











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
INTRODUCTION
Marine macroalgae are of notable economic importance, serving as staples in human diets, substrates in biofuel production, biostimulants in agriculture, and reservoirs of high-value metabolites (Santos et al. 2023a, 2023b, 2024). Green macroalgae of the genus Ulva (Chlorophyta), formally described by Linnaeus in 1753, encompass a range of edible algae belonging to the Ulvaceae family. Among the most recognized species are the sea lettuces, which are widely distributed along coastlines and commonly observed in rocky intertidal shores in urban areas. Ulva species exhibit short life cycles characterized by high biomass followed by losses in biomass density (Calheiros et al. 2021).
Ulva species show high growth rates and a wide tolerance to abiotic factors (Osuna-Ruiz et al. 2019), which allows them to perform optimally in monocultures, in vitro cultures, and integrated multi-trophic aquaculture (IMTA) (Obando et al. 2022, Pitta et al. 2022). In IMTA, Ulva species are used as biofilters to assimilate dissolved nutrients (C, N, and P) from the effluents of fish and shellfish, consequently reducing water treatment costs and ensuring the production of high-value products (Silva et al. 2015, Nardelli et al. 2019, Oliveira Soares et al. 2022).
Considering the potential for the aquacultural production of Ulva biomass and the quest to develop innovative biotechnological applications to enhance the economic value of these species, several studies have emphasized that Ulva species are natural sources of highly valuable metabolites, including carotenoids (El-Baky et al. 2008, 2009; Chakraborty and Paulraj 2010; Eismann et al. 2020). The most abundant species observed in the state of Rio de Janeiro, Brazil, are the foliaceus Ulva fasciata Delile, Ulva lactuca Linneaus (sea lettuce), and the tubular Ulva flexuosa Wulfen (Calheiros et al. 2021, Carneiro et al. 2022). Recently, U. fasciata was recognized as a variant of U. lactuca (Guiry and Guiry 2024). Notably, in the capital city of Rio de Janeiro, these Ulva species have been cultivated in IMTA startups and sold as gastronomic and functional foods (Reis et al. 2017, Derner 2018, Pitta et al. 2022). Recently, the nutritional quality of Ulva species produced via commercial cultivation systems has been analyzed, confirming the potential of these systems to be environmentally sustainable and profitable income sources (Calheiros et al. 2021, Roleda et al. 2021, Pitta et al. 2022). Although research on the carotenoid content of Ulva species has progressed in Brazil, highlighting the biotechnological potential of these macroalgae (Raymundo et al. 2004, Sousa et al. 2008, Melo et al. 2021), additional studies are needed.
Carotenoids are terpenoids with C40 and conjugated double bonds that are commonly used in foods as natural colorants, antioxidants, and nutritional supplements. In the cosmetic and pharmaceutical industries, carotenoids have been utilized as nutraceuticals and antioxidants (Irwandi et al. 2011, Gateau et al. 2017, Maoka 2020). Carotenoids have photosynthetic and photoprotective properties, and their composition in thylakoid membranes is linked to chlorophylls and other factors that affect photosynthesis (Demmig-Adams and Adams 1996, Demmig-Adams 1998, Esteban et al. 2015). The carotenoids identified in Ulva species are synthesized by the methylerythritol phosphate (MEP) pathway from isoprenyl units in chloroplasts (He et al. 2018). It is worth noting that the following are common carotenoids reported in Ulva species: β-carotene, lutein, violaxanthin, antheraxanthin, zeaxanthin, and neoxanthin (El-Baky et al. 2009, Eismann et al. 2020).
The degradation of pigments, including that of carotenoids, is due to oxidation reactions caused by exposure to oxidant factors, including heat, light, acids, and oxygen. Carotenoid losses in stored dry biomass have created a bottleneck in the commercialization of Ulva species (Goldman et al. 1983, Woodall et al. 1997, Pérez-Galvez and Mínguez-Mosquera 2001). Carotenoid conservation in macroalgae biomass depends on different factors and varies among organisms; microalgae and vegetables can conserve carotenoids for periods of time ranging from 10 days to 3 months (Sugumaran et al. 2022). For instance, Nannochloropsis salina was successfully stored at 3 temperatures (5 °C, 20 °C, or 40 °C) for up to 8 weeks (Safafar et al. 2017), while Spinacia oleracea L. samples were stored in polypropylene bags at 2 °C or 10 °C (Bergquist et al. 2006). Furthermore, the effects of storage time (14 days or 35 days), storage conditions (vacuum-packed or non-vacuum packed), storage temperature (-20 °C, 4 °C, or 20 °C), and preservation method (spray-drying or freeze-drying) on the stability of lipids and carotenoids in fresh microalgal paste of Phaeodactylum tricornutum have been analyzed (Ryckebosch et al. 2011). Although different drying methods affect the chemical profiles of Ulva species (Uribe et al. 2018), it is not known how these methods affect carotenoid conservation.
Previous studies of Ulva species have evaluated various factors, such as the sustainability of biomass sources, storage, and carotenoid yield, and focused on commercial demand in the food and pharmaceutical sectors. In this study, we analyzed and compared the carotenoid and chlorophyll profiles of U. lactuca collected from intertidal sites and cultivated in an IMTA facility. Our work enhances our understanding of pigments and carotenoids in U. lactuca grown under controlled aquaculture conditions and collected from natural intertidal beds and their responses to storage conditions. Ulva lactuca grown in IMTA systems exhibited higher carotenoid content and better pigment conservation than U. lactuca collected from the intertidal beds, supporting the suitability of IMTA-grown U. lactuca for biotechnological applications.
MATERIALS AND METHODS
Sample collection
The sampling sites were located in Rio de Janeiro, southeastern Brazil. Naturally growing U. lactuca was collected from beds in the rocky intertidal site of Arpoador Beach, (22°59′23.399″ S, 43°11′17.3024″ W) and the estuarine waters of Boa Viagem Beach (22°54′33.79″ S, 43°7′50.4288″ W). Additionally, U. lactuca was collected from the IMTA startup D’Alga Urban Aquaculture. We collected samples of U. lactuca from these locations during dry austral winter (July 2018) and the rainy austral summer (February 2019).
Sample replicates were collected once per season. Collection was carried out on days with tides below 0.4. All collections were performed in the early morning to avoid direct carotenoid degradation due to high light radiation (Cruces et al. 2019). The IMTA-produced U. lactuca was grown with Nile Tilapia (Oreochromis niloticus Linnaeus, 1758) in a closed recirculating seawater system (40,000 L total volume) using a blend of artificial (80%) and natural seawater (20%) collected at Cabo Frio, Rio de Janeiro (22°52′20″ S, 42°0′20″ W).
We cultivated the algal specimens in indoor alga tanks (0.5 m2 and 80 L) covered with ultraviolet (UV) filters. The initial algal densities were ~3 g·L-1. Data loggers (UA-002-08, HOBO Pendant, Onset, Bourne, USA) were used to monitor the water temperature every hour. Ten specimens from each sampling site were used to identify the presence of the Ulva species. Voucher materials of each species were deposited in the Botanical Garden Herbarium in Rio de Janeiro. We used the information from Guiry and Guiry (2024) to determine the taxonomic nomenclature and identify species.
After collection, the algal biomass was transported to the laboratory in a polystyrene box with ice to ensure it was properly preserved. Subsequently, the samples were stored at 4 °C until the next day. The samples were thoroughly rinsed with tap water to remove adhered sediments and other substances. The excess water in the thalli was removed using a lettuce dryer and dried in ambient air with dim light for 20 minutes. Then, 5 ± 0.01 g of thalli were weighed in an analytical balance and stored for subsequent carotenoid conservation analyses or immediately subjected to pigment extraction.
Short-term storage experiments
The U. lactuca thalli from the dry season were used to analyze the conservation of carotenoids. For this experiment, we stored 5 ± 0.01 g of fresh thalli in paper envelopes (in triplicate) for 1 week, 2 weeks, or 4 weeks. The envelopes containing the algal biomass were stored in a dark, ventilated place in the laboratory at ambient temperature. After each storage period, the entire sample inside the paper envelope was submitted for pigment extraction.
Crude extracts of Ulva lactuca for pigment extraction
In their work, Thayer and Björkman (1990) used 5 g of acetone in 5 mL for the extraction of dry and fresh material. The final extraction time was 0-24 h at 4 °C (Thayer and Björkman 1990). The extract was filtered and evaporated in a rotary evaporator using a thermal bath at 40 °C. Finally, the samples were filled with N2, sealed, and stored at -20 °C, as recommended by Lashbroke et al. (2010) for carotenoid conservation of plant extracts.
UV/vis spectrophotometry
The Gauss-Peak Spectra (GPS) method (Kupper et al. 2007) was used to calculate the carotenoid and chlorophyll content in the crude extracts. In specific terms, solutions of crude extracts in acetone were prepared in a 1:1 ratio (weight:volume). Subsequently, the UV/vis absorption spectra between 350 and 700 nm of the solutions were acquired (1-nm interval) using a UV/vis digital spectrophotometer (Spectramax, Molecular Devices, San Jose, USA) integrated to data acquisition software. The absorbance curves were adjusted between 0.5 and 1 mL by dilution in 100% acetone. To obtain the concentration of carotenoids, chlorophylls, and their degradation products in the solutions, we use the GPS equations programmed in Sigma Plot by Kupper et al. (2007). We quantified the β-carotene + zeaxanthin, lutein + antheraxanthin, violaxanthin, neoxanthin, the β-carotene isomerization product aurochrome, chlorophyll (Chl) a, Chl b, derived pheophytin (Phe) a, and Phe b in 100% acetone solutions (Kupper et al. 2007) (Fig. 1). It is worth noting that the individual concentrations of carotenoid pairs were not separated because they had identical absorption spectra between 350 nm and 700 nm. Finally, the concentration of carotenoids in the extract (C extract ) was calculated using the following equation:
where dil is the dilution factor,
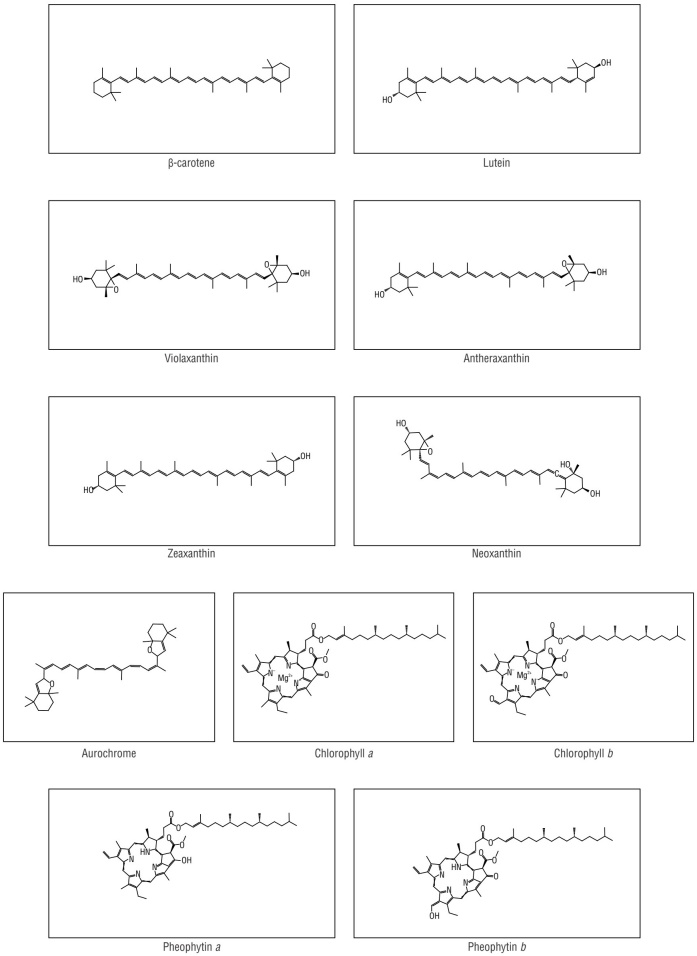
Chemical analysis
Thin layer chromatography (TLC) analyses were performed to evaluate carotenoid conservation using silica gel plates (60 GF254; Merck, Rahway, USA) and an elution solvent of n-hexane:acetone:isopropyl alcohol (69:30:1) (Wright et al. 1997). Each U. lactuca extract (0.5 mg) was spotted on a TLC plate to ensure accurate results. Tentative carotenoid identification was achieved by comparing chromatic profiles and retention factors with the pigment pattern of fresh spinach extract in 100% acetone and information from the literature (Perucka and Oleszek 2000, Rodriguez-Amaya 2001, El-Baky et al. 2008, Mikami and Hosokawa 2013). Additionally, TLC plate adaptation was performed for preparative chromatography. Each extract was chromatographed using 2 different eluent systems (petroleum ether:acetone [7:3] and dichloromethane:ethyl acetate [4:1]). Subsequently, the colored bands were scraped off to isolate compounds with higher spectral purity, which were later confirmed by absorption spectroscopy (350-700 nm) in 100% acetone (Wright et al. 1997). Epoxides were revealed with HCl fume; these structures change color from yellow or orange to blue or green following exposure (Rodriguez-Amaya 2001, Sherma and Fried 2003).
Proton nuclear magnetic resonance (1H-NMR) was used to evaluate carotenoid conservation in the IMTA extracts by comparing the olefinic region of the 1H-NMR spectra (6-7 ppm) according to the quantification methods developed by Valverde and This (2007) and Guadagno et al. (2013). Carotenoid identification was performed using their reported chemical shifts in CDCl3, properly referenced in Putzbach et al. (2005), Sobolev et al. (2005), Valverde and This (2008), and Sivathanu and Palaniswamy (2012). The area below the chemical shifts of the carotenoids was normalized by the tetramethylsilane (TMS) area, according to the methods of Guadagno et al. (2013). The chemical shifts of the solvents, impurities, chlorophylls, and derived molecules (pheophytins and pheophorbide) were discarded from the analyses (Sobolev et al. 2005, Fulmer et al. 2010).
Data analysis
Cochran homogeneity of variance tests were conducted on the data. Parametric data were analyzed using an analysis of variance (ANOVA) and student t-tests. Non-parametric data were evaluated with Kruskal-Wallis or Mann-Whitney tests. All statistical tests were performed in Statistica 7 (StatSoft Inc., Tulsa, USA). Three replicas were used, and the confidence interval for the significance tests was 95% (P = 0.05).
RESULTS
Pigment content in Ulva lactuca fresh biomass
The UV/vis results revealed a higher concentration of carotenoids in fresh U. lactuca thallus extracts during the dry season compared to the rainy season in Arpoador Beach (Fig. 2a). Significant differences were found in β-carotene + zeaxanthin content between the dry and rainy periods at this beach, whereas violaxanthin levels were higher during the rainy period. No significant differences in carotenoid content were observed between the 2 periods in U. lactuca cultivated in the IMTA facility; however, chl a + phe a and chl b + phe b levels were significantly higher during the dry season (Fig. 2b). Ulva lactuca collected from Boa Viagem Beach exhibited a significantly higher carotenoid content during the dry season. Specifically, β-carotene + zeaxanthin and lutein + antheraxanthin were the carotenoids that showed these differences (Fig. 2c). Chlorophyll content was significantly higher in fresh biomass collected from Boa Viagem Beach and the IMTA system during the dry season (Supplementary Material Table S1).
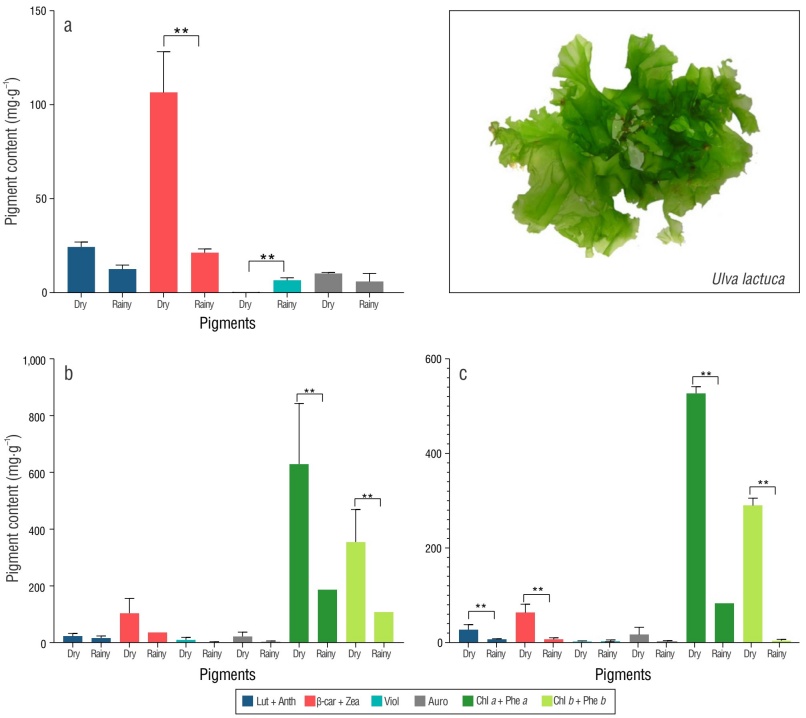
Figure 2 Pigment content (mean ± standard deviation) in Ulva lactuca collected from (a) Arpoador Beach, (b) an integrated multi-trophic aquaculture (IMTA) facility, and (c) Boa Viagem Beach. Pigments: aurochrome (Auro), lutein (Lut) + antheraxanthin (Anth), β-carotene (β-car) + zeaxanthin (Zea), violaxanthin (Viol), chlorophyll (Chl) a + pheophytin (Phe) a, and Chl b + Phe b. *Significant difference (P < 0.05). Pigment content is given per gram of extract.
Pigments conservation during storage
The extracts of U. lactuca thalli cultivated in the IMTA facility exhibited higher levels of carotenoids, chlorophylls, and their degradation products. It is important to highlight that the data used for comparison were from Boa Viagem Beach and the IMTA system, as these values were higher than those of Arpoador Beach.
Pigment conservation was analyzed in U. lactuca biomass collected from the different sites and stored for 1 week, 2 weeks, or 4 weeks. The UV/vis results showed that carotenoid content in U. lactuca from the IMTA facility was well conserved after 4 weeks of storage. However, U. lactuca collected from Arpoador Beach lost β-carotene + zeaxanthin content during the first week of storage and then lost violaxanthin content in the fourth week of storage (Supplementary Material Table S2). In addition, chlorophylls and derived pheophytins were well conserved in the stored biomass collected from the IMTA facility and Arpoador Beach.
Chemical analyses to monitor pigments during storage
Thin layer chromatography profiles were employed to complement the UV/vis results. A noticeable tendency toward color loss in the β-carotene band in extracts from Arpoador Beach and Boa Viagem Beach after 4 weeks of storage was evident, compared to what was observed after 1 week or 2 weeks of storage. Conversely, the intensity of the β-carotene band in U. lactuca from the IMTA facility was maintained. In comparison, the TLC profile of the fresh IMTA extract exhibited more intense bands and an additional yellow band. The yellow and orange bands in these extracts were tentatively identified as β-carotene, lutein + zeaxanthin, and neoxanthin. The additional band observed in the IMTA extract was tentatively identified as violaxanthin. Dried stored samples were analyzed through 1H-NMR; the olefinic region (6-7 ppm) of the spectra was chosen because it accumulates the hydrogen signals of carotenoids (Sobolev et al. 2005). Carotenoids signals were highlighted in this region to aid analyses.
The carotenoid profile was monitored during storage via olefinic signal patterns observed in 1H-NMR spectra, specifically in the chemical shift region below 6.60-6.64 ppm, which is common to all carotenoids (Sobolev et al. 2005). Algae spectra collected from Boa Viagem Beach and IMTA facility showed effective carotenoid preservation during the initial 2 weeks of storage, with noticeable degradation observed only after 4 weeks. In contrast, the spectra of the samples collected from Arpoador Beach showed a significant decrease in signal intensity between fresh and dried stored algae, indicating a decline in carotenoid content from fresh to dried states, which was likely due to degradation or isomerization during the first week of storage.
DISCUSSION
The U. lactuca collected from the IMTA facility presented significantly higher carotenoid and chlorophyll yields in biomass extracts compared to the samples collected from the intertidal beds, indicating that photosynthetic pigments were promoted. The IMTA facility, unlike the natural beds, was not subject to precipitation, desiccation, winds, grazers, or UV radiation; variables, such as salinity and pH, were controlled in the IMTA facility. Thus, nutrient enrichment and a controlled environment may have enhanced the physiology and growth of U. lactuca, preventing oxidation reactions that degrade carotenoids while inducing pigment synthesis (Eismann et al. 2020).
Moreover, U. lactuca grown in the IMTA facility conserved its carotenoid content during 4 weeks of storage, while some carotenoid losses were observed in the stored biomass of U. lactuca collected from intertidal beds. The TLC and 1H-NMR analyses indicated that the pigment profiles of U. lactuca grown in the IMTA facility and collected from the intertidal beds were conserved in extracts stored for 1 week. However, a loss of carotenoids was observed after 2 weeks and 4 weeks of storage in U. lactuca collected from the intertidal beds. This conservation of pigments may be related to the presence of other compounds with antioxidant properties reported in U. lactuca such as tocopherols and tocotrienols (Ortiz et al. 2006), olefins (Ratnayake et al. 2013), sesquiterpenoids, (Chakraborty and Paulraj 2010), and ulvan (Alves et al. 2013). Pigment production and carotenoid coproduction are interesting topics to explore in future research due to the biotechnological potential of these molecules, which have garnered interest across various industrial sectors, including the food, pharmaceutical, cosmeceutical, and nutraceutical industries.
Overall, the U. lactuca extracts were rich in chlorophylls, β-carotene + zeaxanthin, and lutein + antheraxanthin. The content of violaxanthin in IMTA extracts may be associated with a lower oxidation state of the thylakoid membranes (Zhang et al. 2015), as well as higher overall pigment synthesis. The current carotenoid market is predominantly led by β-carotene and lutein, as these essential pigments are utilized in animal feed and dietary supplements. Thus, the abundant presence of violaxanthin in U. lactuca collected from the IMTA facility is particularly promising. This compound swiftly transforms into zeaxanthin through the xanthophyll cycle (Zhang et al. 2015), and the antioxidant properties of zeaxanthin have been well documented (Sajilata et al. 2008, Esteban et al. 2014, Meléndez-Martinez et al. 2022). Thus, IMTA systems could emerge as novel and sustainable sources of zeaxanthin.
The significant differences in the yields of carotenoids and chlorophylls in the fresh biomass of U. lactuca collected during the rainy and dry seasons may be associated with the macroalgae being affected by a wide range of biotic and abiotic conditions (e.g., temperature, hydrodynamics, desiccation, and biological interactions), which directly influence growth and pigment content (Fort et al. 2019, Jiang et al. 2019, Calheiros et al. 2021). On the other hand, the yields of U. lactuca collected from the IMTA facility reflected less variation in growth conditions; the few differences recorded between seasons were associated with photosynthetically active radiation (PAR) and water temperature.
Both PAR and temperature were higher in the rainy season, coinciding with the lower production of carotenoids and chlorophylls that we observed in this work (Narrain et al. 2023). Some studies have identified a negative correlation between photosynthetic pigments and solar radiation in Ulva rotundata (Henley et al. 1989) and in Ulva species collected from natural beds (Figueroa et al. 2003). It is important to highlight that high acclimation to PAR involves a decrease in photosynthetic pigments as a response to the relocation of thylakoid membranes and antenna complexes (Esteban et al. 2015).
Our results indicate that U. lactuca cultivated in the IMTA facility showed higher levels of carotenoids and better preservation than U. lactuca collected from intertidal beds. The algae collected in natural beds showed greater variability and complexity, which may be attributed to the influence of various environmental factors, such as tidal variations, PAR radiation, and periods of desiccation. Further sampling efforts and environmental analyses are necessary to provide conclusive evidence of metabolite production and storage in U. lactuca collected from natural environments.
CONCLUSIONS
The analyses conducted in this study have revealed that cultivation systems, such as IMTA, represent an excellent option to control environmental factors, which allows for macroalgae biomass and pigments to be produced with the specific qualities required for biotechnological applications. It is noteworthy that U. lactuca grown in the IMTA system demonstrated higher carotenoid content and superior pigment conservation over 4 weeks of storage compared to biomass collected from intertidal sites. Additionally, the fresh biomass extracts exhibited lower pigment content during the rainy season, which correlated with increased PAR radiation and water temperature. Cultivation systems for U. lactuca represent a promising and sustainable source of carotenoids, including β-carotene, lutein, and violaxanthin. These systems can also be optimized to produce zeaxanthin, which exhibits high nutritional value and antioxidant capacity, for biotechnological and industrial applications.

