desértica. En su rango de distribución la ganadería de libre pastoreo ha estado presente desde la introducción de la misma, a finales del siglo XVII, con la fundación de las primeras misiones jesuitas (del Barco, ca. 1780). Por tanto, en mayor o menor medida, las poblaciones de cardón de la península de Baja California han interaccionado con el ganado desde hace tres siglos.
Aunque se ha encontrado que el cardón forma parte ocasional de la dieta de algunos animales como chivas (León de la Luz y Domínguez-Cadena, 2006; Ramírez-Orduña et al., 2008; Medina-Córdova et al., 2014; Montes-Sánchez et al., 2021) y vacas (Montes-Sánchez et al., 2021), no se ha referido daño aparente directo sobre la superficie de los individuos adultos debido a ello. En estudios previamente realizados sobre la dinámica y estructura poblacional del cardón a distintas escalas espaciales y temporales en la península de Baja California no se ha analizado el efecto del ganado en las poblaciones de cardón (Valiente-B. et al., 1995; Silva-Pereyra, 1996; Medel-Narváez, 2003; Bullock et al., 2005; Medel-Narváez et al., 2006; Félix-Burruel, 2012; Delgado-Fernández et al., 2016, 2017; Seminario-Peña, 2020). El ganado podría afectar el reclutamiento del cardón a través de la destrucción directa de plántulas por consumo y pisoteo, o por reducción de la abundancia de nodrizas específicas.
Por otro lado, se ha observado relación positiva entre el reclutamiento del cardón y leguminosas arbóreas (p. ej., palofierro: Olneya tesota A. Gray), y varias especies de mezquite (Prosopis glandulosa Torr., P. articulata S. Watson), asociada a su función como lugares de percha o anidamiento de aves dispersoras de sus semillas, la mejora nutricional del suelo o la protección frente a consumidores de semillas o plántulas (Tewksbury y Petrovich, 1994; Suzán et al., 1996; Carrillo-García et al., 1999; West et al., 2000; Sosa y Fleming, 2002; Suzán-Azpiri y Sosa, 2006).
En el presente estudio nos proponemos explorar la potencial influencia que estos dos factores, presencia de ganado y disponibilidad de leguminosas nodrizas, pueden tener sobre la dinámica poblacional del cardón. Para ello, utilizamos como sistema de estudio un experimento no planificado e iniciado en 1984 que consistió en la exclusión de ganado y otras actividades humanas en el cardonal de la actual Reserva Biológica Dra. Laura Arriaga (Centro de Investigaciones Biológicas del Noroeste, La Paz, Baja California Sur, México). Nos preguntamos si la exclusión del ganado afectó la probabilidad de reclutamiento del cardón y si la facilitación de leguminosas nodrizas es influida por la presencia o exclusión de ganado. La primera hipótesis que exploramos fue que la exclusión de ganado elimina la potencial destrucción de reclutas por pisoteo o consumo y, por tanto, aumenta la probabilidad de reclutamiento del cardón. Nuestra segunda hipótesis fue que el reclutamiento, tanto en la exclusión como fuera de ella, es favorecido por la facilitación de leguminosas espinosas (Olneya tesota y Prosopis articulata).
Materiales y Métodos
Área de estudio
El área de estudio está localizada en la Bahía de la Paz (24.13(N, 110.44(W), límite sur del Desierto Sonorense, a 15 km de distancia de la ciudad de La Paz, Baja California Sur, México. Se sitúa en una extensa llanura aluvial, tradicionalmente bajo uso ganadero, caracterizada por suelos arenosos, profundos, con poca materia orgánica y escasa pendiente (<1.5%) (Valderrain-Algara et al., 2010). El estudio se realizó en la Reserva Biológica Dra. Laura Arriaga, perteneciente al Centro de Investigaciones Biológicas del Noroeste, que contiene 170 ha excluidas del uso ganadero desde 1984, y en un área equivalente del rancho adyacente (160 ha), donde persiste la presencia de ganado vacuno, con una densidad histórica aproximada de 0.15-0.25 cabezas/ha.
La vegetación es matorral sarcocaule conformado principalmente por una combinación de: (i) leguminosas leñosas como Prosopis articulata, Olneya tesota, Caesalpinia californica Standl., C. placida Brandegee, (ii) plantas de troncos engrosados como Fouquieria diguetii (Tiegh.) I.M. Johnst., Cyrtocarpa edulis (Brandegee) Standl., Bursera microphylla A. Gray, B. odorata Brandegee, B. epinnata Engl., Jatropha cinerea (Ortega) Müll. Arg., J. cuneata Wiggins & Rollins, (iii) plantas suculentas como P. pringlei, Stenocereus gummosus (Engelm.) A.C. Gibson & K.E. Horak, S. thurberi (Engelm.) Buxb., Ferocactus peninsulae (F.A.C. Weber) Britton & Rose, Lophocereus schottii (Engelm.) Britton & Rose, Cylindropuntia cholla (F.A.C. Weber) F.M. Knuth) y (iv) arbustos como Adelia brandegeei V.W. Steinm., Castela peninsularis Rose y Larrea tridentata (DC.) Coville (Perea et al., 2005).
La zona se caracteriza por un clima muy árido, seco, cálido extremoso con lluvias de verano, BW(h´)h(e)w de acuerdo con Köppen modificado por García (2004). Según datos de la Estación Climatológica de La Paz (24.135(N, 110.336(W; CONAGUA, 2023), tiene una precipitación anual promedio de 160 mm, temperatura mensual promedio de 24.1 (C, y temperatura mínima y máxima promedio de 17.5 y 30.7 (C, respectivamente.
Especie de estudio
El cardón (P. pringlei) es una cactácea columnar arborescente de gran tamaño, que puede alcanzar 15-20 m de altura y más de 1 m de diámetro (Turner et al., 1995). Es una especie endémica del Desierto Sonorense distribuida a lo largo de la península de Baja California (22.52(N - 31.31(N) y en una franja disyunta en la contracosta de Sonora (27.49(N - 30.13(N) (Félix-Burruel, 2012). Al igual que ocurre en otros cactus columnares, la gestión espacial y temporal de un gran volúmen de biomasa (nutrientes y agua) y su papel en la creación de hábitat (León de la Luz et al., 2002; Valiente-Banuet et al., 2002; Garvie, 2003, 2006; Wolf y Martínez-del Río, 2003; Drezner, 2014; Montesinos-Navarro et al., 2019) otorgan a esta especie una gran importancia funcional en su área de distribución.
Las poblaciones analizadas por Medel-Narváez et al. (2006) a lo largo del rango geográfico de distribución de la especie muestran distribuciones desiguales de sus clases de tamaño. Este patrón, común en otras especies de cactus columnares (Godínez-Álvarez et al., 2003), está asociado a pulsos temporales de altas tasas de reclutamiento separados por largos periodos de muy bajo reclutamiento (Bullock et al., 2005; Félix-Burruel et al., 2021). La tasa estimada de reclutamiento anual promedio del cardón varía entre 0.0054 ind/ind.año en el centro de la península (Bullock et al., 2005) y 0.0090 ind/ind.año a lo largo de la península (Félix-Burruel, 2012). La madurez reproductiva del cardón está relacionada con la altura del individuo. En la Reserva Biológica Dra. Laura Arriaga, solo 4% de los individuos maduros reproductivamente tiene una altura inferior a 3 m (Seminario-Peña, 2020).
Muestreo
En septiembre y octubre de 2011 delimitamos cuatro parcelas de aproximadamente 300 × 1000 m (30 ha) cada una: dos de ellas en el área con ganado y dos en el área de exclusión (Fig. 1). En cada parcela distribuimos aleatoriamente 10 transectos de 50 × 2 m y en cada uno de estos registramos y medimos diámetro y altura de todos los individuos de cardón y de las leguminosas O. tesota y P. articulata. Los individuos menores a 2 m se midieron con un flexómetro y los superiores a 2 m de altura, mediante un estadal Mound City (Crain Enterprises, Inc., Mound City, EUA).
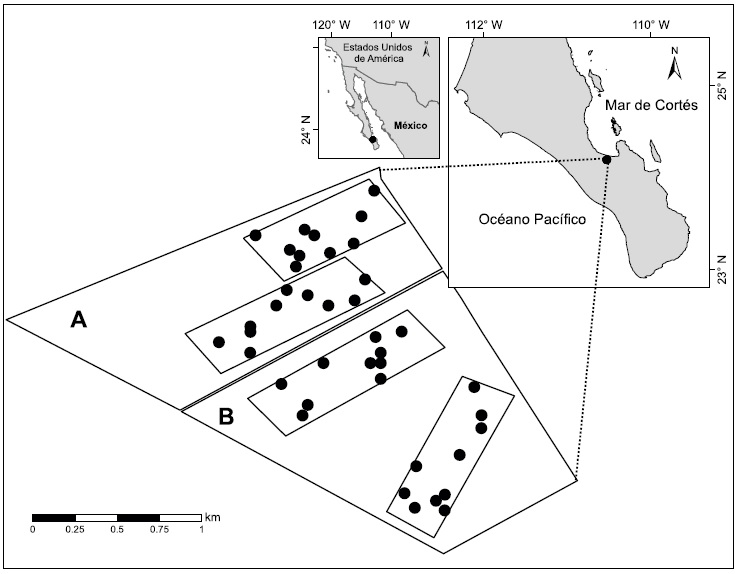
Figura 1: Localización geográfica del área de estudio en la Bahía de La Paz, límite sur del Desierto Sonorense en la península de Baja California Sur, México. Se muestra el área con presencia de ganado (A) y de exclusión (B), las dos parcelas de 30 ha delimitadas en cada área y los transectos de 50 × 2 m realizados (10 transectos por parcela).
Con base en los estudios demográficos del cardón realizados por Medel-Narváez (2003) y Seminario-Peña (2020) en esta área, clasificamos los cardones en cuatro etapas de vida: plántula (P) <51 cm, juvenil (J) 51-175 cm, adulto pre-reproductivo (PR) 176-300 cm) y adulto reproductivo (R) >300 cm. Los individuos adultos reproductivos los desagregamos en cuatro categorías de tamaño: R1=301-400 cm; R2=401-500 cm; R3=501-600 cm y R4=> 600 cm).
Análisis
Delgado-Fernández et al. (2016) dataron mediante análisis de 14C la edad de cardones de distintos tamaños en poblaciones a lo largo de la península de Baja California y estimaron que la relación entre altura (h) y edad (t) de los individuos se expresaba en el modelo h=a/b (e bt - 1), donde el valor estimado de los parámetros fue a=0.016 y b=0.0424. Según este modelo, la altura estimada de un cardón, reclutado en el año que se establece la exclusión, que tendría 27 años sería igual a 81 cm, con un intervalo de confianza asociado al 95% de 33.9-147.7 cm (Delgado-Fernández et al., 2016). A partir de esta estimación y para efecto práctico del presente estudio consideramos que los cardones de altura igual o inferior a 82 cm se reclutaron posteriormente a 1984, fecha del establecimiento de la exclusión.
En un primer paso, analizamos si la distribución de la densidad de cardones por transecto es diferente entre las dos condiciones (ganado/exclusión). Una vez constatado que la densidad no se ajusta a los supuestos de normalidad en el área con ganado (ganado: W=0.92294, P=0.1129; exclusión: W=0.75513, P<0.001), efectuamos una comparación entre la distribución de las densidades observadas en ambos escenarios utilizando el test de suma de rangos de Wilcoxon.
En segundo lugar, examinamos (i) si el patrón observado en la distribución de densidad de individuos por transecto puede ser atribuido a variación aleatoria o, por el contrario, exhibe un patrón significativo, y (ii) en qué medida los individuos reclutados después de la fecha de exclusión son responsables del patrón del conjunto poblacional observado en cada escenario. Para responder al primer inciso, comparamos mediante una prueba G de bondad de ajuste si la densidad observada de la población total en cada escenario se diferenciaba de la esperada de una distribución Poisson, que representa el modelo de distribución aleatoria. Para responder al inciso (ii) realizamos el mismo análisis, una prueba G de bondad de ajuste, a los individuos de cardón establecidos antes de la exclusión (i.e., superiores a 81 cm) en cada escenario y registramos si había cambio respecto al patrón de su población completa. Si existe diferencia sería atribuible al efecto del reclutamiento posterior a la exclusión.
Nuestro segundo análisis se dirigió a explorar si existe diferencia en la distribución de los individuos en las etapas de vida de plántula (P), juvenil (J), adulto pre-reproductivo (PR) y adultos reproductivos (R1, R2, R3 y R4) entre la población de la exclusión y la de presencia de ganado. Para ello comparamos las distribuciones observadas de individuos en etapas de vida en cada escenario mediante un análisis de bondad de ajuste de ( 2, considerando en un primer análisis los adultos reproductivos una sola categoría y, posteriormente, separados en cuatro categorías.
Finalmente, para explorar si existe relación de facilitación por leguminosas leñosas espinosas en el reclutamiento del cardón, analizamos si la presencia de las especies O. tesota y P. articulata muestra relación con el reclutamiento de individuos de cardón ocurrido tras la exclusión (i.e., individuos <82 cm), bajo su copa o inmediatamente próximos, y si la intensidad de esta relación es diferente entre ambos tratamientos. Para ello dividimos los transectos de 50 × 2 m en cinco tramos consecutivos de 10 × 2 m y registramos en cada uno de ellos la presencia de cardones menores a 82 cm de altura y de individuos de O. tesota y P. articulata con altura superior a 2 m y diámetro del tallo a la altura del pecho superior a 2.5 cm.
Obtuvimos 100 tramos en cada condición; sin embargo, para evitar posible autocorrelación espacial seleccionamos una muestra aleatoria de 50 tramos alternos (separados por 10 m) en cada condición. Consideramos la presencia del cardón en los tramos como variable respuesta binaria y la presencia de las leguminosas nodriza como variable explicativa igualmente binaria. Analizamos mediante una regresión logística, por medio de modelos lineales generalizados con error binomial, si la presencia de leguminosas nodriza influye en la probabilidad de la presencia de reclutamiento del cardón en cada uno de los tratamientos. El cociente entre la devianza residual y los grados de libertad fue inferior a uno en ambos casos, por lo que constatamos la ausencia de sobredispersión. Todos los análisis se realizaron en R v. 4.2.3 (R Core Team, 2023).
Resultados
En el área con ganado registramos un total de 37 individuos de cardón frente a 63 individuos encontrados en el área de exclusión. La densidad promedio de cardones en el área de ganado, 0.019 ind/m2( 0.013 D.E., fue menor que en la exclusión, 0.032 ind/m2( 0.033 D.E., pero no se encontraron diferencias significativas en el test de suma de rangos de Wilcoxon (W=237, P=0.308). Las dos distribuciones presentaron la misma mediana (0.02 ind/m2), pero mientras la distribución en el área de ganado fue relativamente simétrica (coeficiente de asimetría de Pearson=-0.34), en el área de exclusión presentó sesgo hacia la derecha (coeficiente de asimetría de Pearson=1.06; Fig. 2A).
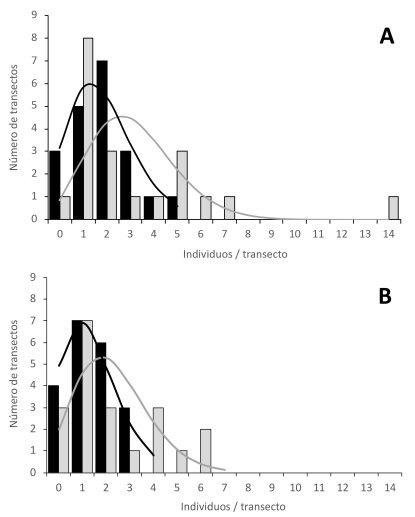
Figura 2: Distribución de la densidad de individuos de cardones (Pachycereus pringlei Britton & Rose) registrada en 20 transectos (50 ´ 2 m) en dos condiciones de uso: con presencia (negro) y exclusión de ganado (gris). A. considerando todos los individuos registrados; B. considerando solo individuos establecidos antes del inicio de la exclusión (>27 años, >81 cm). Las barras muestran la densidad observada y las líneas corresponden a los valores esperados de la distribución Poisson.
En el área de ganado la distribución de la densidad presentó un carácter aleatorio (coeficiente de dispersión=0.93), no distinto significativamente del patrón aleatorio expresado por una distribución Poisson (G=1.078, g.l.=3, P=0.782) (Fig. 2A). En cambio, la densidad de cardones en el área de exclusión mostró una distribución agrupada (coeficiente de dispersión=3.38), significativamente diferente de una distribución Poisson (G=13.377; g.l.=4; P=0.0096) (Fig. 2A).
Los cardones reclutados posteriormente a la fecha de la exclusión (<82 cm de altura) en el área con ganado fueron nueve (24% del total poblacional) y en el de exclusión 17 (27%). Si excluímos del análisis estos cardones inferiores a 82 cm, la densidad en el escenario de ganado siguió mostrando una distribución aleatoria (G=1.022, g.l.=2, P=0.600); sin embargo, en el escenario de exclusión el patrón de la densidad se volvió aleatorio (G=7.216, g.l.=4, P=0.125) (Fig. 2B). El agrupamiento espacial de los cardones observado en la exclusión parece estar relacionado con el patrón espacial del reclutamiento.
La distribución de individuos por etapas de vida (plántulas, juveniles, adultos pre-reproductivos y adultos reproductivos) no mostró diferencias estadísticamente significativas entre el área con ganado y el de exclusión (( 2=4.97, g.l.=3, P=0.174). En ambas condiciones, la distribución de individuos se concentra mayoritariamente en la etapa de adultos reproductivos (ganado 57%; exclusión 60%). El análisis de la distribución de individuos en siete categorías, desagregando la etapa de adultos reproductivos en cuatro categorías a intervalos de 1 m de altura, tampoco mostró diferencias significativas entre exclusión y área con ganado (( 2=10.96, g.l.=6, P=0.090) (Fig. 3).
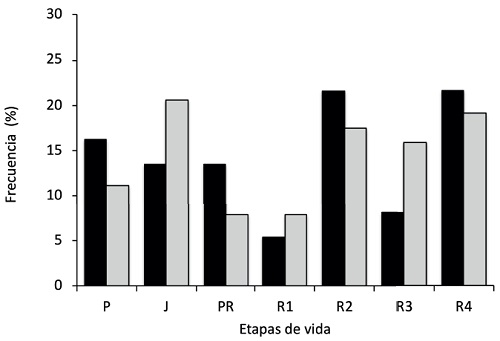
Figura 3: Distribución de individuos de cardón (Pachycereus pringlei Britton & Rose) en etapas de vida en dos condiciones de uso distintos en la Bahía de La Paz, Baja California Sur, México. Barras negras: presencia de ganado, barras grises: exclusión de ganado por 27 años. Basada en 20 transectos de 50 × 2 m en cada escenario. Etapas de vida: P: plántulas, <50 cm; J: juveniles, 51-175 cm; PR: pre-reproductivos, 176-300 cm; Ri: reproductivos, (R1: 301-400 cm, R2: 401-500 cm, R3: 501-600 m, R4: >600 cm).
En los 50 tramos alternos seleccionados en cada condición encontramos leguminosas en 14 tramos del área con ganado y en 11 tramos de la exclusión. En el área con ganado cuatro de los 14 tramos con leguminosas (29%) presentaron reclutamiento, frente a tan solo uno de los 11 tramos con leguminosas (9%) en la exclusión. La presencia de las especies nodriza, O. tesota y P. articulata, en la exclusión no parece tener relación con la localización de nuevos reclutas de cardón (Z=0.335, P=0.738), pero sí está positivamente relacionada con el reclutamiento en el área con ganado (Z=-2.044, P=0.041) (Cuadro 1). La probabilidad de reclutamiento (p) estimada en el escenario de ganado para un tramo con presencia de individuos de leguminosa nodriza, (p=1/(1+1/e ((0 )), fue p=0.29 ( 0.06 E.E. y en ausencia de nodriza, (p=1/(1+1/e ((0+(1 )), p=0.06 ( 0.03 E.E.
Cuadro 1: Resultado del análisis de regresión logística a la relación entre presencia de leguminosas leñosas nodrizas (Olneya tesota A. Gray y Prosopis articulata S. Watson) y presencia de cardones (Pachycereus pringlei Britton & Rose) menores a 0.82 cm de altura, reclutados durante los 27 años previos al muestreo, en un área con ganado y en otra con exclusión de ganado en la Bahía de La Paz, Baja California Sur, México. Z=estadístico de la prueba de Wald; P=probabilidad de Z en una distribución normal; *= significante, P<0.05; n.s.=no significante.
| Tratamiento | Variable | Coeficiente | Error estándar | Z | P |
| Ganado | Intercepto (β0) | -0.9163 | 0.5916 | -1.549 | 0.1214 |
| Nodriza (β1) | -1.9169 | 0.9378 | -2.044 | 0.0409* | |
| Exclusión | Intercepto (β0) | -2.3026 | 1.0486 | -2.196 | 0.0281* |
| Nodriza (β1) | 0.3857 | 1.1528 | 0.335 | 0.7380n.s. |
Discusión
En este estudio nos propusimos analizar si la exclusión de ganado durante 27 años afectó a la tasa de reclutamiento del cardón (Pachycereus pringlei) en un matorral sarcocaule de la Bahía de la Paz, en el límite sur del Desierto Sonorense en la península de Baja California. Nuestros resultados mostraron que (i) no existen diferencias en la proporción de individuos reclutados (<82 cm de altura) dentro y fuera de la exclusión, y que (ii) existe asociación positiva entre la presencia de leguminosas nodrizas (O. tesota y P. articulata) y la probabilidad de reclutamiento de cardón en el área con ganado, pero no en el área de exclusión. El primer resultado contradice nuestra primera hipótesis, que la exclusión de ganado aumenta la tasa de reclutamiento, y el segundo solo respalda parcialmente nuestra segunda hipótesis, que las leguminosas nodrizas facilitan el reclutamiento del cardón en ambos escenarios.
La distribución de frecuencias entre las etapas de vida observada en el área dentro y fuera de la exclusión fue similar. En ambas condiciones predominan los individuos adultos reproductivos con un destacable componente de adultos de gran tamaño. Por lo tanto, podemos considerar que la capacidad reproductiva, en términos relativos, en ambos tratamientos es similar. Sin embargo, sobre esta similitud demográfica de partida esperábamos que la exclusión de ganado estuviera asociada a una mayor tasa de reclutamiento.
La falta de diferencia en la tasa de reclutamiento dentro y fuera de la exclusión podría estar asociada a que el reclutamiento del cardón, al igual que otras cactáceas columnares (Félix-Burruel et al., 2021), presenta pulsos de reclutamiento muy espaciados en el tiempo (Bullock et al., 2005; Medel-Narváez et al., 2006; Félix-Burruel, 2012). Esta discontinuidad temporal en el reclutamiento de la especie podría ser potenciada por las características locales del suelo. En zonas de escasa pendiente y textura de suelo fina, como es nuestra área de estudio, se ha observado que el carácter episódico del reclutamiento de las cactáceas columnares se intensifica respecto a lugares rocosos (Steenbergh y Lowe, 1983; Orum et al., 2016; Conver et al., 2017).
La escala de variabilidad multidecenal en el reclutamiento del cardón se combinaría con escalas temporales menores de variabilidad en otros atributos demográficas que afectarían al reclutamiento en los años no propicios. Por ejemplo, en 2002, en el área de exclusión de nuestro estudio, 76% de 140 individuos adultos revisados produjeron flores (Medel-Narváez, 2003). En cambio, en 2019 solo lo hizo 32% de 468 individuos adultos revisados (Seminario-Peña, 2020).
Fleming et al. (2001) encontraron una alta variación interanual en la intensidad de floración en los cardonales de Sonora, basándose en el monitoreo del mismo grupo de individuos en diferentes años. Esta variación llegó a ser hasta tres veces superior en un año con respecto a otro. Desde esta perspectiva, quizá la variabilidad a diferentes escalas temporales (anuales, decenales y multidecenales) del reclutamiento y de diversas variables demográficas del cardón no haya permitido que en tan solo tres décadas de exclusión se vean reflejadas potenciales diferencias en las tasas de reclutamiento del cardón entre el área con ganado y sin ganado.
La elevada tasa de supervivencia de plántulas observada en un experimento protegido de predación sugiere que la predación puede ser un factor determinante en la supervivencia de plántulas durante sus primeros meses de vida (Sosa y Fleming, 1999). Sin embargo, la respuesta de las poblaciones de roedores a la exclusión de ganado en regiones áridas es compleja y no necesariamente implicaría un aumento de su abundancia total (Mathis et al., 2006; Rickart et al., 2013). La presencia de ganado parece estar asociada a un cambio en las abundancias relativas de las especies de roedores (Jones y Longland, 1999; Tabeni y Ojeda, 2005; Miguel et al., 2017). En este sentido, la exclusión pudiera haber modificado la abundancia relativa de las especies de roedores respecto al área con ganado y afectar, positiva o negativamente, a la tasa de predación sobre semillas y plántulas del cardón. Sin embargo, la contigüidad espacial existente entre los dos tratamientos y la moderada extensión espacial de la exclusión respecto a la región con uso de ganado circundante pueden propiciar que los potenciales efectos de la exclusión sobre la abundancia relativa de las especies de polinizadores, consumidores o dispersores de semillas sean parcialmente neutralizados por la escala de movimiento de estas especies.
Respecto a nuestra segunda hipótesis, el resultado observado de asociación positiva entre leguminosas nodrizas (O. tesota y P. articulata) y reclutamiento en el área con ganado, pero no en la exclusión, puede estar relacionado con que el mejoramiento nutricional del suelo por las leguminosas constituya un efecto diferencial de facilitación en escenarios perturbados (Carrillo-García et al., 1999). Aunque la proporción de suelo desnudo es semejante en ambos escenarios (ganado 36% ( 2.4 E.E.; exclusión 38% ( 2.5 E.E.), parece sensato asumir que la perturbación del mismo por la actividad del ganado debe ser mayor en el rancho.
Otra posibilidad, no excluyente, es que la asociación positiva observada en la zona de ganado se deba a la función de barrera disuasoria que el ramaje espinoso de ambas leguminosas ofrecería al movimiento del ganado en el espacio próximo a estos árboles. La reducción en la frecuencia del paso del ganado disminiuría la probabilidad de que plántulas y juveniles próximos a estos árboles, incluso aunque no estuviesen estrictamente bajo la copa de los mismos, sean pisoteadas o consumidos por el ganado. La protección defensiva de O. tesota sobre las plantas que crecen bajo su cobertura se ha asociado a la posible reducción en la predación de semillas, plántulas y juveniles de saguaro (Niering et al., 1963; McAuliffe, 1984; Búrquez y Quintana, 1994). Sin embargo, en nuestro estudio la función defensiva no sería tanto frente a la predación de semillas o plántulas por roedores u otros herbívoros, que debería reflejarse también en el área de la exclusión, sino frente al potencial consumo o destrucción por pisoteo directo del ganado. Este mecanismo de protección de plántulas y juveniles frente al pisoteo del ganado por parte de O. tesota se ha sugerido como uno de los factores por los que esta leguminosa es una especie nodriza tan importante para los grandes cactus en el Desierto Sonorense (Tewksbury y Petrovich, 1994).
La asociación espacial del reclutamiento con las nodrizas en el escenario de ganado, pero no en la exclusión, puede estar relacionado con el patrón aleatorio de la distribución de la población en el primero y agregado en el segundo escenario. La destrucción de plántulas en el área con ganado, por su mayor frecuencia de paso en las áreas sin leguminosas espinosas, podría modificar la tendencia a la agregación de los nuevos reclutas del cardón respecto a los cardones ya establecidos (patrón observado en el área de exclusión) y desplazar la distribución espacial de los individuos hacia un mayor grado de aleatoriedad (patrón observado en el área con ganado). Esta destrucción diferencial de plántulas en el área con ganado debería reflejarse en una menor tasa de reclutamiento en el área de ganado, como planteamos en nuestra hipótesis de trabajo. Sin embargo, nuestros resultados no lo muestran.
Lo anterior es aún más llamativo si consideramos que los efectos de disturbios, como sequías y ganado, son más intensos en poblaciones de cactus columnares presentes en suelos llanos y no rocosos (Winkler et al., 2018). En suelos rocosos existen más sitios seguros donde semillas y plántulas encuentran cierto mejoramiento del estrés hídrico o protección frente al disturbio del ganado. Si nuestra interpretación de la presencia de estas leguminosas como disuasión al movimiento del ganado es correcta, estas especies representarían una variante de espacio seguro para el reclutamiento y su presencia podría significar un atenuante al posible efecto destructivo del ganado sobre el reclutamiento de los cardones y quizá también de otras especies de plantas.
En ambientes bajo pastoreo libre, este tipo de especies de leguminosas leñosas espinosas podrían crear “islas de protección” a su alrededor, a semejanza de las reconocidas islas de recursos (García-Moya y McKell, 1970), que faciliten el reclutamiento y mantenimiento espacial y temporal de poblaciones de numerosas especies de plantas. Esta relación se suma a la reconocida funcionalidad ecológica que tienen las leguminosas leñosas en regiones áridas, como estructuradoras de hábitat y gestoras de recursos (Suzán et al., 1996; Golubov et al., 2001), reforzando su papel como especies ecológicamente claves en la dinámica y conservación de estos ecosistemas.

