










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de fitopatología
versión On-line ISSN 2007-8080versión impresa ISSN 0185-3309
Rev. mex. fitopatol vol.44 no.2 Texcoco may. 2026 Epub 15-Jun-2026
https://doi.org/10.18781/r.mex.fit.2602-3
Phytopathological Notes
Detection of a tentative alphanucleorhabdovirus infecting Carica papaya in Costa Rica
1Centro de Investigación en Biología Celular y Molecular (CIBCM), Universidad de Costa Rica, San José 11501-2060, Costa Rica.
2Escuela de Agronomía, Universidad de Costa Rica, San José 11501-2060, Costa Rica.
3 Instituto Nacional de Innovación y Transferencia en Tecnología Agropecuaria, Centro de Innovación Agropecuaria Los Diamantes, Guápiles, Pococí, Limón, Costa Rica.
Background/Objective.
Foliar chlorosis, short internodes, and curved petioles with purple streaks were observed in papaya (Carica papaya) crops (North and Atlantic regions) in Costa Rica since 2014. Identification of a putative plant virus associated with these symptoms was the aim of this research.
Experimental development.
Plant material was tested by ELISA (four plant viruses and potyviruses group), transmission electron microscopy (TEM), RT-PCR (degenerate primers for plant viruses), sequencing and phylogenetics.
Results
. All ELISA tests resulted negative. Bullet-shaped particles inside nuclei, and reticulum endoplasmic were only observed by TEM in symptomatic plants. Amplicons of 900 bp were consistently obtained from symptomatic samples using degenerate primers for plant rhabdoviruses. Nucleotide sequences showed 95.6 and 96.8% similarity to a putative papaya alphanucleorhabdovirus (Alphanucleorhabdovirus, Rhabdoviridae).
Conclusion.
This is the first report of a putative alphanucleorhabdovirus associated with symptomatic papaya plants showing streaked petiole (“pecíolo rayado”) disease in Costa Rica, but Koch’s postulates must be fulfilled and vector identified.
Keywords: Plant rhabdoviruses; “Streaked petiole” disease; Papaya necrosis apical
Antecedentes/Objetivo.
Clorosis foliar, entrenudos cortos, y pecíolos curveados con estrías color púrpura se observan en plantaciones de papaya (Carica papaya) en las regiones Norte y Atlántica en Costa Rica desde el 2014. La identificación de un presunto virus de plantas asociado con estos síntomas fue el objetivo de esta investigación.
Desarrollo experimental.
El tejido vegetal fue evaluado mediante ELISA (cuatro virus de plantas y el género potyvirus), microscopia electrónica de transmisión (MET), RT-PCR (con iniciadores universales para virus de plantas), secuenciación y análisis filogenéticos.
Resultados
. Todas las pruebas de ELISA resultaron negativas. Mediante MET, se observaron sólo en las plantas sintomáticas, partículas virales con forma de bala, éstas dentro de membranas en los núcleos y el citoplasma. Amplicones de 900 pb se obtuvieron consistentemente en las muestras sintomáticas usando iniciadores degenerados para rhabdovirus de plantas. Las secuencias nucleotídicas mostraron similitudes entre 95.6 y
96.8 % con un alphanucleorhabdovirus (Alphanucleorhabdovirus, Rhabdoviridae) tentativo que infecta papaya.
Conclusión.
Este es el primer reporte de un alphanucleorhabdovirus tentativo asociado con plantas sintomáticas de papaya mostrando la enfermedad del “pecíolo rayado”, como se le conoce localmente en Costa Rica, pero los postulados de Koch deben ser cumplidos y el vector identificado.
Palabras clave: Rhabdovirus de plantas; Enfermedad del “pecíolo rayado”; Necrosis apical de la papaya
Introduction
Papaya (Carica papaya) production in Costa Rica represents a high-value economic and social activity with an average annual growth of 5% between 2018 and 2022 (Acuña, 2023). The main production areas are found in the Atlantic, Central Pacific and northern regions of the country. About 75% of papaya production is sold locally, and the remainder is exported. During 2022, exports were done to a total of 10 destinations, where North American market (Canada and USA) represented 94% of shipments, while the European market was 4% of Costa Rica's total exports (Acuña, 2023).
This crop is affected by several diseases caused by plant viruses (Quito-Avila et al., 2023) and phytoplasmas (Fiore et al., 2018), causing significant losses worldwide. Additionally, in some countries in the Caribbean region, an organism similar to rickettsia causes “papaya bunchy top” disease (PBT) (Davis et al., 1998). Farmers (mainly in developing countries) neither keep records of crop yield and losses, nor their association with diseases or specific symptoms on the plants. It is very common that farmers coexist with symptoms of disease until the problem is ubiquitous and challenges the plants’ survival. Since 2014, papaya growers in Costa Rica have reported a novel disorder locally designated as ‘pecíolo rayado’ (streaked petiole). This condition affects several commercial varieties, including Maradol, Pococí, and Tainung, as well as wild hybrids. Symptomatology is characterized by curved petioles exhibiting distinctive purple-colored streaks, accompanied by foliar chlorosis and laminar wrinkling. Progressed infections result in shortened internodes and significant fruit set inhibition, posing a potential threat to local production. Those symptoms were observed in commercial orchards of papaya in North and Atlantic regions of Costa Rica. The objective of the work reported herein was to determine if a plant virus is associated to symptoms of streaked petiole (“pecíolo rayado”) disease in papaya plants in Costa Rica.
Experimental development
Leaf samples of 70 mature papaya plants (8-10 months) showing at least two symptoms associated to “streaked petiole” disease, including leaf petiole curved, reduced number of leaves in the apical meristem, cessation of fruit setting, and purple-colored streaks in petiole, were collected (ending 2014) from Muelle de San Carlos (Alajuela province), and from Guácimo and Guápiles (Limón province) in Costa Rica. Selected papaya orchards were free of mineral deficiencies and showed no signs of soil flooding or waterlogging. Samples collected were analyzed serologically by ELISA tests to detect the viral species Potyvirus papayanuli (papaya ringspot virus, PRSV), Potexvirus papayae (papaya mosaic virus, PapMV), Orthotospovirus tomatomaculae (tomato spotted wilt virus, TSWV), Cucumovirus CMV (cucumber mosaic virus, CMV) and with a genus- specific antibody against potyvirus group. The specific antibodies and positive controls to each virus were both supplied by Agdia Inc. (Elkhart, IN). Tests were carried out according to protocols and recommendations from Agdia Inc.
Due to the negative results obtained in ELISA tests, transmission electron microscopy (TEM) was performed to visualize potential phytopathogens. A subset of eight foliar samples of symptomatic papaya plants (Muelle n=4, Guácimo n=2 and Guápiles n=2), and a healthy plant from a garden in the Central Valley, all of them negative to viruses tested by ELISA, were collected (January, 2015) and processed for TEM, according to Montero-Astúa et al. (2017). Lamina and petiole sections were fixed overnight using Karnovsky solution buffered in cacodylate 0.05 M at 15 °C, and post-fixed with osmium tetroxide (1%) at room temperature after four repetitions of a washing step. Samples were dehydrated and then embedded by using an ethanol/propylene oxide series, followed by a mix of propylene oxide: epoxy resin with Spurr´s medium-hard, followed by embedding in pure epoxy resin, and finally, sample polymerization by heating. Thin sections were double stained (aqueous uranyl acetate and Sato’s lead citrate) and observed using a Hitachi H- 7100 electron microscope (Tokyo, Japan) at 100kV. Ten pictures were selected to measure diameter and length of 100 viral particles, using Image J software (Abramoff et al., 2004) to obtain the average of both measures.
Total RNA was extracted using RNeasy Plant Mini Kit (Qiagen, Germany) from those eight symptomatic plants tested by TEM, and two healthy plants. Reverse transcription was done with Maxima H Minus First Strand cDNA Synthesis Kit (Thermo Scientific, Lithuania) primed with random hexamers, according to the suppliers’ recommendation. To determine if bullet-shaped particles found in symptomatic leaves of papaya by TEM, belong to a putative nucleorhabdovirus or dichorhabdovirus, the cDNAs were tested for both plant viruses´ genera by end-point PCR using a pair of degenerated primers for plant rhabdoviruses (Rhab-R / Rhab-F) developed by Lamprecht et al. (2009) to amplified the L (Large) gene that encodes the viral RNA-dependent RNA polymerase (RdRp), and to the dichorhaviruses OFV and CiLV-N (LdF / LdR), designed by Ramos - González et al. (2016) to amplify a conserved region of the L gene (Table 1). Afterwards, a second group of 24 papaya foliar samples (collected in June, 2015) was also evaluated using primer pair Rhab-R / Rhab-F.
Twelve amplicons obtained with primer pair Rhab-R / Rhab-F (Lamprecht et al., 2009) were purified and directly sequenced (Macrogen, Korea) in both directions with the same primer pair used in the PCR. The contig sequence corresponding to each amplicon was obtained using BioEdit v7.7.1 (Hall 1999), and a similarity of sequence search was done using BLASTn algorithm at the GenBank (National Center for Biotechnology Information, USA). Four sequences obtained for a putative alphanucleorhabdovirus were deposited in GenBank (PX637683, PX637684, PX637685, and PX637686).
During 2015-2016, additionally, 193 foliar samples (126 symptomatic plants) were collected from fields in Muelle, Guácimo, and Guápiles and analyzed following the RT- PCR protocol to detect plant rhabdoviruses described above.
Table 1 Oligonucleotides (primers) and thermocycling profiles used to detect RNA plant viruses in Carica papaya plants analyzed in this study.
| Primer name | Oligonucleotide sequence (reference) | Primers pair (expected PCR product size), thermocycle profile(reference) |
|---|---|---|
| Plant rhabdoviruses | ||
| Rhab-R | 5’GTCCABCCYTTTTGYC3´ | Rhab-R/ Rhab-F (900 pb, Lamprecht et al., 2009) |
| Rhab-F | 5’GGATMTGGGGBCATCC3’ | 94 ºC for 2 min; 36 cycles (94 ºC - 30 s; 37 ºC - 30 s; 72 ºC - 90 s); final extension: 72 °C for 7 min (this study) |
| Rhab-R / Rhab-F (Lamprecht et al., 2009) | ||
| Dichorhavirus orchidaceae (Orchid fleck virus) and | ||
| Dichorhavirus leprosis (Citrus leprosis virus N) | ||
| LdF | 5’CCYGTGAGAGAATTCYTGGATG3’ | LdR/ LdF (390-400 pb) |
| LdR | 5’CCAGATTGGTGTARCCRAACAG3’ | 95 °C for 5 min, 35 cycles of (95 °C for 30 s, 55 °C for 30 s), final extension: 72 °C for 30 s. (Ramos - González et al., 2016) |
| LdF / LdR (Ramos - González et al., 2016) | ||
To determine the phylogenetic relationship between the observed bullet-shaped plant virus, a phylogenetic analysis was run with four sequences obtained herein (PX637683 to PX637686), including for comparison 31 different plant rhabdoviruses (total of 34 sequences due to repeated species) and Farmington virus (CT114) as outgroup; all of them retrieved from GenBank (https://www.ncbi.nlm.nih.gov/genbank). There were a total of 1 099 positions in the final dataset. The 39 sequences were aligned using ClustalW algorithm in BioEdit and then analyzed using maximum likelihood method (General Time Reversible model and rate variation across sites using a gamma distribution with invariable sites and 1 000 permutations) in MEGA 12 (Kumar et al., 2024).
Papaya plants exhibiting evident arrested fruit setting (Figure 1A and 1B), curved petioles (Figure 1B and 1E), purple-colored streaks in petioles (Figure 1D); light chlorosis and wrinkling of the leaf lamina, short internodes and reduced fruit petioles, were frequently observed (50 -70% incidence) in commercial orchards of papaya in North and Atlantic regions of Costa Rica. Additional symptoms were clearly observed depending on the plant growth stage, such as fewer flowers and fruits per plant; however, these were not associated with fruit abortion. Loss of fruit production is estimated at 40-50% in the crop, due to the irreversible cessation of fruit set in diseased plants and their eventual dieback. When measuring the angle of insertion of peduncles of flowers and fruits, starting from the apex of the trunk, the infected plants presented a smaller angle of insertion. Diseased plants had fewer leaves in the crown (Figure 1A and 1B), and some of these showed lamina with marginal necrosis. Loss of leaves and necrosis of apical new leaves caused plants to resemble a standing point-up pencil in the field, similar with the final step symptom of plants with PBT. However, latex was present in different tissues, including fruits, contrary to PBT typical symptom (Davis et al., 1996).
Different ELISA tests done (PRSV, PapMV, CMV, TSWV, and potyviruses group) to those symptomatic plants showed negative results for the 70 papaya symptomatic samples (Table 2). Only positive controls, used to each tested virus, exhibited color change of substrate solution after incubation time according Agdia Inc.´s protocols (data not shown).
Table 2 Summary of tests performed and results obtained from foliar samples of symptomatic and asymptomatic Carica papaya plants collected in Costa Rica to determine whether any plant virus is associated with “streaked petiole” disease.
| Region | ELISA | MET | RT-PCR | |||
|---|---|---|---|---|---|---|
| positive/ n tested) | positive/ n tested) | positive/ n tested) | ||||
| S | A | S | A | Sz | A | |
| North | 0/35 | 0/4 | 4/4 | -- | 60/61 | 0/28 |
| Atlantic, Guapiles | 0/18 | 0/3 | 2/2 | -- | 48/50 | 1/18 |
| Atlantic, Guacimo | 0/17 | 0/3 | 2/2 | -- | 42/47 | 0/21 |
| Central Valley | -- | -- | -- | 0/1 | -- | 0/2 |
| Total | 0/70 | 0/10 | 8/8 | 0/1 | 150/158 | 1/69 |
S or A= Samples from symptomatic (S) or asymptomatic (A) plants; ELISA: tests carried out to: Potyvirus papayanuli, Potexvirus papayae, Orthotospovirus tomatomaculae, Cucumovirus CMV, and Potyvirus group; TEM: presence of bullet-shaped particles into the nuclei; RT-PCR: amplicon of 900 bp obtained; z: samples tested included the 32 (8 and 24) symptomatic plants collected to validation tests using primers Rhab-R /Rhab-F; --: Not tested

Figure 1 Morphological alterations observed in Carica papaya plants with “streaked petiole” disease in Costa Rica. A) A symptomatic plant (arrow) exhibiting downward-curved petioles, leaf reduction in the apical meristem, and cessation of fruit set. Adjacent (slightly behind) two healthy plants (right) showing prolific fruit development and typical foliar architecture; B) Detailed view of upper part of a symptomatic plant showing leaf reduction in the apical meristem, leaf petiole curved, and poor fruit setting, behind (left) a healthy plant with normal fruit production; C) Detailed view of upper part of a healthy plant; D) Petioles of a diseased plant displaying purple-colored streaks; E) Comparison between petioles from symptomatic (top) and healthy (bottom) plants.
Presence of particles of a plant virus in symptomatic papaya plants was showed by TEM results. Numerous bullet-shaped particles were observed inside distended nuclei (Figure 2A and 2C) and also budding through the perinuclear space (Figure 2B), and into vesicles scattered in the cytoplasm of cells, but only from tissue of diseased plants. The average diameter and length of the viral particles (n=100) were 70 nm and 260 nm, respectively. No other viral inclusion or viral particles were detected in the samples.
Preliminary RT-PCR analysis of eight symptomatic samples for the presence of plant rhabdoviruses and/or dichorhaviruses yielded amplicons of the expected size (900 bp) only with the primer pair specific for plant rhabdoviruses (Rhab-R/Rhab-F); no amplification was observed in healthy plants. Subsequently, an additional 24 symptomatic plants from a second sampling consistently produced a 900 bp amplicon corresponding to rhabdoviruses, whereas no amplification was detected in healthy papaya samples or in the negative control. Therefore, a plant rhabdovirus was associated with symptomatic plants: initially in a batch of eight samples, then in a batch of 24 samples, and finally in 95% (150/158) of symptomatic papaya samples collected in this study (2015-2016). In contrast, only 1.4% (1/69) of asymptomatic samples tested positive (Table 2).
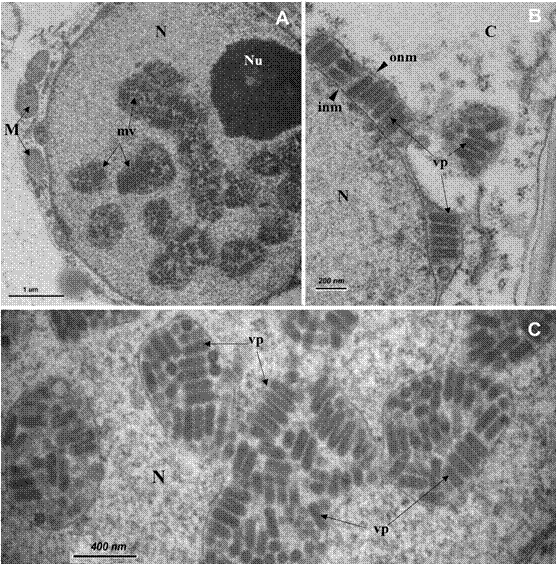
Figure 2 Nucleus of a leaf parenchyma cell of Carica papaya symptomatic plant with “streaked petiole” disease observed with transmission electron microscopy. A) Bullet-shaped particles inside membranous vesicles in the nucleus; B) Detail of particles seen into the perinuclear space; C) Detail of viral particles into the packages scattered in the nucleus. C= cytoplasm, M= mitochondria, N= nucleus, Nu= nucleolus, inm= inner nuclear membrane, mv= membranous vesicle filled of viral particles, onm= outer nuclear membrane, vp= viral particles.
The nucleotide sequences (Sanger) from 12 of the stronger amplicons obtained, confirmed association with a plant virus related to alphanucleorhabdoviruses available at GenBank. The BLASTn hits to these resulting sequences indicate 95.6 and 96.8% similarity to papaya alphanucleorhabdovirus 1 (PNRV-1), GenBank Acc. Nos. MN203193 and MN203194, respectively. The phylogenetic analysis done to the Costa Rican strain related to PNRV-1 showed that both viruses were grouped in the main cluster containing all nucleorhabdoviruses included, and into the alphanucleorhabdoviruses clade (Figure 3).
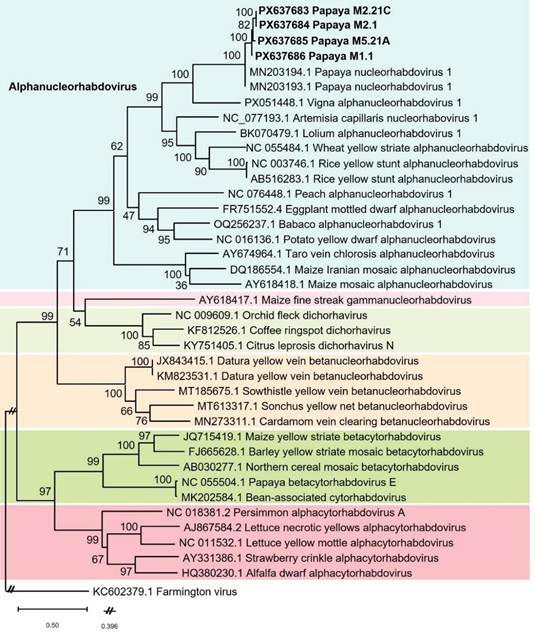
Figure 3 Phylogenetic tree constructed with partial sequences (900 nt) of the viral RNA-dependent RNA polymerase (L gene) of a putative alphanucleorhabdovirus obtained from Carica papaya of Costa Rica (GB Acc. No. PX637683 to PX637686) with 31 different plant rhabdoviruses (34 sequences) and Farmington virus (GB Acc. No. KC602379) as outgroup, all of them retrieved from GenBank. These were aligned with ClustalW algorithm using BioEdit v.7.7.1, and phylogeny inferred with the maximum likelihood method in Mega 12 (General Time Reversible model with a gamma distributed rate of substitution with invariant sites) and a bootstrap of 1 000 replicates.
Similar symptoms to the ones reported herein in Costa Rica, were described in papaya orchards in other countries in the Caribbean Basin, about 45 years ago. Lastra and Quintero (1981) informed the papaya apical necrosis in Venezuela (Zulia state), initially plants showed general wrinkling of leaves, followed by chlorosis of the younger leaves, wilting of those already developed, falling of leaves, and necrosis, leading to the death of the plant. Wan and Conover (1981) also described papaya droopy necrosis disease in Florida-USA (Dade, Monroe, Sarasota counties), and detailed the list of symptoms associated, as re-curvature of the leaves, arching of the petioles downward, chlorosis of new leaves, petioles shorter and stiffer with gray streaks and somewhat corrugated, short internodes (generating a crown rounded with racemose appearance), with short and rigid staminate inflorescences, abortion of pistillate flowers, fruiting setting stops, without fruit abortion. Latex presence when cutting fruits or plants. The last phase in the disease progression presented abscission of leaves, marginal necrosis of new leaves and descending necrosis of the stem. In addition to these reports, symptoms of apical necrosis of papaya were reported in Cuba, La Havana, Holguin and Villa Clara provinces (Mejías et al., 1987; Hernández-Pérez et al., 1988), and Mexico (Oaxaca, Veracruz and Tabasco) (Becerra et al., 1999).
Despite the viruses PRSV and PapMV are two frequent phytopathogens infecting papaya crops in many countries worldwide, both as well as other viruses tested were not detected in the samples collected in North and Atlantic growing areas from Costa Rica. In this country, the PRSV-W strain has been detected infecting cucurbits (Rivera et al., 1993) and other plant hosts (Sanchez et al., 1998), but not the PRSV-P strain. Likewise, PapMV has not been detected in the country. Also, CMV, TSWV and some potyviruses have been detected infecting different plant hosts but not papaya (Garita et al., unpublished).
The bullet-like particles observed in Costa Rican papaya samples, and their localization inside cellular nuclei (Figure 2) resembled bacilliform rhabdoviruses infecting plants (Jackson et al., 2005). Distended nuclei containing bullet particles were similar to previous reports, in symptomatic papaya samples, from Venezuela (Lastra and Quintero, 1981), Florida-USA (Wan and Conover, 1981), Cuba (Mejías et al., 1987) and Mexico (Becerra et al., 1999).
Plant rhabdoviruses (Rhabdoviridae family) have negative-sense single-stranded RNA genomes, infect a wide range of hosts, including monocots and dicots plant species, and cause important crop diseases. Traditionally, these rhabdoviruses were divided into cytorhabdoviruses and nucleorhabdoviruses, based on genome organization, phylogeny, and intracellular replication site. Nucleorhabdovirus genus was recently divided into genera: Alphanucleorhabdovirus, Betanucleorhabdovirus, Gammanucleorhabdovirus, and Deltanucleorhabdovirus (Simmonds et al., 2024; Walker et al., 2022; Dietzgen et al., 2020). High-throughput sequencing (HTS) technology, also known as next generation sequencing (NGS), has allowed increasing the number of plant viruses into these four genera, as well as their plant hosts (Bejerman et al., 2021, 2025).
Associations between papaya diseases and rhabdovirus-like particles were reported decades before molecular tools were available. But more recently, HTS has enabled genomic characterization of papaya-infecting rhabdoviruses, allowing clearer taxonomic placement and epidemiological study. A prevalent papaya-infecting cytorhabdovirus present in commercial orchards in Ecuador, was characterized and named papaya virus E (PpVE) by Medina-Salguero et al. (2019), and later designated Cytorhabdovirus papayae (Bejerman and Dietzgen 2019).
Also, Papaya nucleorhabdovirus 1 (PNRV-1) is a putative alphanucleorhabdovirus found by metagenomics analysis of papaya samples from Chiapas, Mexico (GenBank Acc. No. MN203193- MN203195). This rhabdovirus was detected in mixed infections with other viruses, including both PRSV-symptomatic and non PRSV-symptomatic plants (Alcalá-Briseño et al., 2020). Another putative nucleorhabdovirus (babaco nucleorhabdovirus-1, BabRV-1, GenBank Acc. No. OQ256237) has been recently detected infecting babaco (Vasconcellea × heilbornii) by metagenomics in Ecuador (Reyes-Proaño et al., 2023), but it may not be related to those found in Mexico and Costa Rica, the comparison of PNRV-1 and BabRV-1 did not show a significant similarity according to BLAST algorithm. Following Alphanucleorhabdovirus demarcation criteria of the International Committee on Taxonomy of Viruses (ICTV), which consider complete genome identity (threshold <75%) and differences in ecological niches, including hosts and vectors, the high nucleotide similarity (95.6-96.8%) to PNRV-1 of the L gene partial segment herein obtained, along with their shared host (C. papaya), suggests these may be the same virus. However, obtaining the complete L gene sequence or, ideally, the full genome of the Costa Rican isolate is necessary to definitively confirm its identity.
The phylogenetic analysis performed with the Costa Rican strain showed that Mexican (PNRV-1) and Costa Rican strains are related and grouped in the main cluster containing alphanucleorhabdoviruses (Figure 3). Unfortunately, sequences of putative nucleorhabdoviruses previously detected by TEM in papaya in Venezuela, Florida (USA), Mexico or Cuba are not available at GenBank for comparison purposes. However, the symptoms described in those reports and associated to rhabdovirus-like particles (bullet- shaped), suggest that those reports and the nucleorhabdovirus reported herein from Costa Rica may represent an old phytopathogen of papaya in this geographical region. Sequencing samples from the different countries where those symptoms have been reported would contribute to do an appropriate analysis.
This report and the ones previously done in the region warrant additional studies. It is necessary to fulfill Koch´s postulates in order to determine the relationships of the putative alphanucleorhabdoviruses detected in papaya with the observed symptomatology. Besides, studying associations with other viruses infecting papaya, identification of alternative hosts and detection of putative insect vectors would help to understand aspects related to ecology and epidemiology of this disease and to design appropriate management strategies to help papaya growers.
Conclusions
Results, obtained by TEM, showed that C. papaya affected by streaked petiole (“pecíolo rayado”) disease harbor particles similar to nucleorhabdoviruses, and accordingly, the obtained nucleotide sequences indicate a virus closely related to papaya nucleorhabdovirus 1 (PNRV-1), a putative alphanucleorhabdovirus found using NGS by Alcalá-Briseño et al. (2020). Our speculation, as previously mentioned, is that this plant virus may be related to detections by TEM in papaya in Venezuela and Florida (USA) in the early 1980s, and later in Cuba and Mexico. However, sequencing work must be done in symptomatic papaya plants from countries at the Caribbean Basin, to find evidence to support this suggestion.
To our best understanding, this is the first report of an alphanucleorhabdovirus infecting papaya in Costa Rica. This was found associated to an emerging viral disease with negative effect on the crop, but additional studies must be done to determine how the virus is participating in the etiology of the disease and the vector(s) species associated.
Limitations. Budget constraints and lack of knowledge of the identity of the insect vector (hypothesized to be a cicadellid) represented limitations to complete Koch´s postulates. Moreover, plant rhabdoviruses are very difficult to transmit mechanically, needing techniques such as vascular puncture inoculation which demand technical expertise and specialized tools (Redinbaugh et al. 2001). It is also necessary i) to test a larger number of samples for the different viruses recently associated with papaya diseases in Latin America in order to have the complete panorama of the Costa Rican status, ii) search for alternative host plants, and iii) identify the virus vector(s). This information is needed to propose management recommendations and strategies to help the papaya growers. Therefore, further research is warranted to solve several unanswered questions.
On the other hand, using HTS technology to obtain the complete genome or longer sequences of the alphanucleorhabdovirus consistently detected in papaya symptomatic samples in Costa Rica is essential to enable an appropriate comparison with the Papaya nucleorhabdovirus 1 reported from Mexico (Alcalá-Briseño et al., 2020).
Conflicts interest. Authors declared no conflict interest exists concerning this research.
Funding. This work was supported by research grants FITTACORI (F24-14 and F17-16) and Universidad de Costa Rica (B4-509 and A1-801).
Acknowledgments. Authors greatly appreciate: i. the support of Costa-Rican papaya growers, PITTA PAPAYA, FITTACORI, and Universidad de Costa Rica; ii. the kindness of researchers: Ramón Lastra (Venezuela), Enrique Becerra (Mexico), and Ricardo Hernández-Pérez (Cuba), sharing their observations regarding this papaya disease in their countries.
Authors contribution. All authors contributed to the design of the study, sample collections and processing, and data analysis. All them reviewed and edited the manuscript and approved the final version.
Referencias
Abràmoff MD, Magalhães PJ y Ram SJ 2004. Image processing with ImageJ. Biophotonics International, 11: 36-42. [ Links ]
Acuña A 2023. Papaya: mercado global y las potenciales oportunidades para Costa Rica. https://www.descubre.cr/wp-content/uploads/2025/04/Mercado-global-y-las-potenciales-oportunidades-para-Costa-Rica-para-papaya.pdf. Consultado 3 Mayo, 2025. [ Links ]
Alcalá-Briseño RI, Casarrubias-Castillo K, López-Ley D, Garrett KA y Silva-Rosales L 2020. Network analysis of the papaya orchard virome from two agroecological regions of Chiapas, Mexico. mSystems, 5: e00423-19. 10.1128/mSystems.00423-19 [ Links ]
Becerra EN, Cárdenas E, Lozoya H y Mosqueda R 1999. Rhabdovirus en papayo (Carica papayaL.) en el sureste de México. Agronomía Mesoamericana, 10: 85-90. https://www.mag.go.cr/rev_meso/v10n02_085.pdf [ Links ]
Bejerman N y Dietzgen RG 2019. Letter to the editor: Bean-associated cytorhabdovirus and papaya cytorhabdovirus are strains of the same virus. Viruses, 11: 230. 10.3390/v11030230 [ Links ]
Bejerman N, Dietzgen RG y Debat H 2021. Illuminating the plant rhabdovirus landscape through metatranscriptomics data. Viruses, 13: 1304. 10.3390/v13071304 [ Links ]
Bejerman N, Dietzgen RG y Debat H 2025. Expanding the known nucleorhabdovirus world: the final chapter in a trilogy exploring the hidden diversity of plant-associated rhabdoviruses. bioRxiv. 10.1101/2025.05.09.653113 [ Links ]
Davis MJ, Kramer JB, Ferwerda FH y Brunner BR 1996. Association of a bacterium and not a phytoplasma with Papaya Bunchy Top Disease. Phytopathology, 86: 102-109. 10.1094/Phyto-86-102 [ Links ]
Davis MJ, Ying Z, Brunner BR, Pantoja A y Ferwerda FH 1998. Rickettsial relative associated with Papaya Bunchy Top Disease. Current Microbiology, 36: 80-84. 10.1007/s002849900283 [ Links ]
Dietzgen RG, Bejerman NE, Goodin MM, Higgins CM, Huot OB et al. 2020. Diversity and epidemiology of plant rhabdoviruses. Virus Research, 281: 197942. 10.1016/j.virusres.2020.197942 [ Links ]
Fiore N, Bertaccini A, Bianco PA, Cieślińska M, Ferretti L et al. 2018. Fruit Crop Phytoplasmas. En: Rao G, Bertaccini A, Fiore N, Liefting L. (eds) Phytoplasmas: Plant Pathogenic Bacteria - I. Springer, Singapore. 10.1007/978-981-13-0119-3_6 [ Links ]
Hall TA 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series, 41: 95-98. [ Links ]
Hernández-Pérez R, Suazo-Reynord M y Toledo P 1988. Necrosis apical de la frutabomba (Carica papayaL.), nueva enfermedad viral en la provincia de Villa Clara. Revista Centro Agrícola, 15: 63-67. [ Links ]
Jackson AO, Dietzgen RG, Goodin MM, Bragg JN y Deng M 2005. Biology of plant rhabdoviruses. Annual Review of Phytopathology, 43: 623-660. 10.1146/annurev.phyto.43.011205.141136 [ Links ]
Kumar S, Stecher G, Suleski M, Sanderford M, Sharma S y Tamura K 2024. MEGA12: Molecular Evolutionary Genetic Analysis version 12 for adaptive and green computing. Molecular Biology and Evolution, 41: 1-9. 10.1093/molbev/msae263 [ Links ]
Lamprecht RL, Pietersen G, Kasdorf GGF y Nel LH 2009. Characterization of a proposed nucleorhabdovirus new to South Africa. European Journal of Plant Pathology, 123: 105-123. 10.1007/s10658-008-9339-5 [ Links ]
Lastra R y Quintero E 1981. Papaya apical necrosis, a new disease associated with a rhabdovirus. Plant Disease, 65: 439-440. https://www.apsnet.org/publications/plantdisease/backissues/Documents/1981Articles/PlantDisease65n05_439.PDF [ Links ]
Medina-Salguero AX, Cornejo-Franco JF, Grinstead S, Mollov D, Mowery JD et al. 2019. Sequencing, genome analysis and prevalence of a cytorhabdovirus discovered inCarica papaya. PloS ONE, 14: e0215798. 10.1371/journal.pone.0215798 [ Links ]
Mejías Y, Rodríguez D y González G 1987. Rhabdovirus asociado a síntomas de necrosis apical en fruta bombaCarica papayaen Cuba. Ciencia y Técnica en Agricultura. Serie Protección de plantas, 10: 57-62. [ Links ]
Montero-Astúa M, Dejuk-Protti N, Vásquez E, Garita L y Moreira L 2017. First report of Iris yellow spot virus in Costa Rica. Australasian Plant Disease Notes, 12: 18. 10.1007/s13314-017-0243-8 [ Links ]
Quito-Avila DF, Reyes-Proaño EG, Cañada-Bautista M, Cornejo-Franco JF, Alvarez-Quinto RA et al. 2023. Papaya sticky disease caused by virus “couples”: a challenge for disease detection and management. Plant Disease, 107: 1649-1663. 10.1094/PDIS-11-22-2565-FE [ Links ]
Ramos-González P, Sarubbi-Orue H, Gonzales-Segnana L, Chabi-Jesus C, Freitas-Astúa J y Kitajima EW 2016. Orchid fleck virus infecting orchids in Paraguay: First Report and Use of Degenerate Primers for its Detection. Journal of Phytopathology, 164: 342-347. 10.1111/jph.12420 [ Links ]
Redinbaugh MG, Louie R, Ngwira P, Edema R, Gordon DT et al. 2001. Transmission of viral RNA and DNA to maize kernels by vascular puncture inoculation. Journal of Virological Methods, 98: 135-143. 10.1016/S0166-0934(01)00369-X [ Links ]
Reyes-Proaño EG, Cañada-Bautista M, Cornejo-Franco JF, Alvarez-Quinto RA, Mollov D et al. 2023. The virome of babaco (Vasconcellea × heilbornii) expands to include new members of theRhabdoviridaeandBromoviridae. Viruses, 15: 1380. 10.3390/v15061380 [ Links ]
Rivera C, Villalobos W, Sánchez MV, Zumbado C y Rodríguez CM 1993. Identification and distribution of melon-infecting viruses and their vectors in two provinces of Costa Rica. Turrialba, 43: 210-215. https://repositorio.catie.ac.cr/handle/11554/9667 [ Links ]
Sánchez MV, Agüero R y Rivera C 1998. Plantas hospederas de los virus más importantes que infectan el melón,Cucumis melo(Cucurbitaceae) en Costa Rica. Revista de Biología Tropical, 46: 13-25. https://revistas.ucr.ac.cr/index.php/rbt/article/view/19348/19397 [ Links ]
Simmonds P, Adriaenssens EM, Lefkowitz EJ, Oksanen HM, Siddell SG et al. 2024. Changes to virus taxonomy and the ICTV statutes ratified by the International Committee on Taxonomy of Viruses. Archives of Virology, 169: 236. 10.1007/s00705-024-06143-y [ Links ]
Walker PJ, Freitas-Astúa J, Bejerman N, Blasdell KR, Breyta R et al. 2022. ICTV virus taxonomy profile:Rhabdoviridae2022. Journal of general Virology, 103: 001689. 10.1099/jgv.0.001689 [ Links ]
Wan Sh y Conover RA 1981. A rhabdovirus associated with a new disease of Florida papayas. Proceedings Florida State Horticulture Society, 94: 318-321. https://journals.flvc.org/fshs/article/view/96333/92343 [ Links ]
Received: February 11, 2026; Accepted: April 24, 2026
 Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
