











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
INTRODUCTION
Reproduction is a significant event in the life cycle of living beings and, along with growth and feeding, can ensure the persistence of organisms in the environments they inhabit (Nikolsky 1963). Studies describing fish reproduction in species involved in commercial fisheries are important. These studies provide basic information on the biology and population characteristics of fish, particularly concerning their reproduction. Therefore, these studies are essential inputs to evaluate and manage actions for fishery management (West 1990, Cailliet et al. 1996, Froese 2004).
The species of the Sciaenidae family are found in temperate and tropical regions worldwide, including the Indo-Pacific region, the Caribbean Sea, Amazon waters, the Mediterranean Sea, and the Atlantic and Pacific oceans. This is one of the most abundant families, with 69 genera and 289 species (Parenti 2020). In Mexico, it is distributed along the entire coast of the Pacific Ocean, including the Gulf of California (Robertson and Allen 2015), and is also observed on the Atlantic side, in the Gulf of Mexico (Nakamura 1981).
Sciaenids are known as croakers or drums due to the sounds they make. These sounds are used to locate aggregations; nevertheless, they can be heard more frequently during the reproductive period, especially while males attract females (Lagardère and Mariani 2006, Robertson and Allen 2015). Most species in this group are carnivorous and consume benthic macroinvertebrates, a variety of small fish, crustaceans, burrowing worms, and occasionally organic matter (FAO 1995, Robertson and Allen 2015). They are common in local markets because they are caught by fishers in the region (Robertson and Allen 2015).
Umbrina xanti is a species of the Sciaenidae family distributed from Mexico (Baja California) to northern Peru. It is a demersal fish that inhabits shallow coastal areas with sandy bottoms and can enter coastal lagoons and estuaries (Robertson and Allen 2015). It is a commercially important species in artisanal fisheries in the Mexican Pacific (Sandoval-Ramírez et al. 2020); in particular, its capture is frequent in the central Pacific (Espino-Barr et al. 2003, 2004; Gómez-Vanega et al. 2021), including in the region of the present study (Rojo-Vázquez et al. 2001, 2008). Despite this, and its particular ecological importance, little is known about certain aspects of its life history that would clarify population dynamics for fishery management.
With the reproduction of fish, the first life stage of an entire generation begins, and this introduction of new organisms is one of the main factors that influence the dynamics of fish populations (Csirke 1989). Because there is no information on the reproductive aspects of the Polla Drum U. xanti, the aim of this study was to estimate reproductive variables such as the gonadosomatic index, sex ratio, length at sexual maturity, and microscopic description of the gonads. The results of this study will expand the existing biological information on the species, which can be used by authorities to develop and design a fisheries management plan on the southern coast of Jalisco, Mexico.
MATERIALS AND METHODS
Fish were collected monthly for 5 consecutive days from January 1998 to December 2008. The organisms analyzed came from the commercial catch of artisanal fisheries operating in Bahía de Navidad (19°10′27″ to 19°13′18″ N, 104°41′54″ to 104°49′07″ W) located in the south of Jalisco, Mexico (Fig. 1). The fishers used nylon monofilament gillnets with different mesh sizes (7.20, 8.89, 10.16, and 11.43 cm) operating at the bottom for approximately 12 effective fishing hours at night. The total length (TL ± 1 mm) and total weight (TW ± 0.1 g) were obtained for each U. xanti individual. Sex and maturity were determined by direct observation of the gonads (Palazón-Fernández 2007), and their weight was recorded (GW ± 0.01 g). The gonads were fixed in 10% neutral formalin (buffered with dibasic and monobasic sodium phosphate) for subsequent histological analysis.
Figure 1 Study area: Bahía de Navidad, Jalisco, Mexico. The dots represent the sites where monthly sampling was conducted.
A 0.5-cm transverse section was taken from the middle portion of the gonads to describe the internal structure. Histological sections of ovaries and testes were taken for each stage of development. For the histological process, samples were dehydrated in a series of ethyl alcohol solutions in increasing concentrations, from 50% up to the absolute concentration, embedded in paraplast, cutting sections of 3-5 μm, and stained with hematoxylin-eosin dyes (Lucano-Ramírez et al. 2001a). Gonads were observed and analyzed using a microscope (Primo Star HD, Carl Zeiss, Oberkochen, Germany) equipped with an integrated camera (Axiocam MRC5, Carl Zeiss), using AxioVision v. 4.8.2 software (Carl Zeiss). The different stages of oocyte development were classified according to Yamamoto and Yamazaki (1961), Lucano-Ramírez et al. (2001a), and Brown-Peterson et al. (2011). The structure of the internal cells of the testis was described according to the methods of Lucano-Ramírez et al. (2001b) and Uribe et al. (2018).
To identify the reproductive season, we used 3 complementary methods; the first was based on the monthly calculation of the gonadosomatic index (GSI) (Sánchez-Cárdenas et al. 2007):
For the second, we applied the morpho-chromatic scale of 4 stages of gonadal development (immature, maturing, mature, and spawned or regressing) monthly, according to the criteria of Palazón-Fernández (2007). For the third method, we analyzed monthly variations in the average oocyte diameter, assuming that a larger average diameter corresponds to a greater degree of ovarian development; for this, we measured 30 oocytes from each stage of maturation from each ovary (largest diameter and smallest diameter of the oocytes that presented a nucleus) (West 1990, Lucano-Ramírez et al. 2014).
The weight-length relationship was calculated using the potential model equation proposed by Ricker (1975):
where TW is the total weight, TL is the total length, and a and b are parameters of the fitted curve. The value of b of the weight-length relationship was used to calculate the relative condition factor (CF) for each organism (Froese 2006):
where TW is total weight, TL is the total length and b is the parameter of the fitted curve. The sex ratio was obtained as the ratio of the number of males to the number of females and was calculated for the total sample, for each month, and by length class. The chi-square (χ2) test with Yates’ correction was used to assess whether the sex ratio differed from the expected value of 1:1 (Zar 2010).
The mean length at sexual maturity (L 50) was considered the length at which 50% of the sampled individuals had reached sexual maturity. To obtain this, the logistic model was fitted to the frequency of mature fish (in 2-cm intervals of TL). The L 50 value for each sex was obtained with the estimated parameters in each case. The percentage of mature individuals at a given length (P TL ) was estimated:
where a and b are the parameters of the logistic model. The L 50 was estimated from the result of a divided by b (Echeverría 1987). This model was fitted for each sex with a nonlinear method based on the Levenberg-Maquard algorithm using STATISTICA v. 7.1 (Statsoft 2006). To evaluate possible differences in maturation ogives between sexes, a residuals or coincident curves analysis was performed (Chen et al. 1992).
The existence of significant differences between mean monthly values of GSI, CF, and oocyte diameter was verified using an analysis of variance (ANOVA), and the combined monthly variation between different variables was verified using the nonparametric Spearman’s rank correlation test (r s). For all tests, STATISTICA v. 7.1 (Statsoft 2006) was used, with a significance level of 0.05.
RESULTS
A total of 1,202 U. xanti organisms were collected, with TL values ranging from 17.9 to 42.3 cm. Of these, 80% were between 28 and 32 cm TL, and the remaining percentage had TL values <28 cm and >32 cm. The TL values of females ranged from 17.9 to 42.3 cm (mean: 29.73 ± 0.11 cm) and their TW values ranged from 67.0 to 837.7 g (mean: 293.54 ± 3.36 g). In males, TL values ranged from 23.6 to 42.1 cm (mean: 29.70 ± 0.09 cm) and their TW values ranged from 143.0 to 807.0 g (mean: 289.29 ± 3.11 g) (Fig. 2).
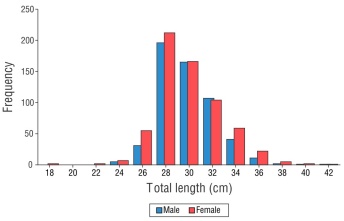
Figure 2 Frequency distribution of the total length of females and males of Umbrina xanti captured in the central Mexican Pacific.
Sex ratio
The sex of 1,197 organisms was identified, of which 637 were females (53%) and 560 were males (47%); the sex ratio for the organisms analyzed was 1:0.88 (F:M). This differed significantly (χ 2 = 4.95, P = 0.03) from the expected 1:1 ratio. In turn, the analysis of the number of females and males per length class showed no significant differences were observed with respect to the expected 1:1 ratio in any class (Fig. 1).
In the analysis of the monthly sex ratio, there were significant differences in March (χ 2 = 4.49, P = 0.03), April (χ2 = 7.91, P = 0.01), and October (χ 2 = 7.26, P = 0.01); in these 3 months the number of females exceeded that of males.
Reproductive season
The monthly distribution of gonadal maturity percentages indicated that the immature stage exhibited relatively low percentages in females (January, April, and September-December) and in males (January-June and November-December). The maturing stage and the spawning or regressing stage occurred in all months in both sexes, with percentages above 30% in several of them. The mature stage in females occurred from January to June, with high percentages of 51% in February and 55% in May. In males, this stage showed a similar trend to that of females; the highest percentages occurred from January to April, with 34% in January and 51% in March (Fig. 3).
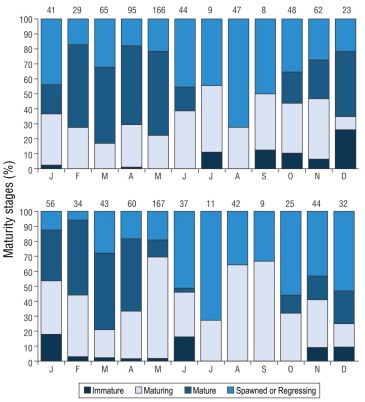
Figure 3 Monthly percentages of gonadal maturity stages of female (top) and male (bottom) Umbrina xanti in the central Mexican Pacific. Numbers at the top indicate sample size.
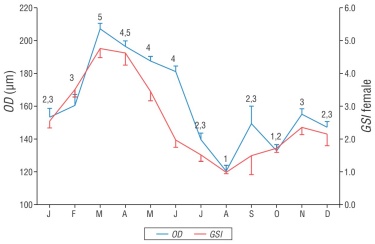
The GSI of females showed significant variations throughout the year (F 11, 625 = 11.98, P < 0.01), with the highest values in March (4.75 ± 0.27) and April (4.62 ± 0.36). Conversely, the lowest value was obtained in August (0.98 ± 0.04). The average value of the GSI in females for the entire study was 2.8. In addition, the variation in the GSI in males was significant throughout the year (F 11, 548 = 16.92, P < 0.01), with the maximum value found in April (1.63 ± 0.14) and the minimum values in June (0.34 ± 0.04) and August (0.25 ± 0.03). The overall mean value of the GSI in males was 0.8. The SNK multiple contrast test identified groups with high values in March and April in both sexes (C and 3-4), and low values from June to December (A-B and 1-2), also in both sexes. The Spearman’s correlation coefficient indicated a high correlation between the monthly averages of the GSI of males and females (r s = 0.867, P < 0.01, n = 12); this suggests that the values in both sexes showed a similar seasonal trend (Fig. 4).
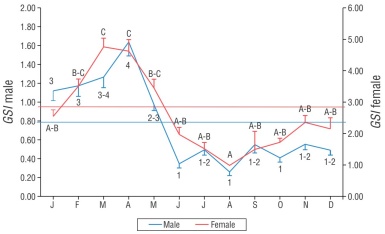
Figure 4 Monthly variation in the gonadosomatic index (GSI) of female and male Umbrina xanti captured in the central Mexican Pacific. Bars indicate the standard error. Transverse lines represent the mean. Numbers (males) and letters (females) indicate significant differences.
The monthly variation in U. xanti oocyte diameter was significant (F 11, 7510 = 46.86, P < 0.01), with the largest diameters occurring from March to June (groups 4 and 5). The smallest diameters (groups 1 and 2) were recorded from July to October and from December to January. A correlation was determined between oocyte diameter and the GSI of females (r s = 0.853, P < 0.01, n = 12) (Fig. 5). The monthly variations in the percentages of gonadal maturity, the GSI, and oocyte diameter suggest that the species reproduces from January to May.
Relative condition factor
The relative condition factor (CF) varied significantly throughout the year for females (F 11, 625 = 4.56, P < 0.01) and males (F 11, 548 = 4.48, P < 0.01). In females, the lowest values occurred in July, August, and September; in males, they occurred in January, August, and September. The SNK test identified 2 groups for both sexes (A and B, 1 and 2), with overlaps occurring in several months. A similar seasonal trend was observed in both sexes, as the Spearman’s correlation coefficient indicated a correlation between the monthly averages of the CF of females and males (r s = 0.60, P < 0.04, n = 12) (Fig. 6).
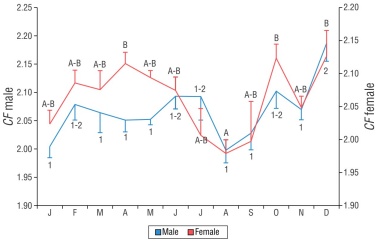
Figure 6 Monthly variation in the relative condition factor (CF) of female and male Umbrina xanti captured in the central Mexican Pacific. Bars indicate the standard error. Numbers (males) and letters (females) indicate significant differences.
The correlation analysis between the variables GSI and CF indicated no significance in females (r s = 0.524, P = 0.80, n = 12). However, months with high CF values also showed high GSI values, which could indicate that there is no high cost or energy investment in females at the time of reproduction. In males, the correlation was also not significant (r s = -0.287, P = 0.37, n = 12). However, in March, April, and May, the CF showed low values and, in the same months, the GSI showed the highest values. Therefore, we can infer that, in males, gonadal maturity could affect the physiological state of the organisms.
Microscopic description of the gonads
The ovaries are covered by the ovarian tunic, which is made of muscular tissue. Inside the ovary, we observed groups of oocytes at different stages of development, delimited by lamellae (Fig. 7). We identified 7 stages of oocyte development: 1) oocytes in primary growth, with an average diameter of 65.00 ± 2.10 µm, characterized by having little cytoplasm and a large nucleus with nucleoli. 2) Oocytes with vitelline vesicles or cortical alveoli with an average diameter of 135.15 ± 5.48 µm. Oil droplets are distinguished in the cytoplasm, and with these, the formation of the yolk begins. 3) Oocytes in primary vitellogenesis with an average diameter of 202.88 ± 7.51 µm. Yolk globules are observed in the cytoplasm. 4) Oocytes in secondary vitellogenesis with an average diameter of 307.38 ± 8.25 µm. A large number of yolk globules were present throughout the cytoplasm. 5) Oocytes in tertiary vitellogenesis with an average diameter of 319.58 ± 12.23 µm. The yolk globules began to fuse, and the nucleus was found migrating toward the animal pole. 6) Mature oocytes with a diameter of 335.64 ± 9.27 µm. The yolk was completely fused, and the nucleus was not observed. 7) Hydrated oocytes with an average diameter of 367.03 ± 19.62 µm. The simultaneous presence of oocytes in different development stages in the mature ovary indicates that U. xanti has an asynchronous-type development (Fig. 6).
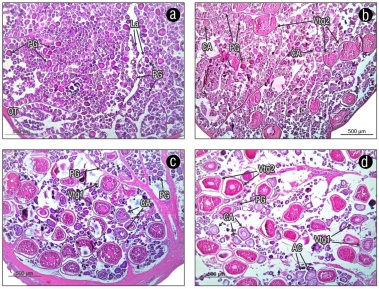
Figure 7 Cross sections of ovaries at different stages of maturity of Umbrina xanti: immature (a), maturing (b), mature (c), and spawned or regressing (d). Hematoxylin-eosin staining. Scale: 500 µm. OT = ovarian tunic; PG = primary growth oocyte; CA = oocyte in cortical alveolus; Vtg1 = oocyte in primary vitellogenesis; Vtg2 = oocyte in secondary vitellogenesis; La = lamella.
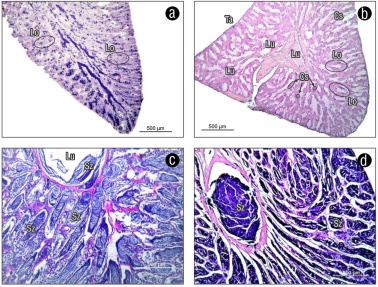
The germ cells within the testis were found organized into lobules and, within these, clusters of cells were observed forming cysts; this is where the entire process of spermatogenesis took place. Once the sperm were formed, they concentrated in the lumen of the lobules and subsequently accumulated in the receptor duct to be expelled. This type of sex cell arrangement suggests an unrestricted lobular type development (Fig. 8).
Average length at sexual maturity (L 50 )
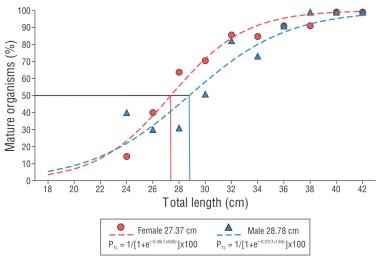
The smallest female with mature gonads had a TL value of 22.6 cm, and the smallest male with the same characteristic had a TL value of 24.5 cm. According to the results obtained with the calculation of the average length at maturity, 50% of females reached sexual maturity at 27.37 cm TL and males at 28.78 cm TL. The overall average length for the females analyzed was 29.48 cm, which corresponds to 68% of the females having characteristics of gonadal maturity. For males, the overall average length was 29.27 cm, which corresponds to 66% of the males sampled having mature gonads. Thus, we can deduce that more than 50% of the organisms obtained from commercial captures would have had at least one reproductive event. For each of the 4 nets used in this study, the average length was greater than that of sexual maturity; it is worth mentioning that the 7.20-cm net caught only 28% of the organisms below the sexual maturity length. The residuals or coincident curves analysis indicated that the maturation curves between sexes showed no significant difference (F = 2.054, P = 0.14); therefore, the average length at maturity for combined sexes was 27.98 cm TL with a slope value of 0.31 (Fig. 9).
DISCUSSION
The study of reproductive aspects in commercially important species, and even in those without this status, contributes useful basic elements to manage commercial fishing appropriately. This information can be used to estimate the potential impacts of environmental and anthropogenic effects on the population dynamics of U. xanti.
Total length distribution
The average length and the intervals with the highest frequency of U. xanti organisms collected in the present study were similar to those found by Espino-Barr et al. (2003) in Colima (30.16 cm) and Espino-Barr et al. (2004) in Jalisco (29.62 cm), where the highest TL values of the organisms captured were between 25.35 and 32.25 cm and 27.16 and 31.46 cm, respectively. The similarity between values obtained in previous studies and those obtained in this study could be due to the fish coming from commercial fisheries, which use gillnets with similar characteristics, and the study areas being close together.
Sex ratio
The ratio of females to males is an important element that allows us to understand reproductive dynamics in a population. Most species that reproduce sexually have a sex ratio close to 1:1; this maximizes reproductive success and genetic diversity and tends to increase resilience to changes and disturbances. However, when there are greater numbers of either sex, these tend to balance out over time through natural selection (Fisher 1930, Nikolsky 1963). In the present study, females were more abundant than males; this result is similar to those found in other studies conducted with sciaenids, such as Sciaena umbra in Spain, where Grau et al. (2009) also found that females dominated over males with a ratio of 1.3:1.0 (F:M). The same was observed for Bairdiella ronchus in the Colombian Caribbean, with a notable abundance of females (1.7:1.0; F:M) (Torres-Castro et al. 1999). In addition, Fennessy (2000) described this same pattern in sciaenids off the coast of South Africa. This difference between sexes, according to Sadovy and Shapiro (1987), occurs in gonochoric species due to differences in growth and longevity, spatial segregation, sexual survival rate, and spawning migration. Furthermore, Silberschneider and Gray (2008) noted that in some species a higher survival rate has been recorded in females.
Reproductive season
One of the simplest, most widely used, and fastest methods to assess the level of gonadal development is the use of macroscopic scales (West 1990, Brown-Peterson et al. 2011). The scale proposed by Palazón-Fernández (2007) to characterize ovarian and testicular development consists of 4 maturation stages, which were identified by specific characteristics such as coloration, size, and the presence or absence of fluids and oocytes. This scale was used in the present study and, conjointly with the GSI, allowed us to identify the reproductive period of U. xanti. However, other authors have used different maturation scales that adapt more closely to the characteristics they observed in a particular species; for example, Prista et al. (2014) used a 5-stage scale to characterize female and male maturation in the croaker Argyrosomus regius.
Microscopic studies help confirm the degree of gonad maturity and define the type of oocyte development and arrangement of sperm cells in the testicle. At the microscopic level, the stage of testicle maturation can be distinguished based on structures and characteristics, such as the thickening of the tunica albuginea, the presence or absence of sperm, the amount of interstitial tissue, and the differentiation of the receptor duct; these vary depending on the degree of maturity (Grier et al. 2009). The testes of U. xanti males were heavier when mature because they are ready to release sperm. This development stage shows well-defined branches of the lobes, revealing the interstitial tissue, which is arranged radially and extends to the tunica albuginea (Grier 1981). Sperm accumulate in the ducts toward the receptor duct and cluster in it; these characteristics are specific to lobular testicular development. The species of sciaenids that exhibit these characteristics and the same developmental pattern are Micropogonias ondulatus (Barbieri et al. 1994), Roncador stearnsii (Miller et al. 2009), Umbrina roncador (Miller et al. 2009), and Larimichthys polyactis (Kyu-Lim et al. 2010).
The mature ovary of U. xanti shows 7 stages of oocyte development simultaneously. The increase in oocyte size is very clear from the beginning of vitellogenesis; the oocyte grows gradually due to the accumulation of yolk in the cytoplasm. Grau et al. (2009) classified oocyte development in S. umbra into 7 stages based on the diameter shown in each of the oocyte stages. The fact that various stages of oocyte development occurred simultaneously in mature U. xanti ovaries could confirm that they develop asynchronously. Consequently, oocytes are expected to be released in multiple spawnings. Another study on S. umbra off the coast of Turkey recorded 12 stages of oocyte development in the ovary; the authors assumed that oocyte development was asynchronous and considered this species to be iteroparous and gonochoric with multiple spawning events (Engin and Seyhan 2009). Different species of the Sciaenidae family, such as M. ondulatus (Baribieri et al. 1994), Sciaenops ocellatus (Wilson and Nieland 1994), Micropogonias furnieri (Vizziano et al. 2005), R. stearnsii (Miller et al. 2009), L. polyactis (Kyu-Lim et al. 2010), Menticirrhus americanus (Clardy et al. 2014), and Macrodon ancylodon (Dos Santos-Cardoso et al. 2018), show this same type of asynchronous development. Species that have oocytes with asynchronous development usually have partial spawning or multiple spawning, with a relatively broad reproductive season or with several reproductive events per year (González and Oyarzún 2002, Dos Santos-Cardoso et al. 2018).
The GSI is frequently used to identify reproductive periods (Brewer et al. 2007). In this study, both sexes of U. xanti showed maximum GSI values in spring. According to Mendo and Samame (1988), GSI values increase during the reproductive season and decrease after spawning. In the study region, coastal upwelling occurs from February to May (Ambriz-Arreola et al. 2012); this could support the increased reproductive activity of U. xanti, since during this period the larvae could benefit from increased primary productivity due to these coastal upwellings. In addition, Cárdenas (2012) mentions that this family reproduces mainly in warm temperatures, when conditions are more favorable for egg development and larval growth; therefore, several species of sciaenids have greater reproductive activity in spring and summer. This is also observed in Argyrosomus japonicus, which has a reproductive period between March and June in Taiwan (Ueng et al. 2007), and in A. regius, which reproduces between March and August off the coast of Spain (Jiménez et al. 2007). In turn, Vizziano et al. (2005) recorded the reproductive period from December to March for M. furnieri off the coast of Brazil. Meanwhile, for Cynoscion nebulosus in the Gulf of Mexico, spawning occurs from May to July (Brown-Peterson et al. 2002) and for S. umbra from March to June in the Mediterranean Sea (Chakroun-Marzouk and Ktari 2003).
Relative condition factor
The relative condition factor (CF) is a variable that reflects the body condition of fish and can be related to the quantity, quality, and availability of food in the environment where each species thrives (Froese 2006). High CF values indicate that organisms are in good or better physiological condition. In Bahía Navidad, Mexico, U. xanti females had peak CF values in April, October, and December, and males in February, June-July, October, and December. High CF values are usually associated with periods of increased or improved feeding.
In the study region, coastal upwellings were reported to increase productivity in these areas from February to May (Ambriz-Arreola et al. 2012), which could explain the increased CF during these months. The increased CF observed at the end of the year may be the effect of an intermediate climatic condition, since it is the final stretch of the rainy season and a transition period between tropical-subtropical and temperate conditions, where no single environmental condition clearly dominates (Ambriz-Arreola et al. 2012).
The highest CF values were observed at the beginning and end of the year (more pronounced in females), and increases in GSI were also recorded during this same period. The relationship between GSI and CF was not very clear in U. xanti. In females, reproduction does not appear to affect body condition; however, a negative trend was observed in males, although not significant. In some species, as reported by Wootton (1990), the energy investment of females results from the accumulation of yolk during oogenesis, though this is not the case in U. xanti. However, it has also been mentioned that energy expenditure for both sexes is due to the requirements of individuals, such as survival, growth, or reproduction, in relation to the environment in which they develop. In the case of reproduction, the investment could be associated with the increase in gonad size relative to the weight of the organism, which indicates that during this period the gonads are at their peak maturity. In some species, a decrease in CF values indicates a decrease in energy stored in the form of body lipids and proteins; these substances help meet energy requirements during reproduction (González and Oyarzún 2002).
Length at sexual maturity
In U. xanti, the average length of maturation did not differ between females and males. Similar to the findings in this study, on the Brazilian coast, Dos Santos-Cardoso et al. (2018) observed that both sexes of M. ancylodon mature at 21.3 cm, and for M. furnieri, maturation occurred at 11 cm (Vizziano et al. 2005); the authors suppose that this is possible if both sexes perform the same migrations and feed in similar locations; therefore, they reach sexual maturity at the same time. Other studies with sciaenids indicate that females reach sexual maturity at a greater length than males; in S. umbra, females matured at 23.4 cm and males at 22.1 cm (Chakroun-Marzouk and Ktari 2003). For the same species, Grau et al. (2009) reported that females reached sexual maturity at 29.9 cm and males at 25.4 cm. In turn, for B. ronchus in the Colombian Caribbean, females had an average length at sexual maturity of 15.8 cm and males of 15.4 cm (Torres-Castro et al. 1999). In most sciaenids, females reproduce at a greater length than males (Cárdenas 2012); this could be because females, in order to begin reproduction, need a greater amount of food to grow first and subsequently produce the number of eggs necessary to ensure an increase in the number of individuals in the population (Kunz 2004).
In U. xanti, the average capture length (29.7 cm) was slightly greater than the mean length at sexual maturity (28.1 cm). This suggests that slightly more than half of the captured organisms had already had at least one reproductive event. The average capture length was greater than the maturation length, which is a desirable situation in species subject to commercial exploitation according to Espino-Barr et al. (2008). In any case, future studies are needed to continue analyzing the population dynamics of species, mainly and especially those that are subject to continuous commercial capture.
CONCLUSIONS
The 3 methods used to identify the reproductive period of U. xanti indicated that both females and males had a prolonged reproductive period, which lasted from January to May. Umbrina xanti is a gonochoric species; microscopically, the germline cells of the testis display an unrestricted lobular organization, and oocyte development in the ovary is asynchronous. The average maturation length suggests that 62% of females and 61% of males had already had at least one reproductive event. The present study provides basic information on the reproduction of U. xanti, a species of commercial interest in the central Mexican Pacific. This information can be used to estimate the potential impacts of environmental and anthropogenic effects on the population dynamics of U. xanti.

