











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El cultivo de huertos familiares ofrece numerosos beneficios, como la disponibilidad de alimentos frescos y de alta calidad, la generación de oportunidades laborales, y la reducción de costos en la economía familiar, además del aprovechamiento de residuos vegetales (Giaconi y Escaff, 2001). Las hortalizas, que forman parte de la agricultura de traspatio, pueden ser de cultivos anuales o perennes según la especie. Estas plantas son esenciales en la dieta humana, ya que, consumidas crudas o cocidas, son ricas en vitaminas y minerales, poseen propiedades antioxidantes y antiinflamatorias, contribuyendo significativamente a la salud (Vilaplana, 2004). Aunque se consumen partes específicas de estos vegetales, los residuos restantes suelen ser desechados. Lo que ocasiona un exceso de residuos vegetales que podrían ser reciclados favorablemente para fines alimenticios, pecuarios y agrícolas, aprovechando su potencial (Burezq y Davidson, 2021).
En México, las hortalizas más destacadas por su consumo son cebolla, jitomate, zanahoria, rábano, brócoli, betabel y repollo. Las tres últimas son el foco de la presente investigación debido a su presencia en los traspatios locales. Estas hortalizas se utilizan como acompañantes en comidas típicas de la dieta mexicana, como tostadas, garnachas, tacos fritos y empanadas, siendo de uso y consumo diario en la cocina típica nacional. Asimismo, el brócoli, por su aporte de fibra y vitaminas, es un componente esencial en la alimentación (Baker, 2019).
Es importante tener en cuenta que, estos cultivos tienen altos requerimientos de nutrimentos esenciales como nitrógeno, fosforo y potasio (Sierra, 2013), que son extraídos del suelo o suministrados mediante actividades como la fertilización. Sin embargo, la disponibilidad de estos nutrimentos dependerá de su presencia en la solución del suelo. Para determinar la presencia de los nutrimentos esenciales, previo al establecimiento de un cultivo es de suma importancia realizar un análisis de suelo, lo que permite evaluar la disponibilidad de nutrimentos, pH, conductividad eléctrica. Esto ayuda a determinar si el suelo es apto para cierto cultivo y prever posibles deficiencias durante su desarrollo (Osorio, 2012; Orozco-Corral, Valverde, Martínez, Chávez y Benavides, 2016).
El análisis de nutrimentos suele realizarse una vez cada dos años en toda la superficie del terreno a ser cultivado (Schweizer-Lassaga, 2010). La confiabilidad de los resultados depende de la experiencia del operario y de los criterios de muestreo utilizados. Con el análisis se establece el plan de fertilización, que en la mayoría de los casos se realiza según un calendario (p. ej.: cada 15 o 30 días) y no según puntos críticos de alta demanda de nutrimentos relacionados con la fenología y el desgaste de nutrimentos en la rizósfera.
Actualmente, en el mercado agrícola existen equipos con sensores que determinan in situ el contenido de nutrimentos en la solución del suelo. Además, se han integrado herramientas de mecatrónica e inteligencia artificial, debido al alto rendimiento, precisión y rentabilidad que ofrecen (Bonilla, Dávila y Villa, 2021). Ante el problema del costo elevado de los análisis de suelos y la escasez de laboratorios en las comunidades agrícolas, como en el caso de Chiapas, el uso de equipos portátiles para la determinación de nutrimentos del suelo y otras variables químicas surge como una herramienta valiosa para el seguimiento de la nutrición de los cultivos desde la siembra hasta las etapas fenológicas importantes. La fenología de las plantas influye fuertemente en la concentración requerida de nutrimentos, a medida que la planta crece y pasa por etapas de su desarrollo la absorción de nutrimentos varía. Este proceso es crucial para entender las necesidades de la planta en diferentes momentos y para optimizar la fertilización y otras prácticas agronómicas (Escudero y Mediavilla, 2003). Sin embargo, es importante tener conocimiento sobre la fertilización de los cultivos y el monitoreo de los nutrimentos presentes en el suelo, ya que, estos son absorbidos por las raíces estimulando el crecimiento vegetal debido a su traslocación en las partes de la planta (Fernández, 2001).
Por otro lado, la producción de hortalizas, son fuentes significativas de diversos tipos de residuos, ya que, se desprecian ciertas partes de la planta para consumo, que pueden ser utilizadas como biofertilizantes o fuente de nutrimentos para alimentación humana y animal (Giaconi y Escaff, 2001; Moreno et al., 2012).
La generación de estos residuos no solo plantea desafíos en términos de gestión y disposición ambiental (Manchado y Villamizar, 2003) sino que también ofrece oportunidades para el desarrollo de estrategias de reutilización y reciclaje de nutrimentos que puedan convertir estos subproductos en recursos valiosos para otros procesos industriales, uso y aprovechamiento de las propiedades nutricionales que estos también ofrecen o para la generación de energía, Por lo que el estudio del contenido y bioacumulación de nutrimentos en estos tejidos resulta relevante (Corredor y Pérez, 2018; Aguiar, Estrella y Cabadiana, 2022). La adecuada gestión de estos residuos es crucial para minimizar su impacto ambiental, mejorar la eficiencia de los recursos y fomentar al sector agroindustrial (Cury, Aguas, Martínez, Olivero y Chams, 2017; Aguiar et al., 2022). Los residuos vegetales derivados de la producción de hortalizas representan una fuente rica de lignocelulosa y algunos nutrimentos como Zn, Fe y Mg, también pueden contener ingredientes bioactivos. El cultivo de betabel o el de crucíferas como el repollo y brócoli aportan un alto valor nutritivo y medicinal por su contenido de fibras, proteínas, vitaminas y carotenos (Jaramillo y Díaz, 2006; Pinto-Zapata, 2012). Por lo anterior, el objetivo de la presente investigación fue evaluar el comportamiento de macronutrimentos y propiedades químicas del suelo, relacionadas con la rizósfera de cultivos de importancia agrícola como el brócoli, repollo y betabel, al finalizar la etapa vegetativa. Además, analizar la acumulación de biomasa y la distribución de Fe y Mg en órganos vegetales considerados como residuos.
Materiales y Métodos
El experimento se estableció en el rancho “Capricho Divino”, ubicado en la comunidad de Plan De Mulumí, perteneciente al municipio de Suchiapa, Chiapas, con las coordenadas 16° 37’ 59” N y 93° 07’ 15” O a una altitud de 454 metros de altitud. Se utilizaron plántulas de repollo (Brassica oleracea var. Capitata), Betabel (Beta vulgaris) y brócoli (Brassica oleracea var. Itálica), que pertenecen al grupo de plantas que comúnmente se siembran en los traspatios.
Manejo agronómico
Se realizó una limpieza manual de la cobertura vegetal utilizando machete. Para la labranza, se emplearon coa y azadón. Se construyeron camellones de 20 m de largo, 0.4 m de ancho y 0.25 m de alto. El suministro de agua se llevó a cabo mediante riego por goteo. Para el manejo de arvenses, se instaló un acolchado plástico de color blanco. La siembra se realizó en horas de la mañana y se aplicó una lámina de riego de un litro por gotero. Se dejaron espacios vacíos para análisis posteriores del suelo. La incidencia de plagas se presentó solo en las crucíferas, principalmente lepidópteros, los cuales se controlaron con Spinetoram. La incidencia de enfermedades no fue agronómicamente significativa. No se realizó la aplicación de fertilizantes (condición de manejo local).
Análisis fisicoquímico del suelo
Para determinar el contenido de macronutrimentos del suelo antes del establecimiento de los cultivos, se marcaron 10 zonas de muestreo distribuidas en el área de cultivo en Zigzag. Se midieron los niveles de nitrógeno (N), fósforo (P), potasio (K), pH y conductividad eléctrica (CE) utilizando un equipo multiparamétrico de suelo modelo GEMHO-TY, China. La textura del suelo se determinó mediante el tacto, siguiendo la metodología reportada por Bautista y Oliver (2017), y se identificó como un suelo franco arenoso (Fr-A). Posteriormente, cuando los cultivos culminaron su etapa de crecimiento vegetativo BBCH v-39 (Meier, 2018), correspondiente a 60 días después del trasplante, se realizó un análisis del suelo en la rizósfera de los tres cultivos, a 1 cm de la base de la planta y en las zonas sin cultivar dentro del surco.
Determinación de contenido de biomasa
Se cuantificó el contenido y la distribución de la biomasa fresca en tallo, hoja y raíz utilizando una balanza analítica. Posteriormente, cada muestra fue secada en un horno de convección a 60 °C hasta obtener pesos constantes. Finalmente, se cuantificó la biomasa seca de los tejidos aéreos y subterráneos.
Evaluación nutrimental
Se siguió la metodología adaptada de Gutiérrez-Miceli et al. (2021). La digestión de las muestras (500 mg) se realizó con 9 mL de ácido nítrico, 2 mL de agua oxigenada al 30% y 1 mL de ácido clorhídrico a 180 °C, hasta lograr la digestión total del material vegetal. Después de la digestión, las muestras fueron filtradas con papel Whatman No. 4 y diluidas a un volumen final de 50 mL. La cuantificación de hierro y magnesio en hojas y raíz se realizó mediante espectroscopía de emisión atómica con plasma de acoplamiento inductivo (ICP-OES, Perkin Elmer, NexIon 300 D, Shelton, EE. UU.).
Análisis estadístico
Cada especie vegetal constituyeron los tres tratamientos, con cinco repeticiones cada uno. La unidad experimental fue una planta, distribuida en un diseño completamente al azar. Para la evaluación nutrimental se utilizaron tres repeticiones. Se realizó un análisis de varianza unidireccional (ANOVA) y la prueba de Tukey se utilizó para evaluar la diferencia de medias entre tratamientos (P ≤ 0.05). Los datos se analizaron utilizando el software Statgraphics Centurion versión 16 (Statgraphics Technologies, 2009).
Resultados y Discusión
En el Cuadro 1 se presentan los resultados de los análisis del suelo realizados antes del establecimiento de los cultivos y 60 días después del trasplante. No se observaron diferencias significativas en las propiedades fisicoquímicas del suelo entre ambos momentos, lo que indica que los nutrimentos se mantienen constantes en la solución del suelo hasta la interacción con las plantas. Los valores correspondientes al muestreo al día 60 se utilizaron como referencia para determinar el nivel de extracción de nutrimentos y los cambios en las propiedades químicas del suelo en la rizósfera.
Temperatura y pH en la rizósfera
Se observó una variación significativa en la temperatura del suelo en ausencia y presencia de especies vegetales, lo que sugiere un comportamiento diferencial en la transferencia de energía. La temperatura más alta del suelo sin cultivo alcanzó los 33.8 °C. Sin embargo, con la cobertura foliar de las crucíferas (repollo y brócoli), se observó una reducción de 3 °C en la temperatura de la rizósfera (Figura 1). Beta vulgaris, con un crecimiento menor y una menor cobertura, mostró una disminución de 1 °C en la temperatura del suelo. Estos hallazgos indican que la presencia y el tipo de cultivo influyen en la temperatura del suelo, lo que puede tener importantes implicaciones para el crecimiento y desarrollo de las plantas. La temperatura del suelo varía debido a factores climáticos, que dependen de la altitud del cultivo, así como del manejo agrícola y del suelo. La cobertura foliar actúa como una barrera natural que protege el suelo de la radiación solar directa, reduciendo así la temperatura del suelo y previniendo la pérdida excesiva de humedad por evaporación (Fischer y Lüdders, 1998; Scavo et al., 2022).
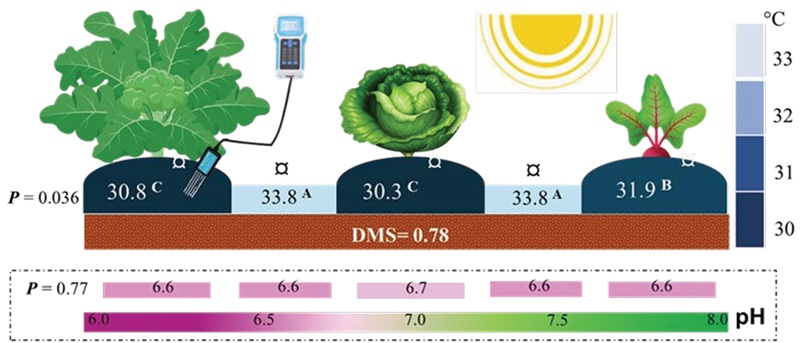
Los cultivos de brócoli y coliflor son considerados hortalizas de temporada fría, pero se cultivan en zonas cálidas al inicio de las lluvias. En nuestro estudio, la temperatura local supera los límites superiores adecuados para el crecimiento de estas especies; en el caso de las coles, el rango óptimo es de 26 a 30 °C, y para el betabel es de 20 a 34 °C (Kahn, Rebek y Damicone, 2009). Sin embargo, aunque las plantas se desarrollaron normalmente hasta la etapa vegetativa, no alcanzaron una producción adecuada en el caso de las coles. El betabel logró alcanzar su madurez fisiológica, aunque con un tamaño de tubérculo menor (5.1±1.1 cm).
El pH del suelo fue ligeramente ácido y no experimentó cambios notables con la presencia de los cultivos, salvo un ligero incremento de 0.1 en el caso de Brassica oleracea var. Capitata. Este aumento marginal podría atribuirse a diversos factores, como la liberación de ciertos compuestos por parte de las raíces de las plantas o la actividad microbiana en el suelo. La estabilidad del pH sugiere que las interacciones en la zona radicular, incluyendo el intercambio de nutrimentos y la liberación de exudados por parte de las plantas, no tienen un impacto significativo en este parámetro del suelo. Este hallazgo destaca la capacidad de los cultivos para mantener un equilibrio en el entorno del suelo, incluso mientras absorben nutrimentos y liberan diversas sustancias. Por otro lado, el pH y la temperatura del suelo juegan un papel crucial en el desarrollo radicular, así como en la eficiencia con la que las plantas absorben agua y nutrimentos (Boczulak, Hawkins y Roy, 2014). Variaciones en estos parámetros puede afectar directamente la tasa de crecimiento de las raíces y la absorción de elementos esenciales como potasio y nitrógeno, lo que impacta la salud general y el rendimiento de los cultivos (Sierra, 2013; Du et al., 2021).
Dinámica NPK en la rizósfera
El comportamiento de los nutrimentos en la zona radicular de cada cultivo se muestra en el Cuadro 2. Se observó una menor concentración de nitrógeno, seguido de fósforo y potasio, lo cual puede estar relacionado con el nivel de absorción de cada nutrimento. La conductividad eléctrica (CE) del suelo mostró una tendencia similar a la absorción de NPK en la zona radicular. En el cultivo de repollo se registró una CE de 167 µS cm-¹, en brócoli de 221.38 µS cm-¹ y en betabel de 309 µS cm-¹, indicando variaciones en la CE según el cultivo y el contenido de NPK. Esta propiedad del suelo engloba la presencia de factores como el contenido de agua, la presencia de arcilla, concentración de iones intercambiables los cuales están directamente relacionados con sus propiedades nutritivas (Cortés, Pérez, John y Camacho, 2013; Simón, Peralta, y Costa, 2013). De igual modo, indica la cantidad de sales presentes en el suelo y depende de múltiples factores, uno de ellos es la humedad, por lo que, la CE disminuye cuando el suelo está saturado de agua, debido a la dilución de las sales, y aumenta en condiciones de sequía, ya que las sales se concentran (Bosch, Costa, Cabria y Aparicio, 2012).
Cuadro 2: Contenido de NPK y conductividad eléctrica (CE) del suelo en la rizosfera de repollo, brócoli y betabel.
Table 2: NPK content and soil electrical conductivity (EC) in the rhizosphere of cabbage, broccoli and beet.
| Cultivo | N | P | K | CE |
| - - - - - - - - - - mg kg-1- - - - - - - - - - | µS cm-1 | |||
| Repollo | 11.5 a* | 16.4 b | 32.8 b | 167.4 b |
| Brócoli | 15.4 b | 21.5 b | 43.9 b | 221.4 b |
| Betabel | 22.8 a | 32.0 a | 64.6 a | 309.0 a |
| DMS† | 6.86 | 9.84 | 19.6 | 73.48 |
| Suelo | 92 | 140.34 | 253.2 | 1276 |
† Diferencia mínima significativa. *Diferente literal entre columnas indica diferencias significativas entre tratamientos (Tukey, P < 0.05).
† Least Significant Difference. *Different literal between columns indicates significant differences between treatments (Tukey, P < 0.05).
En cuanto a la absorción de nutrimentos en la rizósfera, el cultivo de betabel alcanzó su madurez fisiológica BBCH 49, momento en el cual el tubérculo alcanzó el tamaño de cosecha, utilizando el 79% de los nutrimentos presentes en el suelo. Este hallazgo sugiere que no se requiere un aporte adicional de nutrimentos al suelo bajo las condiciones del estudio. La disponibilidad de los nutrimentos en la solución del suelo parece favorecer su absorción, posiblemente influenciada por la clase textural del suelo y su pH (6.6±0.1). Se ha señalado que la mayoría de los nutrimentos no presentan limitaciones en su disponibilidad en un rango de pH de 6.0 a 7.0 (Cremona y Enríquez, 2020).
Por otro lado, tanto el brócoli como el repollo exhibieron un comportamiento similar, alcanzando la etapa BBCH 39, donde el tallo principal alcanza la altura típica para la variedad, con una absorción de NPK del 86, 91 y 86%, respectivamente. Estos resultados sugieren que, para las características nutricionales del suelo estudiado, por agotamiento de NPK, el punto crítico de aporte de nutrimentos para lograr una buena cosecha corresponde a la etapa BBCH 39. Se recomienda que el suministro de nutrimentos mediante la aplicación de materia orgánica se realice a la par con la preparación del suelo y en las primeras prácticas de labranza, como el desyerbe o el aporque, dado que su descomposición es lenta y, de este modo, estará disponible para el cultivo en la etapa mencionada. En el caso de los fertilizantes de síntesis química, su efectividad dependerá de su solubilidad (Navarro-García, 2023).
Contenido y distribución de biomasa
La biomasa vegetal es un factor cuantitativo que determina el crecimiento, la productividad y la nutrición del cultivo, a mayor acumulación se relaciona con mayor absorción de nutrimentos. La capacidad de absorción de nutrimentos de una especie vegetal puede ser determinada tomando como indicador la biomasa acumulada (Sifuentes et al., 2015). En el cultivo de brócoli, la biomasa fue mayor en comparación con el repollo y el betabel. El brócoli mostró un 52% más de biomasa fresca y un 65% más de biomasa seca que el repollo, y un 70% más de biomasa fresca y un 75% más de biomasa seca que el betabel (Figura 2). Esta información es de interés al momento de utilizar material vegetal de residuo con fines de nutrición para calcular la cantidad de nutrimentos en este caso Fe y Mg, que estaríamos aportando. En investigaciones recientes, la inclusión de cultivos con alta producción de biomasa y acumulación de nutrimentos esenciales, mejoran significativamente la capacidad de mejorar la fertilidad del suelo y almacenar carbono mejorando su estructura y fertilidad (Burezq y Davidson, 2021).
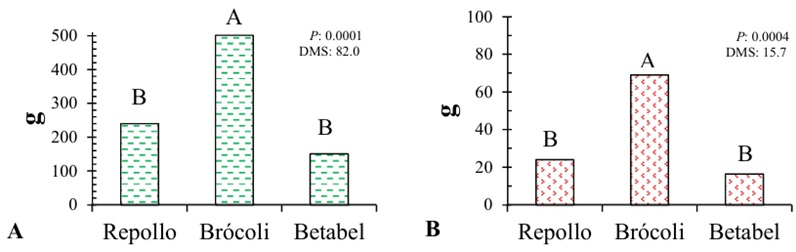
El comportamiento de los cultivos y la distribución de sus reservas entre los órganos vegetales evaluados se presenta en el Cuadro 3. Se observaron diferencias estadísticas significativas entre las tres especies. El cultivo de brócoli mostró la mayor cantidad de biomasa acumulada en tallo y hojas, seguido por el repollo, mientras que el betabel tuvo la menor producción de biomasa en estos órganos vegetales. En cuanto a la biomasa seca, el brócoli también presentó la mayor acumulación, mientras que el repollo y el betabel mostraron comportamientos similares entre sí. La acumulación de reservas en la raíz fue significativamente mayor en el betabel, seguido por el brócoli, con el repollo presentando la menor proporción de biomasa en la raíz.
Table 3: Percentage distribution of fresh and dry dough.
| Cultivo | Fresco* | Seco | ||||||||
| Total | Tallo | Hoja | Raíz | Total | Tallo | Hoja | Raíz | |||
| - - - - - - - g - - - - - - - | - - - - - - - g - - - - - - - | |||||||||
| Repollo | 90.0 a | 8.3 b | 86.7 a | 5.3 b | 10.0 b | 8.0 b | 79.7 a | 12.3 b | ||
| Brócoli | 86.3 b | 13.6 a | 78.3 b | 7.3 b | 13.7 b | 13.3 b | 69.7 b | 17.0 b | ||
| Betabel | 89.3 a | 0.0 c | 45.7 b | 54.3 a | 10.7 ab | 0.0 b | 43.7 c | 56.3 a | ||
| DMS† | 2.7 | 1.8 | 9.5 | 9.3 | 3.4 | 3.6 | 8.8 | 9.6 | ||
† Diferencia mínima significativa. *Diferente literal entre columnas indica diferencias significativas entre tratamientos (Tukey, P < 0.05).
† Least Significant Difference. *Different literal between columns indicates significant differences between treatments (Tukey, P < 0.05).
En el betabel, se observa una proporción similar en la distribución de reservas. En estado fresco, el tubérculo representa el 55% de las reservas, mientras que en estado seco esta cifra asciende al 57%. El porcentaje restante corresponde a la roseta foliar (hojas). En contraste, en las crucíferas como el repollo, el 87% de las reservas se encuentran en las hojas, cifra que se mantiene en un 80% en estado seco. El brócoli presenta valores similares, con una diferencia de 10% por debajo, distribuidos entre el tallo y la raíz. Durante esta etapa fenológica, el brócoli aumenta las reservas en el tallo y la raíz en comparación con el repollo. La distribución de la biomasa vegetal es dependiente de la especie y su genotipo, y está influenciada por el crecimiento y la tasa de distribución, los cuales son afectados por el ambiente y la disponibilidad de nutrimentos. La productividad de las plantas y la biomasa incrementan cuando hay nutrientes disponibles en el suelo (Barbosa et al., 2014). La materia seca almacenada en los órganos vegetales representa los excedentes de asimilados, que serán utilizados para la formación de tejidos en etapas posteriores (Barrientos, del Castillo y García, 2015).
Contenido y distribución de Fe y Mg
El contenido de Fe y Mg en los órganos vegetales de las especies evaluadas se muestra en el Cuadro 4. Los resultados obtenidos revelan que el comportamiento entre las crucíferas no presenta diferencias estadísticas, por lo que el contenido de minerales es similar entre ellas. Sin embargo, las raíces presentaron un mayor contenido de hierro (Fe) y las hojas un mayor contenido de magnesio (Mg) en el cultivo de repollo. En cuanto al brócoli, se observó un contenido ligeramente mayor de Mg en las raíces, sin diferencias en el contenido de Fe entre los dos tejidos vegetales evaluados.
Cuadro 4 Contenido de hierro (Fe) y magnesio (Mg) en hoja y raíz Table 4. Iron (Fe) and magnesium (Mg) content in leaf and root.
Table 4: Color of squash fruit peel (Cucurbita pepo L.) var. ‘Grey Zucchini’s.
| Cultivo | Hoja | Raíz | ||
| Fe | Mg | Fe | Mg | |
| - - - - - - - - - mg de metal por g de muestra - - - - - - - | ||||
| Repollo | 0.069 a | 1.97 b | 2.2 a | 2.40 a |
| Brócoli | 0.171 ab | 1.77 b | 1.6 ab | 2.96 a |
| Betabel | 0.214 a | 12.81 b | 0.13 b | 2.70 a |
| DMS | 0.108 | 2.51 | 2.27 | 0.65 |
| P | 0.0158 | 0.0001 | 0.1103 | 0.095 |
Letras distintas indican diferencias estadísticas significativas (P ≤ 0.05).
Different letters indicate statistically significant differences (P ≤ 0.05).
En las raíces de las coles se encontró un elevado contenido de Fe, un tejido que generalmente es desechado, lo que incrementa la problemática de acumulación de residuos vegetales. El Fe es un microelemento esencial para el desarrollo fisiológico en plantas. Además, las hojas y raíces con alto contenido de Mg y Fe son esenciales en la dieta de los rumiantes en donde se puede aprovechar. El Mg es crucial para funciones enzimáticas y el Fe es vital para el transporte de oxígeno en los animales (Mata, 2015; Rakhi y Abdullah, 2022).
El cultivo de betabel presentó una elevada concentración de Mg en sus hojas, superior al encontrado en las crucíferas, cabe destacar que las hojas del cultivo de betabel generalmente son consideradas residuos vegetales. El Mg es un elemento esencial en las plantas porque participa en la fotosíntesis y la partición de carbohidratos.
Un estudio reciente de Yue et al. (2024) evaluó el contenido de minerales (Mg, Fe y Zinc) en hojas cosechables de 159 variedades de col (Brassica oleracea L.). Los resultados mostraron que el contenido de Fe fue mayor en variedades de repollo verde, con un contenido de Mg de 7.6 mg g-1 y Fe de 0.6 mg g-1, superiores a los encontrados para la variedad de repollo verde de este estudio, que fueron de Mg 1.97 mg g-1 y Fe 0.07 mg g-1. Es importante resaltar que estos valores más bajos se deben a que el análisis se realizó en hojas maduras que se desechan del repollo al momento de la venta. Estos resultados indican que hay una traslocación de estos nutrientes hacia hojas o tejidos jóvenes, aunque no en su totalidad, ya que un porcentaje permanece en el tejido maduro.
Hermans Johnson, Strasser y Verbruggen (2004) realizaron un análisis sobre la deficiencia de magnesio en plantas de Beta vulgaris (remolacha), encontraron que el Mg afecta la fotosíntesis y la partición de carbohidratos. Este mineral se desplaza en el suelo mediante un proceso de difusión por transporte pasivo, la translocación y asimilación de este elemento por la planta implica transporte vía floema desde hojas viejas hasta hojas jóvenes, interviene en la síntesis de hormonas, proteínas y lípidos durante la asimilación de nitrógeno por lo que es un elemento sumamente importante, necesario para la formación de clorofila (Farhat et al., 2016; López, 20161). La presencia de altos niveles de Mg en las hojas del betabel sugiere que este cultivo podría ser una fuente valiosa de Mg no solo para la nutrición vegetal, sino también para la alimentación animal, especialmente en sistemas de producción que buscan reducir residuos vegetales.
La concentración de Fe fue mayor en las hojas del betabel y menor en la raíz, contrario a lo observado en las crucíferas que concentran mayor cantidad de Fe en la raíz. El hierro es un mineral requerido en bajas concentraciones y está relacionado con la actividad fotosintética de la planta (Baran, 2021). La distribución de Fe en general tiende a ser más acumulado en raíces en las crucíferas, mientras que, en el betabel muestra mayor concentración en las hojas (Farhat et al., 2016).
En los residuos vegetales del betabel, las hojas contienen un elevado contenido de magnesio (Mg). Este mineral es un macronutrimento requerido en la dieta diaria de muchos organismos, en el caso de los rumiantes, junto con otros elementos, forma parte de los componentes del esqueleto y dientes, está involucrado en el funcionamiento celular del animal y participa en reacciones enzimáticas, lo cual beneficia al rumen. Del mismo modo, el Fe es un microelemento necesario diariamente en el rumen y en el transporte de oxígeno (Cseh, 2015). En los cultivos de crucíferas, hay un contenido importante de estos minerales en las raíces, las cuales son desechadas durante el consumo humano. Estos residuos pueden ser aprovechados para la alimentación animal.
Conclusiones
El análisis del suelo previo al cultivo reveló importantes respuestas sobre la variación de la temperatura, pH, CE y absorción de NPK. Se observó una variación significativa en la temperatura del suelo, con una reducción notable en la zona radicular con la presencia de cultivos. Aunque el pH del suelo permaneció estable, la CE mostró una tendencia similar a la absorción de nutrimentos, destacando la importancia de estas mediciones para evaluar la productividad del suelo. El nivel de desgaste de nutrimentos en la rizósfera permitió determinar el punto crítico para el aporte de nutrimentos. El análisis del contenido de Fe y Mg en los residuos vegetales de repollo, brócoli y betabel reveló diferencias significativas en la distribución de estos minerales entre los diferentes órganos vegetales. Las raíces de las crucíferas presentan altos niveles de Fe, mientras que las hojas de betabel contienen niveles elevados de Mg. Estos hallazgos resaltan la importancia de aprovechar los residuos vegetales ricos en minerales para beneficio de la nutrición animal, especialmente en sistemas de producción de rumiantes. Futuras investigaciones deberían centrarse en optimizar el uso de estos residuos para maximizar sus beneficios nutricionales y minimizar su impacto ambiental.

