











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
INTRODUCTION
The family Sparidae comprises teleost fishes that are widely distributed in tropical and temperate waters (Sheaves 2006, Pavlidis and Mylonas 2011). Most sparids are carnivorous and feed on benthic invertebrates (Smith 1938, Siddiqui et al. 2022), although some sparids are herbivores or omnivores that ingest large amounts of plant material (Chavance et al. 1986, Castillo-Rivera et al. 2007). It should be noted that sparids exhibit a wide variety of mating systems and reproductive strategies, such as gonochorism and simultaneous and sequential hermaphroditism (Buxton and Garrat 1990, De Mitcheson and Liu 2008), and relatively long reproductive periods (Houde 1975, Pavlidis and Mylonas 2011). Several members of this family are important for recreational riverine fisheries due to their high economic value, with the catch destined for human consumption (Hoşsucu and Cakir 2003, Pavlidis and Mylonas 2011).
The Pacific Porgy (Calamus brachysomus) is the only representative of the family Sparidae that is distributed in the eastern tropical Pacific and Gulf of California (Salas et al. 2011). Juveniles inhabit shallow waters over mixed sandy and rocky substrates. Adults are found on sandy bottoms and at the margins of rocky reefs at a maximum depth of ~80 m (Eschmeyer et al. 1983, Thomson 1987). Although this species is not the target of a directed fishery, it is caught in high volumes as bycatch in multispecific riverine fisheries in Mexico (CONAPESCA 2021) and is common in regional (Baja California Sur) and national markets (Veranes-Dip et al. 2022).
Biological studies of the Pacific Porgy based on samples obtained in Bahía Magdalena, Baja California Sur (BCS), have included descriptions of eggs and larvae and reported hatching temperatures of 20 to 26 °C (Matus-Nivón et al. 1987, 1989). This species reproduces year-round in Santa Rosalía, BCS, with 2 reproductive peaks (May-July and November) when the sea surface temperature (SST) ranges from 21 to 28 °C (Veranes-Dip et al. 2022).
The only prior study on the diet of the Pacific Porgy reported that the main prey items of C. brachysomus were invertebrates, such as bivalves, gastropods, echinoderms, and crustaceans, and occasionally small fishes (Abitia-Cárdenas et al. 1990). Another study, which grouped the Pacific Porgy with 7 other species in the category of small demersal fishes, found that its overall diet consisted of bivalves, gastropods, crabs, shrimp, and sessile invertebrates (Ainsworth et al. 2009). Currently, there are no published studies on many important aspects of the feeding habits of this species or how diet could be affected by multiple factors, such as reproductive condition or the physical characteristics of habitats.
Considering that feeding strategies can vary during the reproductive period in response to energy demands, especially during maturation and spawning (Luquet and Watanabe 1986, Saborido-Rey 2004), information on the feeding habits of C. brachysomus during different reproductive stages will be useful for maximizing the reproductive effort of the species in aquaculture settings (Mylonas et al. 2010). Given the growing interest in C. brachysomus as a fishery resource, the lack of information on its biology must be addressed. Therefore, the objectives of the present study were to characterize the feeding habits of the Pacific Porgy in Santa Rosalía, BCS (central Gulf of California), and to analyze possible variations in diet as a function of sex, season, and reproductive stage.
MATERIALS AND METHODS
Study area
Monthly sampling was conducted from July 2015 to July 2016. In all, 235 C. brachysomus specimens were collected. Specimens were obtained from commercial catches made by the fishers of Santa Rosalía in the central region of the Baja California Peninsula, Mexico (Fig. 1). A total of 480 coastal fishing boats (6.7-10 m in length) operated in the area. Calamus brachysomus was caught using gillnets (10-cm mesh) installed between 10 and 35 m depth (Pérez-Rojo et al. 2022).
Figure 1 Study area. Black dots indicate Calamus brachysomus sampling sites in Santa Rosalía (27°20′23″ N, 112°15′41″ W), Baja California Sur, Mexico.
The total length (TL ± 0.1 cm) and total weight (TW ± 0.1 g) of each specimen were recorded. Sex and reproductive stage were determined based on the histological gonadal characterization of Veranes-Dip et al. (2022). These authors established 3 groups based on reproductive stage: developing, spawning-capable, and post-spawning. We employed these previously established reproductive-stage groups in this study.
Monthly data of SST were obtained from satellite images downloaded from Scripps Institution of Oceanography (https://scripps.ucsd.edu/). These images were processed using Windows Image Manager (WIMSoft, Windows, Redmond, USA). The mean SST value was calculated, and months with temperatures above and below this mean value (anomalies) were considered warm and cold periods, respectively. Two climatic seasons were identified: a cold season (November 2015 to May 2016) and a warm season (July to October 2015 and June to July 2016) (Fig. 2).
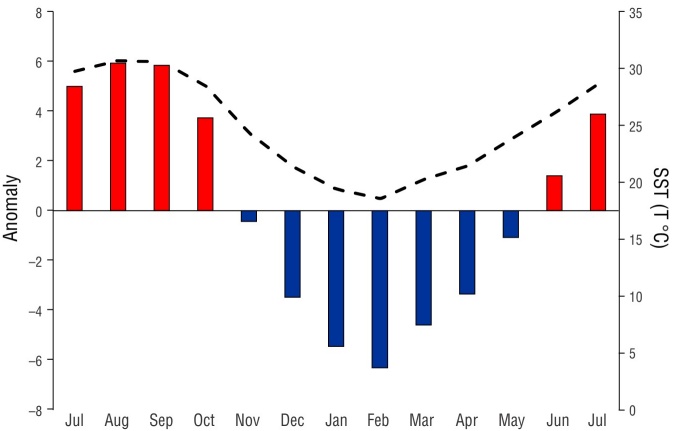
Figure 2 Sea surface temperature (SST) by month (dashed line) in the central Gulf of California from July 2015 to July 2016. The primary axis shows anomalies with respect to the mean; the secondary axis shows the temperature in degrees Celsius. Black bars indicate warm months (months with SST above the mean value), and grey bars indicate cold months (months with SST below the mean value).
Trophic analysis
Stomach contents were classified by taxonomic group, and each item was identified to the lowest possible taxonomic level. A stereoscopic Stemi 2000-C microscope (Zeiss, Oberkochen, Germany) and specialized literature were used to identify prey of the following groups: echinoderms (Hendrickx 1995a, Solís-Marín et al. 2005, Hernández-Herrejón et al. 2008), gastropods (Baqueiro et al. 1982, Poutiers 1995b, Hendrickx et al. 2014, Aguilar-Estrada et al. 2017), bivalves (Holguín-Quiñones and González-Pedraza 1994, Poutiers 1995a), malacostracans (Hendrickx 1995b, 1995c; Salgado-Barragán and Hendrickx 2010), and other invertebrates such as annelids, poriferans, and isopods (Brusca 1980, Morris et al. 1980). All taxonomic names were verified using the World Register of Marine Species (WoRMS). We analyzed all data (overall) and data by category: sex (male or female), reproductive stage (developing, spawning-capable, or post-spawning), and season (cold or warm).
Accumulation curves of prey in C. brachysomus stomachs were analyzed for all data (overall) and data by category (sex, reproductive stage, and season). This involved comparing the cumulative counts of prey species in the stomach contents using the Clench equation to determine the slope. A slope of less than 0.1 (a value) indicated an asymptotic curve and validated the representativeness of the collected prey items. Furthermore, the proportion of recorded prey species, which was used to determine the representativeness of the inventory of stomach contents, ranged from zero (not representative) to 100% (highly representative) (Jiménez-Valverde and Hortal 2003). The curves, model, and slope plots were generated using the ‘Vegan’ package in R v. 3.0.1 (Oksanen et al. 2016).
The prey-specific index of relative importance (%PSIRI), which was proposed by Brown et al. (2012), was used to quantitatively analyze the diet of the Pacific Porgy. This index quantifies the abundance of prey in the stomach of a predator using the following equation:
where %PSIRI is the relative importance of the prey-specific index, %FO is the frequency of occurrence, %PN is the percentage by number of the specific prey, and %PW is the percentage by weight of the specific prey.
Trophic breadth was calculated using the standardized Levin index (Hurlbert 1978). Index values below 0.6 indicated a specialist diet, and values above 0.6 indicated a generalist diet (Krebs 1999). This index was calculated using the following equation:
where B i is the trophic breadth of predator i, p ij is the proportion of prey j in the diet of predator i, and n is the total number of prey.
The Costello method (1990), with modifications by Amundsen et al. (1996), was used to interpret the feeding strategy of the Pacific Porgy for all data (overall) and data by category (sex, reproductive stage, and season) and to corroborate the trophic niche breadth results. Four feeding strategies can be identified with the Costello method: (1) a specialist consuming several trophic resources, (2) a generalist with individual variations in trophic breadth, (3) a specialist consuming one predominant prey type while also occasionally consuming other prey, and (4) a mixed strategy in which some individuals in the population exhibit specialist diets and others exhibit generalist diets (Amundsen et al. 1996). We used TrophLab, which can be downloaded from FishBase (https://www.fishbase.org) (Pauly et al. 2000), to estimate trophic position. This software uses the equation developed by Christensen and Pauly (1992) to calculate the trophic level of predators and information from fishbase.org to determine the trophic level of prey.
Statistical analysis
A permutational multivariate analysis of variance (PERMANOVA) with 1,000 permutations was used to evaluate possible differences in diet with respect to sex, season, and reproductive stage using the ‘Adonis()’ function in the ‘Vegan 2.2-1’ package (Oksanen et al. 2016) in R v. 3.0.1. The PERMANOVA was based on the Bray-Curtis dissimilarity distance and employed a 95% significance level.
RESULTS
Trophic spectrum
A total of 235 C. brachysomus specimens (108 males and 127 females) were collected, all of which contained food in their stomachs. Specimens measured between 15 and 37 cm TL and weighed between 120 and 860 g TW. The prey accumulation curves exhibited good fit; the number of prey species was close to the asymptote predicted by the model for the entire sample (overall) and for each category (sex, reproductive stage, and season). The slope values ranged between 0.06 and 0.09 (except for reproductive stage and cold season, which had slope values of 0.1), and the proportion of recorded prey species ranged between 84% and 97% (Table 1, Fig. 3).
Table 1 Prey accumulation curve based on the stomach contents of Calamus brachysomus for all data (overall) and by category (sex, reproductive stage, and season).
| Category | Ns | a | RF | R 2 |
| General | 235 | 0.06 | 96.61 | 0.99 |
| Sex | ||||
| Female | 127 | 0.08 | 94.07 | 0.99 |
| Male | 108 | 0.08 | 92.78 | 0.99 |
| Reproductive stage | ||||
| Developing | 89 | 0.1 | 91.88 | 0.99 |
| Spawning-capable | 60 | 0.08 | 84.00 | 0.99 |
| Post-spawning | 86 | 0.09 | 91.18 | 0.99 |
| Season | ||||
| Cold season | 120 | 0.1 | 94.80 | 0.99 |
| Warm season | 115 | 0.06 | 90.67 | 0.99 |
Ns: number of analyzed stomachs; a: slope; RF: registered fauna; R 2: coefficient of determination.
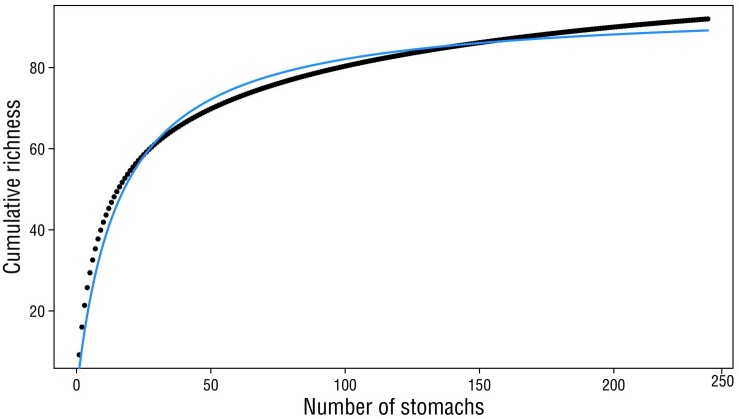
Figure 3 Cumulative richness curve used to characterize the general feeding habits of Calamus brachysomus. The black points are species richness, and the blue line is the Clench model.
The diet comprised 90 prey items, including 58 mollusks, 14 echinoderms, 13 arthropods, 2 chordates, 11 annelids, and 11 poriferans, as well as unidentified algae. In general, the most important prey based on %PSIRI were the bivalves Transennella spp. (7.24%), Argopecten spp. (6.94%), Anadara spp. (6.25%), Septifer zeteki (6.19%), Tivela spp. (6.1%), and Glycymeris spp. (5.59%) (Fig. 4, Table 2).
Table 2 Calamus brachysomus prey in Santa Rosalía, Mexico. Values for each prey item by number (N), weight (W), and frequency of occurrence (FO) expressed as absolute numbers and percentages. The results of these indices were integrated into the prey-specific index of relative importance (%PSIRI).
| Species | FO | N | W | %FO | %N | %W | %PSIRI | ||
| Mollusca | |||||||||
| Bivalvia | Lucinidae | 21 | 27 | 1.04 | 8.94 | 0.73 | 0.68 | 1.02 | |
| Arcidae | Anadara spp. | 77 | 194 | 22.13 | 32.77 | 5.28 | 14.54 | 6.25 | |
| Arca spp. | 36 | 42 | 1.27 | 15.32 | 1.14 | 0.84 | 1.34 | ||
| Barbatia spp. | 8 | 9 | 0.94 | 3.4 | 0.24 | 0.62 | 0.66 | ||
| Americardia spp. | 42 | 66 | 6.57 | 17.87 | 1.8 | 4.32 | 2.91 | ||
| Papyridea spp. | 55 | 89 | 1.88 | 23.4 | 2.42 | 1.24 | 2.28 | ||
| Corbulidae | Corbula spp. | 30 | 37 | 1.31 | 12.77 | 1.01 | 0.86 | 1.40 | |
| Glycymerididae | Tucetona spp. | 13 | 27 | 1.72 | 5.53 | 0.73 | 1.13 | 0.97 | |
| Hiatellidae | Glycymeris spp. | 62 | 231 | 15.93 | 26.38 | 6.28 | 10.46 | 5.59 | |
| Mytilidae | Septifer zeteki | 111 | 269 | 4.66 | 47.23 | 7.32 | 3.06 | 6.19 | |
| Pectinidae | 2 | 2 | 0.09 | 0.85 | 0.05 | 0.06 | 0.06 | ||
| Argopecten spp. | 105 | 232 | 8.18 | 44.68 | 6.31 | 5.37 | 6.94 | ||
| Pteriidae | Pteria sterna | 16 | 32 | 0.56 | 6.81 | 0.87 | 0.37 | 0.73 | |
| Veneridae | Transennella spp. | 109 | 226 | 7.04 | 46.38 | 6.15 | 4.63 | 7.24 | |
| Tivela spp. | 76 | 252 | 16.11 | 32.34 | 6.85 | 10.58 | 6.1 | ||
| Chioneryx spp. | 71 | 129 | 1.7 | 30.21 | 3.51 | 1.12 | 2.78 | ||
| Chione spp. | 24 | 28 | 1.42 | 10.21 | 0.76 | 0.93 | 1.51 | ||
| Leukoma spp. | 20 | 37 | 0.57 | 8.51 | 1.01 | 0.37 | 1.16 | ||
| Gastropoda | Patellogastropoda | 8 | 8 | 2.36 | 3.4 | 0.22 | 1.55 | 0.91 | |
| Architectonicidae | Heliacus spp. | 20 | 21 | 0.16 | 8.51 | 0.57 | 0.11 | 0.37 | |
| Buccinidae | 3 | 3 | 0.02 | 1.28 | 0.08 | 0.01 | 0.08 | ||
| Bullidae | Bulla spp. | 37 | 54 | 0.45 | 15.74 | 1.47 | 0.3 | 0.99 | |
| Calyptraeidae | Crepidula spp. | 26 | 28 | 0.71 | 11.06 | 0.76 | 0.47 | 0.84 | |
| Cerithiopsidae | Seila spp. | 16 | 22 | 0.09 | 6.81 | 0.6 | 0.06 | 0.05 | |
| Columbellidae | Mitrella millepunctata | 20 | 42 | 0.39 | 8.51 | 1.14 | 0.26 | 0.91 | |
| Drilliidae | Splendrillia spp. | 27 | 144 | 0.45 | 11.49 | 3.92 | 0.3 | 1.68 | |
| Eratoidae | Hespererato spp. | 11 | 11 | 0.08 | 4.68 | 0.3 | 0.05 | 0.18 | |
| Eulimidae | Eulima spp. | 1 | 1 | 0 | 0.43 | 0.03 | 0 | 0.03 | |
| Fasciolariidae | 2 | 2 | 0.04 | 0.85 | 0.05 | 0.03 | 0.08 | ||
| Latirus spp. | 21 | 28 | 0.18 | 8.94 | 0.76 | 0.11 | 0.46 | ||
| Ficidae | 7 | 10 | 0.02 | 2.98 | 0.27 | 0.01 | 0.11 | ||
| Fissurellidae | Fissurella spp. | 15 | 20 | 0.61 | 6.38 | 0.54 | 0.4 | 0.44 | |
| Mangeliidae | Steironepion spp. | 23 | 31 | 0.15 | 9.79 | 0.84 | 0.1 | 0.57 | |
| Agathotoma spp. | 4 | 4 | 0.01 | 1.7 | 0.11 | 0.01 | 0.08 | ||
| Marginellidae | Volvarina taeniolata | 27 | 40 | 0.17 | 11.49 | 1.09 | 0.11 | 0.72 | |
| Volvarina spp. | 28 | 37 | 0.14 | 11.91 | 1.01 | 0.09 | 0.57 | ||
| Mitromorphidae | Mitromorpha spp. | 3 | 12 | 0.52 | 1.28 | 0.33 | 0.34 | 0.26 | |
| Nassariidae | Nassarius spp. | 45 | 83 | 2.18 | 19.15 | 2.26 | 1.43 | 1.51 | |
| Naticidae | Polinices uber | 33 | 36 | 0.43 | 14.04 | 0.98 | 0.28 | 0.91 | |
| Pisaniidae | Engina jugosa | 11 | 25 | 0.36 | 4.68 | 0.68 | 0.24 | 0.51 | |
| Olivellidae | 8 | 9 | 0.21 | 3.4 | 0.24 | 0.14 | 0.21 | ||
| Planaxidae | 8 | 11 | 0.07 | 3.4 | 0.3 | 0.05 | 0.17 | ||
| Planaxis spp. | 1 | 1 | 0 | 0.43 | 0.03 | 0 | 0.06 | ||
| Potamididae | Cerithidea spp. | 1 | 1 | 0 | 0.43 | 0.03 | 0 | 0.03 | |
| Rissoinidae | Rissoina spp. | 43 | 90 | 1.09 | 18.3 | 2.45 | 0.72 | 1.44 | |
| Pseudomelatomidae | Crassispira spp. | 31 | 72 | 0.66 | 13.19 | 1.96 | 0.43 | 1.51 | |
| Pilsbryspira spp. | 37 | 69 | 0.57 | 15.74 | 1.88 | 0.38 | 1.18 | ||
| Tegulidae | Tegula spp. | 54 | 87 | 1.6 | 22.98 | 2.37 | 1.05 | 1.72 | |
| Siphonariidae | 9 | 11 | 0.34 | 3.83 | 0.3 | 0.23 | 0.25 | ||
| Williamia spp. | 1 | 2 | 0.02 | 0.43 | 0.05 | 0.01 | 0.05 | ||
| Terebridae | Terebra spp. | 40 | 78 | 0.8 | 17.02 | 2.12 | 0.53 | 1.37 | |
| Tornatinidae | 2 | 3 | 0.01 | 0.85 | 0.08 | 0 | 0.05 | ||
| Triphoridae | Triphora spp. | 35 | 63 | 0.19 | 14.89 | 1.71 | 0.12 | 0.82 | |
| Trochidae | Turbo spp. | 1 | 9 | 1.37 | 0.43 | 0.24 | 0.65 | 0.11 | |
| Turridae | 13 | 15 | 0.07 | 5.53 | 0.41 | 0.04 | 0.23 | ||
| Velutinidae | Lamellaria inflata | 1 | 1 | 0.14 | 0.43 | 0.03 | 0.09 | 0.09 | |
| Polyplacophora | Chitonidae | 19 | 34 | 0.61 | 8.09 | 0.92 | 0.4 | 0.72 | |
| Scaphopoda | 3 | 3 | 0.01 | 1.28 | 0.08 | 0 | 0.07 | ||
| Porifera | 2 | 2 | 0.34 | 0.85 | 0.05 | 0.22 | 0.17 | ||
| Echinodermata | 1 | 1 | 0.01 | 0.43 | 0.03 | 0.01 | 0.03 | ||
| Asteroidea | Astropectinidae | Astropecten armatus | 3 | 3 | 1.85 | 1.28 | 0.08 | 1.22 | 0.46 |
| Ophidiasteridae | Phataria unifascialis | 5 | 5 | 0.33 | 2.13 | 0.14 | 0.22 | 0.04 | |
| Ctenodiscidae | Ctenodiscus crispatus | 6 | 7 | 0.1 | 2.55 | 0.19 | 0.06 | 0.13 | |
| Echinoidea | Echinometridae | Echinometra vanbrunti | 106 | 113 | 5.15 | 45.11 | 3.07 | 3.39 | 3.93 |
| Cidaridae | Eucidaris thouarsii | 6 | 6 | 0.94 | 2.55 | 0.16 | 0.62 | 0.62 | |
| Toxopneustidae | Toxopneustes roseus | 5 | 5 | 0.07 | 2.13 | 0.14 | 0.05 | 0.19 | |
| Holothuroidea | 3 | 9 | 4.03 | 1.28 | 0.24 | 2.65 | 0.45 | ||
| Ophiuroidea | 11 | 11 | 1.07 | 4.68 | 0.3 | 0.7 | 0.37 | ||
| Ophiocomidae | Ophiocoma spp. | 16 | 16 | 5.09 | 6.81 | 0.44 | 3.34 | 0.85 | |
| Ophiodermatidae | Ophioderma panamense | 3 | 3 | 0.46 | 1.28 | 0.08 | 0.3 | 0.26 | |
| Ophiotrichidae | Ophiothela mirabilis | 5 | 6 | 0.04 | 2.13 | 0.16 | 0.03 | 0.16 | |
| Amphiuridae | Amphiodia spp. | 1 | 1 | 0.07 | 0.43 | 0.03 | 0.05 | 0.09 | |
| Asteronychidae | Asteronyx spp. | 1 | 1 | 0 | 0.43 | 0.03 | 0 | 0.03 | |
| Annelida | 5 | 6 | 1.21 | 2.13 | 0.16 | 0.79 | 0.27 | ||
| Arthropoda | |||||||||
| Malacostraca | Alpheidae | Alpheus lottini | 86 | 119 | 4.82 | 36.6 | 3.24 | 3.17 | 4.65 |
| Xanthidae | Nanocassiope polita | 54 | 58 | 2.94 | 22.98 | 1.58 | 1.93 | 2.81 | |
| Portunidae | Euphylax robustus | 35 | 49 | 6.14 | 14.89 | 1.33 | 4.03 | 2.56 | |
| Panopeidae | Eurypanopeus spp. | 22 | 29 | 1.68 | 9.36 | 0.79 | 1.11 | 1.23 | |
| Mithracidae | Mithrax spp. | 9 | 13 | 0.7 | 3.83 | 0.35 | 0.46 | 0.55 | |
| Calappidae | 2 | 3 | 0.8 | 0.85 | 0.08 | 0.52 | 0.22 | ||
| Varunidae | Hemigrapsus oregonensis | 4 | 5 | 1.02 | 1.7 | 0.14 | 0.67 | 0.30 | |
| Pinnotheridae | 2 | 3 | 0.05 | 0.85 | 0.08 | 0.03 | 0.08 | ||
| Cancridae | 2 | 2 | 0.14 | 0.85 | 0.05 | 0.09 | 0.11 | ||
| Parthenopidae | 1 | 1 | 0 | 0.43 | 0.03 | 0 | 0.03 | ||
| Leiolambrus spp. | 1 | 1 | 0.01 | 0.43 | 0.03 | 0 | 0.04 | ||
| Isopoda | 2 | 2 | 0.01 | 0.85 | 0.05 | 0.01 | 0.07 | ||
| Stomatopoda | 1 | 1 | 0.06 | 0.43 | 0.03 | 0.04 | 0.07 | ||
| Chordata | |||||||||
| Ascidiacea | 6 | 10 | 0.92 | 2.55 | 0.27 | 0.6 | 0.63 | ||
| Actinopterygii | 2 | 2 | 0.89 | 0.85 | 0.05 | 0.58 | 0.14 | ||
| Unidentified algae | 30 | 0 | 0.85 | 12.77 | 0 | 0.56 | 0.92 |
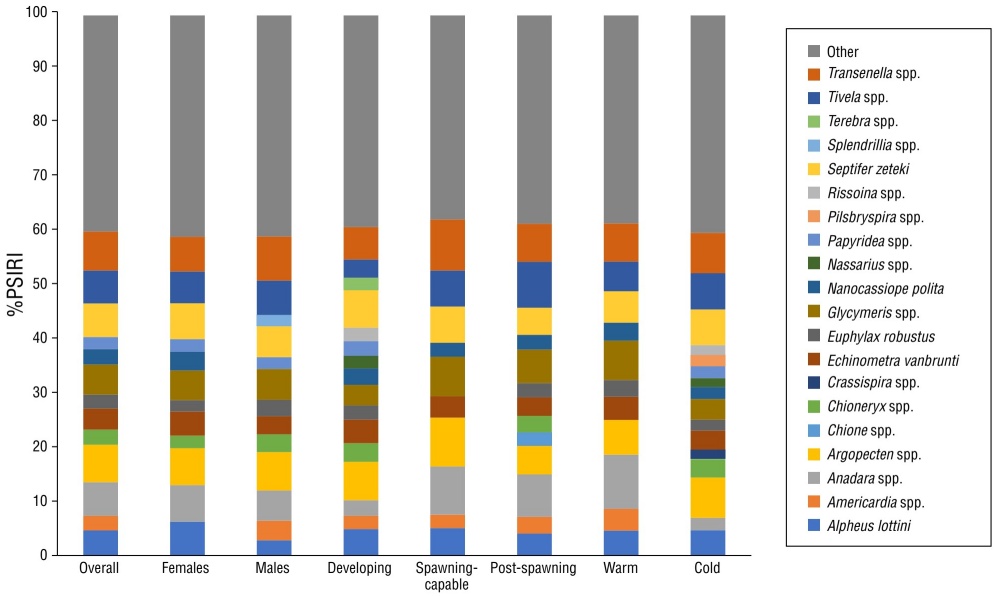
Figure 4 Percentages of the prey-specific index of relative importance (%PSIRI) for the main prey items consumed by Calamus brachysomus in the central Gulf of California, Mexico, for all data (overall) and data by category: sex (male or female), season (warm or cold), and reproductive stage (developing, spawning-capable, and post-spawning).
By sex
The trophic spectrum analysis by sex included 108 stomachs from males and 127 stomachs from females. The trophic spectrum of males comprised 83 prey items. Based on %PSIRI, the most important prey items were the bivalves Transennella spp. (8.2%), Argopecten spp. (7.09%), Tivela spp. (6.36%), Glycymeris spp. (5.71%), and S. zeteki (5.7%). The trophic spectrum of females comprised 80 prey items. Based on %PSIRI, the most important prey items were the bivalves Argopecten spp. and Anadara spp. (6.82%), S. zeteki (6.61%), and Transennella spp. (6.43%) and the crustacean Alpheus lotinni (6.22%) (Fig. 4). The PERMANOVA analysis did not result in significant differences between males and females (F (1) = 1.47, P > 0.09) (Table 3).
Table 3 Permutational multivariate analysis of variance (PERMANOVA) results of abundance data for prey in the diet of Calamus brachyosomus by sex, season, reproductive stage, and interactions.
| Category | d.f. | F | P |
| Sex | 1 | 1.47 | 0.095 |
| Season | 1 | 5.25 | 0.001* |
| Reproductive phase | 2 | 1.42 | 0.06 |
| Sex: Season | 1 | 1.73 | 0.06 |
| Sex: Reproductive phase | 2 | 1.7 | 0.09 |
| Season: Reproductive phase | 2 | 1.34 | 0.08 |
* Indicates significant differences, d.f. = degrees of freedom, F = PERMANOVA, P = probability value
By reproductive stage
The trophic spectrum analysis by reproductive stage included 74 specimens in the developing stage, 73 in the spawning-capable stage, and 76 in the post-spawning stage. Based on %PSIRI, the most important prey items in the stomach contents of specimens in the developing stage were the bivalves Tivela spp. (8.51%), Anadara spp. (7.85%), Transennella spp. (7.02%), Glycymeris spp. (6.22%), and Argopecten spp. (5.33%). Based on %PSIRI, the most important prey items the stomach contents of specimens in the spawning-capable stage were the bivalves Transennella spp. (9.43%), Argopecten spp. (9.04%), Anadara spp. (8.93%), and Glycymeris spp. (7.33%). Based on %PSIRI, the most important prey items in the the stomach contents of specimens in the post-spawning stage were the bivalves Tivela spp. (8.51%), Anadara spp. (7.85%), Transennella spp. (7.02%), and Glycymeris spp. (6.22%) (Fig. 4). There were no significant differences in the stomach contents of specimens in different reproductive stages (F (1) = 1.42, P > 0.06) (Table 3).
By season
The trophic spectrum analysis by season included 15 specimens sampled during the warm season and 120 specimens sampled during the cold season. The stomach contents of specimens collected in the warm season included 80 prey items. Based on %PSIRI, the most important prey items were the bivalves Anadara spp. (10.05%), Glycymeris spp. (7.28%), Transennella spp. (7.04%), and Argopecten spp. (6.43%). The stomach contents of specimens collected in the cold season also included 80 prey items. Based on %PSIRI, the most important prey items were the bivalves Argopecten spp. (7.48%), Transennella spp. (7.45%), Tivela spp. (6.73%), and S. zeteki (6.59%) (Fig. 4). The PERMANOVA indicated significant differences in diet between the warm and cold seasons (F (1) = 5.2, P < 0.001). There were no significant differences in diet resulting from the interactions between sex and season (F (1,1) = 1.73, P > 0.06) or between sex and reproductive phase (F (1,2) = 1.7, P > 0.09) (Table 3).
Trophic analysis
According to the Levin index, the feeding strategy of the Pacific Porgy revealed this species to be a predator with a narrow trophic niche (B i = 0.31). This feeding strategy remained unaltered when considering the categories of sex (females: B i = 0.33; males: B i = 0.34), reproductive stage (developing stage: B i = 0.37; spawning-capable stage: B i = 0.26; post-spawning stage: B i = 0.37), and season (cold season: B i = 0.36; warm season: B i = 0.29). The Costello graphs indicated that C. brachysomus consumed mainly benthic prey, the most important of which were the bivalves Argopecten spp., S. zeteki, Transennella spp., and Anadara spp. These prey items were relatively abundant and frequently consumed, but their importance in the diet of C. brachysomus varied according to the analyzed categories (Fig. 5).
Considering all data, the trophic level of C. brachysomus was 3.18. The trophic level was also calculated by sex (3.16 for males and females), reproductive stage (3.17 for specimens in the developing stage, 3.18 for specimens in the spawning-capable stage, and 3.14 for specimens in the post-spawning stage), and season (3.15 during the cold season and 3.26 during the warm season).
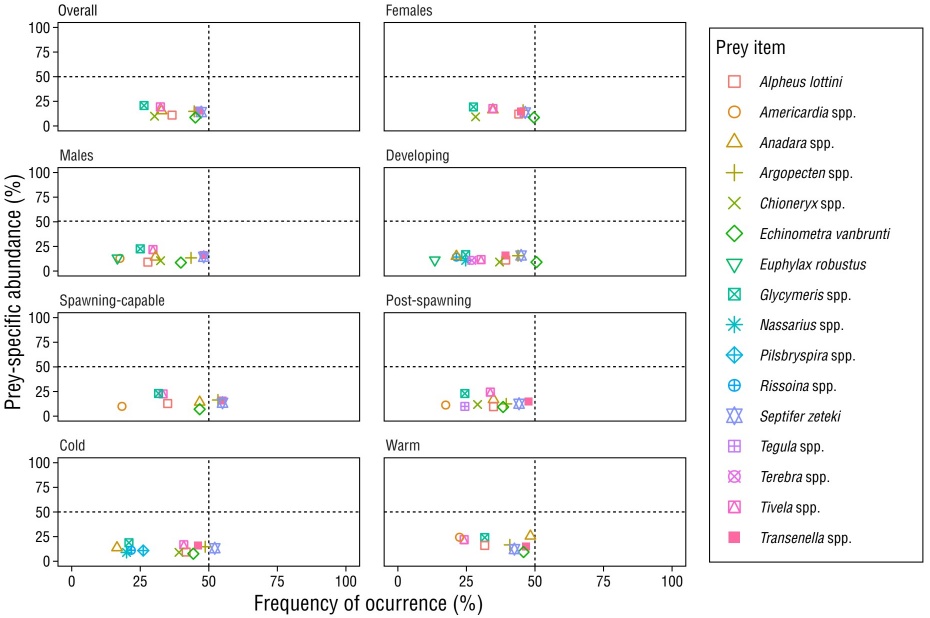
Figure 5 Feeding strategies of Calamus brachysomus represented with Costello graphs. Prey-specific abundance (%N) and frequency of occurrence (%FO) in the diet of C. brachysomus in the central Gulf of California, Mexico, for all data (overall) and data by sex (females and males), reproductive stage (developing, spawning-capable, and post-spawning), and season (cold season and warm season).
DISCUSSION
Feeding habits
Our results confirm that C. brachysomus is a predator that feeds mainly on benthic invertebrates and occasionally on small fish. This was verified by the fish remains found in the stomachs of 2 individuals; there were also algal remains found in some stomachs. These results coincide with what was found by Abitia-Cárdenas et al. (1990), who identified bivalves, crustaceans, echinoderms, some small fishes, and algal remains in the stomach contents of Pacific Porgy specimens from Bahía Concepción in the Gulf of California. During the 9 months of our study, algal remains were found in 2 to 33 individuals per month (30 stomachs contained algal remains). Most sparids are characterized as carnivorous predators, and the presence of algae in the stomach contents of some species has been attributed to accidental ingestion (Randall 1968, Manooch 1977, Horvath et al. 1990). To date, members of the genus Calamus have been described as carnivorous predators that exhibit conical teeth in the anterior region of the mandibles, which they use to grab prey, and wide molars on the sides of the mandibles, which they use to grind food (Randall and Caldwell 1966, Randall 1967). Therefore, it is possible that the algal remains found in this study were accidently ingested by the fish when capturing their prey.
Species of the genus Calamus feed mainly on mobile benthic invertebrates, which agrees with what was found in this study. Reid (1954) reported that the diet of Calamus arctifrons comprised copepods, amphipods, mysids, shrimps, bivalves, and gastropods. Dubovitsky (1977) reported that the main food items of Calamus proridens were bivalves and small crabs, while its main prey in the Gulf of Mexico was the crustacean Penaeus durorarum. Sedberry (1989) analyzed the diet of Calamus leucosteus in South Carolina in the United States and reported that its main food source was gastropods, although decapods, polychaetes, pelecypods, echinoderms, and fishes were also consumed. A study of Calamus nodosus in North Carolina and South Carolina in the United States reported that this species is an opportunistic carnivore that preys on benthic organisms; its trophic spectrum comprised mostly mollusks, as well as crabs, polychaetes, echinoderms, cirripeds, shrimps, squids, nematodes, fishes, anemones, sea cucumbers, and octopuses (Horvath et al. 1990).
The histological analysis of gonads sampled in this study showed that the Pacific Porgy presents asynchronous development of the oocytes and partial spawning, and it can reproduce year-round in Santa Rosalía (Veranes-Dip et al. 2022). These characteristics are displayed by fishes that adjust oocyte recruitment based on the exogenous energy available during development and gamete spawning (McBride et al. 2015, Saborido-Rey 2016). Therefore, food resources are one of the most important factors for reproductive success (do Carmo et al. 2019).
Given that reproduction demands large amounts of energy, it can induce variability in feeding behavior in terms of intensity or the types of prey consumed. Predators may prefer more energetic prey during maturity and spawning (Ysla-Guzmán et al. 2021). Therefore, lipids are an important energy resource for fish reproduction (Johnson 2009). In particular, highly unsaturated fatty acids (HUFAs) have been linked to gonadal development, fecundity, fertilization, egg quality, and larval hatching rates (Fernández-Palacios et al. 1995, Johnson 2009).
The most important items in the diet of C. brachysomus were bivalves. This group of organisms is known for being an important source of lipids, as they concentrate HUFAs obtained from phytoplankton (Ricardo et al. 2015). This accumulation is even greater in the gonads of bivalves during maturity and spawning (Giese 1966). There are reports of several species in the families Veneridae (Baqueiro and Aldana 2000), Chionidae (Baqueiro and Aldana 2000, Castillo-Durán et al. 2013), Mytilidae (García-Domínguez et al. 2018), and Pectinidae (Baqueiro and Aldana 2000) that can reproduce year-round in the Gulf of California. Therefore, bivalves may be an energy source that is available year-round and that could provide the lipids necessary for reproduction, which would explain the absence of significant variations in diet during the reproductive stages analyzed in this study.
The PERMANOVA results indicated significant variations between seasons in the most important prey consumed. Seasonal variations in the trophic spectrum of other sparids have been reported by several authors (Pallaoro et al. 2003, Osman and Mahmud 2009). These variations could be related to abiotic conditions, as well as to natural fluctuations in the abundance and availability of prey species (Chakroun and Kartas 1987, Bock et al. 2010). There is a marked seasonality in the Gulf of California in terms of the SST gradient, as well as mesoscale oceanographic processes that can stimulate changes in community structure (Avendaño-Ibarra et al. 2013, Inda-Díaz et al. 2014, Contreras-Catala et al. 2015). The closeness of Santa Rosalía to productivity centers, such as the Midriff Islands of the Gulf of California (Martínez-Zavala et al. 2010), could influence the abundance of prey species. Therefore, we consider that the Pacific Porgy is a predator with non-specific feeding habits that can adjust its diet to the most abundant and available food resources in its habitat.
The standardized Levin index for all data (overall) and data by category showed that C. brachysomus may be considered a predator with a narrow trophic niche. The Costello graphs suggested that there were only a few dominant prey species, and their importance varied. Some sparids, including species within the Calamus genus, break up food with their molars and pharyngeal teeth, which makes taxonomic identification difficult. Several of the prey items obtained in this study could not be identified to either species or genus level due to the advanced degree of digestion. However, we found a wide trophic spectrum that comprised 90 prey items, most of which were present in low abundance in the stomach contents.
The 5 most important prey species based on %PSIRI represented 38% of all diet items. There were also significant differences in diet by season. Thus, we can infer that although the Levin index indicated that the feeding strategy was that of a specialist predator, C. brachysomus could be categorized as an opportunistic predator that takes advantage of the availability and abundance of prey in the moment, which can be corroborated by the results reported for other sparid species (Ghanbarzadeh et al. 2014, Goldman et al. 2015, Jin et al. 2023). The trophic level calculated for the Pacific Porgy was 3.18. This classifies it as a tertiary consumer, making it an important link between lower and upper trophic levels. This coincides with what has been reported for other sparid species (Jin et al. 2023). The trophic level of the Pacific Porgy indicates that it is an important species for energy transfer, as are serranids and lutjanids (Moreno-Sánchez et al. 2019, Quiroga-Samaniego et al. 2022).
CONCLUSIONS
Calamus brachysomus is an opportunistic carnivorous predator that consumes mostly benthic prey. It has non-specific feeding habits and adjusts its diet based on the most abundant and available prey in its habitat. The trophic spectrum of this species was not altered by reproductive events, and the energy acquired from prey during each developmental stage is sufficient for recruitment and spawning without these fish needing to consume more energy-rich prey.

