










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de fitopatología
versión On-line ISSN 2007-8080versión impresa ISSN 0185-3309
Rev. mex. fitopatol vol.44 no.2 Texcoco may. 2026 Epub 15-Jun-2026
https://doi.org/10.18781/r.mex.fit.2601-2
Phytopathological Notes
Potential antagonist of native rhizobacteria of Parkinsonia aculeata on Fusarium spp. associated with native maize varieties from Bajío, Mexico
1Departamento de Ingenierías, Tecnológico Nacional de México campus Valle del Yaqui. Av. Tecnológico s/n, Bácum, C. P. 85276 Sonora, México.
2Tecnológico Nacional de México campus Pabellón de Arteaga, Carretera a la estación de Rincón Km 1, Pabellón de Arteaga, C. P. 20670, Aguascalientes, México.
3Tecnológico Nacional de México campus Roque, Carretera Apaseo el Alto-Jerécuaro Km.12, Celaya, C. P. 38525, Guanajuato, México.
4Universidad Autónoma de Sinaloa, Facultad de Agricultura del Valle del Fuerte, Juan José Ríos, C. P. 81110, Sinaloa, México.
Background/Objective.
The genus Fusarium comprises some of the most important phytopathogenic fungi affecting maize crops. Fusarium cause diseases such as root, stalk, and ear rot, which negatively impact the crop's agricultural productivity. In the search for agrobiotechnological alternatives for controlling this fungus, this study was aimed to evaluate the antagonistic potential of Parkinsonia aculeata rhizobacteria against Fusarium spp. strains associated with native maize varieties from the Bajío region of Mexico.
Experimental development.
Two strains of Fusarium spp. (MC-03 and MC-05), isolated from roots of native maize varieties from the Bajío region of Mexico, exhibiting Fusarium wilt symptoms, were used. These fungi were tested in vitro against nine rhizobacteria of P. aculeata: Enterobacter cloacae (BA1), Priestia megaterium (BA4 and BA-7B), Sinomonas halotolerans (BA10-B), Staphylococcus warneri (BP5), P. endophytica (BP6), Bacillus subtilis (TP1 and TP2), and S. hominis (TM6). Microscopic and macroscopic characterization of the fungal strains and biochemical analysis of the rhizobacteria were performed. Radial growth inhibition of the fungal isolates was determined by triplicate dual fungus-rhizobacteria confrontations. A completely randomized design was used, analyzing the data obtained in the STATISTICA software using an ANOVA based on a linear model of fixed effects and a mean comparison test by DMS (p>0.05).
Results.
With the exception of P. endophytica (BP6), all rhizobacteria exhibited enzymatic activity related to fungal antagonism mechanisms. B. subtilis (TP1) produced glucanases, lipases, and proteases. Microscopic and macroscopic characterization of the fungal strains indicated that they belong to Fusarium spp. In the rhizobacteria-MC-03 confrontation, the B. subtilis bacterial strains (TP1 and TP2) were statistically similar and achieved the greatest inhibition of mycelial growth (23%). With the fungal strain MC-05, P. endophytica (BP6) and S. hominis (TM6) were statistically superior to the other rhizobacteria in inhibiting mycelial growth (17%), followed by the two B. subtilis strains (TP1 and TP2), which inhibited 10% and 17%, respectively.
Conclusion.
There was a significant variability in the mycelial growth response of the fungi to rhizobacteria. B. subtilis (TP1 and TP2), S. hominis (TM6), and P. endophytica (BP6) exhibited an antagonistic effect, inhibiting the mycelial growth of Fusarium spp. strains by up to 23% compared to the absolute and commercial controls. This study establishes the preliminary scientific basis for obtaining a biofungicide with specific inhibitory capabilities against the fungi studied.
Keywords: Bacillus subtilis; Phytopathogenic fungi; Priestia endophytica; Staphylococcus hominis; Zea mays
Antecedentes/Objetivo.
El género Fusarium comprende algunos de los hongos fitopatógenos más importantes que afectan al cultivo de maíz, ya que ocasionan enfermedades como la pudrición de la raíz, tallo y mazorca, que afectan la productividad agrícola de este cultivo. En la búsqueda de alternativas agrobiotecnológicas para el control de este hongo, se desarrolló el presente estudio para evaluar el potencial antagonista de rizobacterias de Parkinsonia aculeata contra cepas de Fusarium spp. asociadas a maíces criollos del Bajío, México.
Desarrollo experimental.
Se emplearon dos cepas de Fusarium spp. (MC-03 y MC-05), aisladas de raíces de maíces nativos del Bajío, México con síntomas de Fusariosis. Estos hongos se confrontaron in vitro con nueve rizobacterias de P. aculeata: Enterobacter cloacae (BA1), Priestia megaterium (BA4 y BA-7B), Sinomonas halotolerans (BA10-B), BP5 (Staphylococcus warneri), BP6 (P. endophytica), Bacillus subtilis (TP1 y TP2) y S. hominis (TM6). Se realizó una caracterización micro-macroscópica de las cepas fúngicas y bioquímica para las rizobacterias. Se determinó la inhibición del crecimiento radial de los aislados fúngicos mediante confrontaciones duales hongo-rizobacterias por triplicado. Se empleó un diseño completamente aleatorizado, analizando los datos obtenidos en el software STATISTICA mediante una ANOVA basada en un modelo lineal de efectos fijos y una prueba comparación de medias por DMS (p>0.05).
Resultados.
A excepción de P. endophytica (BP6), todas las rizobacterias presentaron actividad enzimática relacionada con mecanismos de antagonismo fúngico. B. subtilis (TP1) produjo gluacanasas, lipasas y proteasas. La caracterización micro y macroscópica de las cepas fúngicas indicó que pertenecen a Fusarium spp. En la confrontación rizobacterias- MC-03, las cepas bacterianas de B. subtilis (TP1 y TP2) fueron estadísticamente iguales y lograron la mayor inhibición del crecimiento micelial (23 %). Con la cepa fúngica MC-05, P. endophytica (BP6) y S. hominis (TM6) fueron estadísticamente superiores al resto de las rizobacterias al momento inhibir el crecimiento micelial en 17 %, seguidas por las dos cepas de B. subtilis (TP1 y TP2) que inhibieron el 10 y 17 % respectivamente.
Conclusión.
Existió variabilidad significativa en la respuesta del crecimiento micelial de los hongos ante las rizobacterias. B. subtilis (TP1 y TP2), S. hominis (TM6) y P. endophytica (BP6), presentaron un efecto antagónico, ya que inhibieron el crecimiento micelial de las cepas de Fusarium spp. hasta en 23 % respecto al control absoluto y comercial. El presente estudio establece las bases científicas preliminares para la obtención de un biofungicida con capacidades específicas de inhibición de los hongos estudiados.
Palabras clave: Bacillus subtilis; Hongos fitopatógenos; Priestia endophytica; Staphylococcus hominis; Zea mays
Introduction
The Fusarium genus comprises some of the most important pathogens in agriculture from an economic point of view (Sumerell et al., 2010), which, along with its phytopathogenic behavior, synthesize highly toxic mycotoxins in its infection process that lead to health problems in humans and animals (Velarde et al., 2018). In the maize crop, it causes the rot of root, stem and ear. Fusarium verticillioides is probably the most common one in the world; however, other species such as F. subglutinans and F. proliferatum are related to the same symptomatology (Leyva-Madrigal et al., 2014) and have been reported in the Bajío region (Figueroa-Rivera et al., 2010). The rotting caused by the Fusarium species in maize can hardly be controlled using chemical products due to different factors, mainly the endophytic characteristic of the infection (Bacon et al., 2001). The current trend is the search for bioprotective or biocontrol agents, with the use of microorganisms that allow for the fighting of phytopathogenic diseases and present characteristics associated to the promotion of plant growth (Canchignia-Martínez et al., 2024, Bhattacharyya and Jha, 2012). Different studies have shown that the infections caused by Fusarium in maize can be fought with bacterial biocontrol agents (Bhargavi et al., 2024; Sánchez-Ceja et al., 2023; Ayala-Torres et al., 2023; Leyva-Madrigal et al., 2014).
In regard to this, Figueroa-López et al. (2016) analyzed a collection of bacteria to detect their potential activity against F. verticillioides. The assays that they carried out in planta showed that three Bacillus isolates (B. megaterium, B. cereus sensu lato and Bacillus spp.) displayed the greatest antagonistic activity, as well as a reduction of the disease. Castro del Ángel et al. (2021) evaluated the antifungal activity of two endophytic B. amyloliquefaciens isolates against F. verticillioides under laboratory and greenhouse conditions. The in vitro inhibition in dual crops of the phytopathogenic fungus was obtained in a range of 59 to 62%. The endophytic bacteria reduced the incidence and severity of F. verticillioides by over 80% in plants that received the treatment in regard to the control.
In recent years, our research group has focused on the search for agrobiotechnological alternatives for the management of pests and diseases in crops of interest in Mexico. A bank of rhizobacteria associated to P. aculeata under drougth and salinity has been developed, which allowed us to obtain a group of molecularly identified bacteria (Peñuelas-Rubio et al., 2024b) with a potential antagonist (Herrera-Sepúlveda et al., 2023) and promoting the maize plant development (Argentel-Martínez et al., 2025), which is an incentive for the analysis of the microbial biocontrol capacities of these species. Due to this, our hypothesis is that, not only these rhizobacteria promote plant development, they can biocontrol the mycelial growth of Fusarium from their biochemical properties. Studies are also conducted on the isolation of fungal strains of the genus Fusarium from the roots of native maize grown under different irrigation regimes (Peñuelas-Rubio et al., 2024a). In this context, the aim of this investigation was to evaluate the potential antagonist of rhizobacteria against Fusarium spp. strains associated to native maize.
Experimental development
Collection of rhizobacteria and fungi. Nine antagonistic bacteria and two fungi associated to the roots of the maize crop that are a part of the Colección de Microorganismos Edáficos y Rizosféricos del Instituto Tecnológico del Valle del Yaqui (Collection of Edaphic and Rhizospheric Microorganisms of the Yaqui Valley Technological Institute) were used. The bacteria used were Enterobacter cloacae (BA1; ON869243), Priestia megaterium (BA4; ON869245 and BA7B; ON869246), Staphylococcus warneri (BP5; MH1982281.1), Priestia endophytica (BP6; MT373518.1), Bacillus subtilis (TP1; MN538531.1 and TP2; MN756672.1), Sinomonas halotolerans (BA10-B; ON869249) and Staphylococcus hominis (TM6; MT585539.1) isolated from the rhizosphere of Parkinsonia aculeata in two geographic regions of the Sonoran semi-desert (Peñuelas-Rubio et al., 2024). These rhizobacteria displayed antagonistic capacity against Fusarium strains isolated from tomato roots (data not published).
Biochemical characterization of the rhizobacteria. Biochemical hemolysis tests were carried out (Khalil et al., 2021), production of enzymes: chitinases (Roberts and Selitrennikoff, 1988), glucanases (Teather and Wood, 1982), proteases (Jones et al., 2007) and lipases (Salwoom et al., 2019; Stead, 1984), as well as plant growth promoting mechanisms (PGP): production of siderophores (Pérez-Miranda et al., 2007), indoleacetic acid (Gravel et al., 2007) and phosphorous solubilization (Edi-Premono et al., 1996; Pikovskaya, 1948). The PGP biochemical characterization of strains BA1 and BA7B was published in an earlier study (Argentel-Martinez et al., 2025).
Macro and microscopic characterization of the fungi. The fungal strains were isolated from the roots of Gavilán (Guanajuato) and Halcón (Michoacán) native maize that displayed symptoms (necrosis) associated to the presence of Fusarium spp. (Peñuelas-Rubio et al., 2024). Pathogenicity tests were performed using a rolled paper towel assay, reported by Leyva-Madrigal et al. (2014), where the strains presented a severity of 80 and 100% according to the scale by Figueroa-Rivera et al. (2010). In this study, a macro and microscopic characterization of these strains was carried out. For the evaluation of the macroscopic growth, pieces of fresh mycelium were planted (~5 mm in diameter) in the center of sterile Petri dishes containing PDA (Papa Dextrose Agar) medium. The dishes were incubated at 28 °C for 10 days under partial darkness conditions. At the end of the incubation period, the morphological characteristics of the growth were recorded, including texture, color, shape and the presence of pigmentation in both the front (top surface of the crop) and the back (base of the medium). For the observation of fungal structures, microcultures were performed by placing a piece of fresh mycelium in the center of a Petri dish containing freshly prepared PDA medium. On the inoculum, a sterile coverslip was carefully placed. The fungal cultures were incubated at 28 °C for 5 days under partial darkness conditions, enabling the development of reproductive structures adhered to the inner surface of the coverslip. After the incubation period, the coverslip was removed using sterile forceps and placed on a microscope slide with a drop of lactophenol blue as a mounting and staining medium. Observations were made under a compound light microscope (Primo Star, Zeiss, Germany) at 40 and 100X (with immersion oil), documenting the mycelial and conidial structures of each strain. For both cases, images were taken using a digital camera (MU-500, AmScope, United States) under standardized light conditions to ensure the comparability between strains (Riddell, 1950).
Dual confrontation bioassay. The methodology proposed by Khalil et al. (2021) was followed, in which the rhizobacteria were transferred to Luria Bertani (LB) agar and incubated at 30 °C for 24-48 h at 200 rpm. The bacterial suspension to inoculate was adjusted at 2x108 CFU mL-1. In the center of Petri dishes with SNA (Spezieller Nährstoffarmer Agar), a mycelial plug of the fungus (8 mm in diameter) was placed, and 2 cm away, 5 μL of the bacterial suspension were placed. The dishes were incubated for five days at 28 °C.
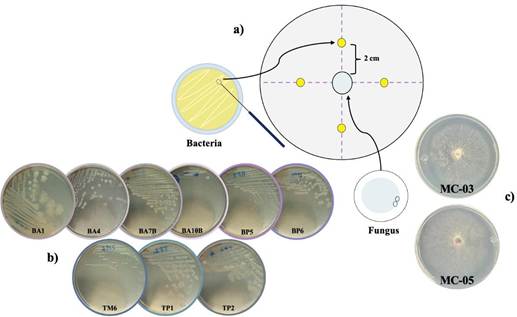
Figure 1 Diagram of the dual fungus-rhizobacteria confrontation assay. a) Distribution of microorganisms in the Petri dish; b-c) Macroscopic characteristics of the nine bacteria [E. cloacae (BA1), P. megaterium (BA4 and BA7B), S. halotolerans (BA10B), S. warneri (BP5), P. endophytica (BP6), S. hominis (TM6), B. subtilis (TP1 and TP2)] and two fungi (MC-03 and MC-05), respectively.
Experimental design and statistical analysis. A completely randomized design was used, in which the fungi (MC-03 and MC-05) were confronted with the nine rhizobacteria (BA1, BA4, BA7B, BP5, BP6, BA10B, TM6, TP1, TP2,) and two controls: the commercial control (CC), based on B. subtilis (Biosutil Bs at 2 L Ha-1; FIASA) and an absolute control (CA; fungus only). Each dual confrontation was performed in triplicate. Mycelial growth was determined after 5 days of incubation with the radial measurement of the fungus (cm) using a millimetric ruler, determining the distance between the fungal and the bacterial growth. A homogeneity and normality test were carried out on the data obtained from the mycelial growth. Subsequently, ANOVAs were performed, based on a linear fixed effects model. When statistically significant differences were presented between the means of the fungal mycelial growths due to the effect of the treatments (rhizobacteria), the DMS means multiple comparison test. To verify the efficiency of the mathematical model used, the unadjusted determination coefficient (R2) was calculated. The STATISTICA for Windows professional software, version 14.0, was used for these analyses.
According to Table 1, with the exception of BP6, all rhizobacteria presented enzymatic activity; no rhizobacteria were positive to the chitinase test. B. subtilis (TP1 and TP2) presented the ability to synthesize glucanases and P. megaterium (BA4) produce proteases. S. warneri (BP5), S. hominis (TM6) and B. subtilis (TP1) were the most efficient in the production of lipases. Regarding the plant growth promotion (PGP) mechanisms, E. cloacae (BA1), S. halotolerans (BA10B) y B. subtilis (TP1 ad TP2) produce siderophores most efficiently. On the ability of auxins synthesis (indoleacetic acid), E. cloacae (BA1),
P. megaterium (BA4 and BA7B) and BA10B (S. halotolerans) were positive in Salkowski’s reaction, E. cloacae (BA1) presented the highest production of auxins, and was also the most efficient in the phosphorous solubilization test. In general terms, TP1 (B. subtilis) presented the highest efficiency in the production of enzymes related to fungal antagonism mechanisms. These bacterial species have been reported for their antagonistic potential (Sallam et al, 2024; Biedendieck et al., 2021).
Table 1 Biochemical characterization of rhizobacteria isolated from palo verde (Parkinsonia aculeata).
| Rhizobacteria | Enzymatic activity | Mechanisms of PPG | |||||
|---|---|---|---|---|---|---|---|
| Chitinases | Glucanases | Proteases | Lipases | Siderophores | IAA(µg mL-1) | PSI | |
| P. megaterium (BA4) | - | - | +++ | + | ++ | 3.00 | - |
| S. halotolerans (BA10B) | - | - | - | + | +++ | 4.59 | - |
| S. warneri (BP5) | - | - | - | +++ | + | - | 2.28 |
| P. endophytica (BP6) | - | - | - | - | - | - | 2.43 |
| S. hominis (TM6) | - | - | - | +++ | - | - | 2.71 |
| B. subtilis (TP1) | - | +++ | +++ | +++ | +++ | - | - |
| B. subtilis (TP2) | - | +++ | +++ | - | +++ | - | - |
Results are expressed as the diameter of the halos formed around the colonies. Symbols: + < 3 mm; ++ > 3 < 4 mm; +++ > 4 mm. IAA (indoleacetic acid), PSI (phosphorus solubilization index). All tests were performed in duplicate. IAA and PSI values are the means obtained.
According to the macro and microscopic of the fungal strains presented in Figure 2, the texture of the colonies was observed to be cotton-like, with aerial mycelia and the color after 10 days was intensely wine-colored (Figure 2A). As single-celled characteristics, conidial structures, typical of the Fusarium genus were observed (Figure 2B and C). For both fungi, macro and microconidia were found, as well as chlamydospores (SENASICA, 2020).
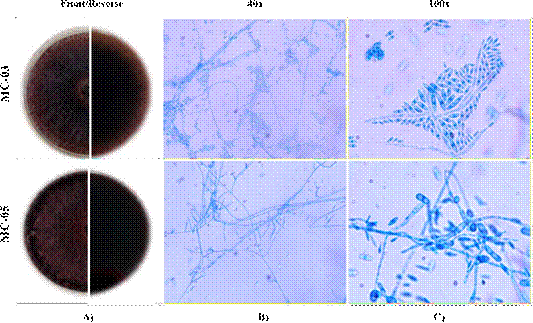
Figure 2 Morphological characterization of Fusarium fungal strains. A) Macroscopic characteristics of the fungal strains: front (top) and back (bottom) views of Petri dishes with PDA medium 10 days after sowing; B and C) Mycelial and conidial structures observed by optical microscopy at 40 and 100X (with immersion oil), from microculture at 5 days after sowing and stained with lactophenol blue (Riddell, 1950).
The results of the dual confrontation between strains MC-03 (Figure 3a) and MC-05 (Figure 3b), presented statistically significant differences (p<0.05) between the rhizobacteria evaluated regarding the mycelial growth of both fungi.
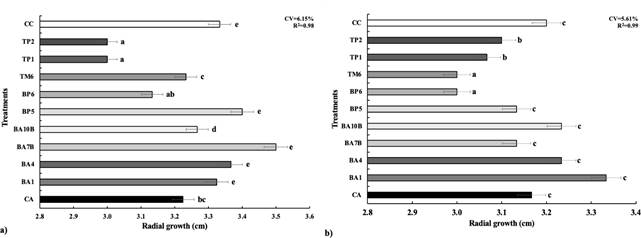
Figure 3 Mycelial growth exhibited by the Fusarium spp. strains MC-03 (a) and MC-05 (b) during dual confrontations with antagonistic bacteria. The treatments are shown on the Y-axis: CA: absolute control; CC: commercial control; antagonistic bacteria: BA1 (E. cloacae), BA4 and BA7B (P. megaterium), BA10B (Sinomonas halotolerans), BP5 (S. warneri), BP6 (P. endophytica), TM6 (S. hominis), TP1 and TP2 (B. subtilis).
In the confrontation between rhizobacteria and MC-03 (Figure 3a), the B. subtilis strains (TP1 and TP2) were statistically equal and they achieved the greatest inhibition of the mycelial growth (23%). With fungus MC-05, P. endophytica (BP6) and S. warneri (TM6) were statistically superior to the rest of the rhizobacteria, as they inhibited 17% of the mycelial growth, followed by the two B. subtilis strains (TP1 and TP2) that inhibited 10 and 7% respectively. Although P. endophytica (BP6) did not turn out positive for the biochemical antagonisms tests, it stands out for its potential in both confrontations, which suggests the possibility of this rhizobacteria being present in other Fusarium spp. inhibition mechanisms that were not considered in this study, such as the production of hydrogen cyanide (HCN) or cellulase, amylase and xylanase activity (Sharma et al., 2023). The analysis of the unadjusted determination coefficient led to infer that, due to the effect of the treatments, more than 98% of the contribution to the total variability of the fungi’s mycelial growth was obtained. Therefore, these average percentages of the evaluation for both fungi were the contribution of the linear fixed-effect mathematical model used to process the data (Figure 3).
In both confrontations, the rhizobacteria E. cloacae (BA1), P. megaterium (BA4) and S. halotolerans (BA10B), and the commercial control (Biosutil Bs), led to the growth of both fungal strains, possibly related to the ability to produce compounds that the fungus uses to grow (Pangihotan, 2025).
The control of phytopathogenic fungi using eco-friendly practices has developed based on the use of antagonistic microorganisms which, by using competitive mechanisms, antibiosis and the induction of resistance, limit their development. This analysis suggests that rhizobacteria have different degrees of effectiveness in the inhibition of Fusarium spp., possibly related to the production of specific secondary metabolites. This represents a potential antagonist related to the typical characteristics of the bacteria. Additionally, this study confirms that rhizobacteria present several characteristics related to the adaptation to semi-arid regions, the synthesis of phytohormones, the solubilization of phosphates and the synthesis of secondary metabolites (Peñuelas et al., 2024b). Finally, the assay shows the importance of exploring other metabolic pathways of the rhizobacteria; this approach would allow the understanding of its action mechanisms and potential for biological control. In addition, the need to confirm the species-level identification of the evaluated fungal strains is highlighted. For this purpose, it is crucial to characterize them using molecular markers and the fulfilment of Koch’s postulates. These considerations open up the possibility of future investigations that include a modification of the concentrations of the bacterial inoculants or the formation of consortia between the rhizobacteria. Such strategies can increase their antagonistic potential, improving their efficiency for Fusarium biocontrol.
Conclusions
Rhizobacteria B. subtilis (TP1 and TP2), TM6 (S. hominis) and BP6 (P. endophytica), presented an antagonistic effect, since they inhibited the mycelial growth of the Fusarium spp. strains by up to 23% in regard to the absolute and commercial control. The results prove the existence of significant response variability of the fungi against the different rhizobacteria. This study establishes the preliminary scientific foundations to obtain a biofungicide with specific capacities to inhibit the growth of the studied fungi.
Limitations. This study represents the basis for the selection of antagonistic bacteria that can be tested in planta in future assays.
Conflict of interest. The authors declare to have no conflict of interest in relation to this article.
Funding. The completion of this work was funded by the project “Native Rhizobacteria and Plant Extracts as Sustainable Alternatives for the Biocontrol of Fusarium Associated with the Planting of Maize (19252.24-P)” funded by the Tecnológico Nacional de México.
Acknowledgements. The authors thank the Tecnológico Nacional de México for the financial resources provided.
Contributions of the authors. Argentel-Martínez and Peñuelas-Rubio participated in the research, conceptualization, validation, project administration and acquiring funds. Writing, revision and editing: Argentel-Martínez and Peñuelas-Rubio. Peñuelas-Rubio, Fierro-Coronado and Romo-Rodríguez developed the investigation and methodology, Cervantes-Ortíz and Arias-Moscoso wrote and prepared the original draft. Writing: revision and editing was carried out by Peñuelas-Rubio. Argentel-Martínez carried out the writing, revision and formal analysis.
Referencias
Argentel-Martínez L, Peñuelas-Rubio O, Pérez López L, Arias Moscoso JL, Cadena Cadena F et al. 2025. An enhancement of growth and physiological performance in maize hybrids through a dual consortium of native rhizobacterial strains. Plant Growth Regulation, 105: 1559-1574. 10.1007/s10725-025-01354-y [ Links ]
Ayala-Torres AM, Aranda-Ocampo S, De León-García de Alba C, Nava-Díaz C y Sánchez-Pale JR 2023. Antagonistic bacteria againstFusariumspp. isolated from sclerotia ofClaviceps giganteain maize (Zea mays). Mexican Journal of Phytopathology, 41 : 143-164. 10.18781/R.MEX.FIT.2208-2 [ Links ]
Bacon C, Yates I, Hinton D y Filmore M 2001. Biological control ofFusarium moniliformein maize. Environmental Health Perspective, 109: 325-332. 10.2307/3435026 [ Links ]
Bhargavi G, Arya M y Jambhulkar PP 2024. Evaluation of biocontrol efficacy of rhizosphere dwelling bacteria for management ofFusariumwilt andBotrytisgray mold of chickpea. BMC Genomic Data, 25 : 7. 10.1186/s12863-023-01178-7 [ Links ]
Bhattacharyya PN y Jha DK 2012. Plant growth-promoting rhizobacteria (PGPR): emergence in agriculture. World Journal of Microbiology and Biotechnology, 28: 1327-1350. 10.1007/s11274-011-0979-9 [ Links ]
Biedendieck R, Knuuti T y Moore SJ 2021. The “beauty in the beast”-the multiple uses ofPriestia megateriumin biotechnology. Applied Microbiology and Biotechnology, 105: 5719-5737. 10.1007/s00253-021-11424-6 [ Links ]
Canchignia-Martínez HF, Vera-Benites LF, Tapia-Quintana DN, Cedeño-Moreira AV, García-Intriago E et al. 2024. Caracterización bioquímica de rizobacterias endófitas con actividad biocontroladora contraPhytophthora palmivorayLasiodiplodia theobromae. Terra Latinoamericana, 42: 1-10. 10.28940/terra.v42i0.1807 [ Links ]
Castro del Ángel E, Hernández-Castillo F, Gallegos-Morales G y Ochoa-Fuentes Y 2021. Actividad antifúngica de bacterias endófitas para el control deFusarium verticillioidesen maíz. Ecosisistemas y Recursos Agropecuarios, 8 : e2790. 10.19136/era.a8n2.2790 [ Links ]
Edi-Premono M, Moawad MA y Vleck P 1996. Effect of phosphate solubilizingPseudomonas putidaon the growth of maize and its survival in the rhizosphere. Indonesian Journal of Crop Science, 11: 13-23. [ Links ]
Figueroa-López AM, Cordero-Ramírez JD, Martínez-Álvarez JC, López-Meyer M, Lizárraga-Sánchez G et al. 2016. Rhizospheric bacteria of maize with potential for biocontrol ofFusarium verticillioides. SpringerPlus, 5: 330. 10.1186/s40064-016-1780-x [ Links ]
Figueroa-Rivera MG, Rodríguez-Guerra R, Guerrero-Aguilar BZ, González-Chavira MM, Pons-Hernández JL et al. 2010. Caracterización de especies deFusariumasociadas a la pudrición de raíz de maíz en Guanajuato, México. Revista Mexicana de Fitopatología, 28: 124-134. [ Links ]
Gravel V, Antoun H y Tweddell R 2007. Growth stimulation and fruit yield improvement of greenhouse tomato plants by inoculation withPseudomonas putidaorTrichoderma atroviride: Possible role of indole acetic acid (IAA). Soil Biology and Biochemistry, 39 : 1968-1977. 10.1016/j.soilbio.2007.02.015 [ Links ]
Herrera-Sepúlveda A, Argentel-Martínez L, Peñuelas-Rubio O y Azizoglu U 2023. First report ofSinomonas halotoleransfromParkinsonia aculeatarhizosphere. Biologia, 79: 621-627. 10.1007/s11756-023-01581-6 [ Links ]
Jones BV, Sun F y Marchesi JR 2007. Using skimmed milk agar to functionally screen a gut metagenomic library for proteases may lead to false positives. Letters in Applied Microbiology, 45: 418-420. [ Links ]
Khalil MMR, Fierro-Coronado RA, Peñuelas-Rubio O, Villa-Lerma AG, Plascencia-Jatomea R et al. 2021. Rhizospheric bacteria as potential biocontrol agents againstFusariumwilt and crown and root rot diseases in tomato. Saudi Journal of Biological Sciences, 28 : 7460-7471. 10.1016/j.sjbs.2021.08.043 [ Links ]
Leyva-Madrigal K, Larralde-Corona C, Apodaca-Sánchez M, Quiroz-Figueroa F, Mexia-Bolaños P et al. 2014. Fusariumspecies from deFusarium fujikuroispecies complex involved in mixed infections of maize in northern Sinaloa, Mexico. Journal of Phytopatology, 163 : 486-497. 10.1111/jph.12346 [ Links ]
Pangihotan NT 2025. Agricultural relevance of fungal mycelial growth-promoting bacteria: Mutual interaction and application. Microbiological Research, 290: 127978. 10.1016/j.micres.2024.127978 [ Links ]
Peñuelas-Rubio O, Argentel-Martínez L, Cervantes Ortíz F, Vázquez GD, Díaz GIM et al. 2024a. Presencia deFusariumsp. en maíces nativos cultivados bajo distintos regímenes de riego. Pp. 87-95. In: Zuffo AM, Gonzalez AJ, Façanha ML (eds.). Inovações em pesquisas agrárias e ambientais - Volume V. Nova Xavantina-MT: Pantanal, Brasil. [ Links ]
Peñuelas-Rubio O, Argentel-Martínez L, Herrera-Sepúlveda A, Maldonado-Mendoza I, Gonzalez-Aguilera J et al. 2024b. Rhizobacteria associated withParkinsonia aculeataL. under drought and saline conditions in semi-desert of Sonora, Mexico. Biology, 79 : 2591-2603. 10.1007/s11756-024-01731-4 [ Links ]
Pérez-Miranda S, Cabirol N, George-Téllez R, Zamudio-Rivera LS, Fernández FJ 2007. O-CAS, a fast and universal method for siderophore detection. Journal of Microbiological Methods, 70: 127-131. [ Links ]
Pikovskaya R 1948. Mobilization of phosphorus in soil connection with the vital activity of some microbial species. Microbiology, 17: 362-370. [ Links ]
Riddell RW 1950. Permanent stained mycological preparations obtained by slide culture. Mycologia, 42 : 265-270. [ Links ]
Roberts WK y Selitrennikoff CP 1988. Plant and bacterial chitinases differ in antifungal activity. Journal of General Microbiology, 134: 169-176. [ Links ]
Sallam AA, Haroun SA, Aboulnaga EA y Mowafy AM 2024. Enterobacter cloacaeinduces SA-dependent systemic acquired resistance ofZea maysagainstFusarium oxysporum. Journal of Plant Growth Regulation, 43: 2536-2554. 10.1007/s00344-024-11280-4 [ Links ]
Salwoom L, Rahman RNZRA, Salleh AB, Shariff FM, Convey P et al. 2019. Isolation, characterization, and lipase production of a cold-adapted bacterial strainPseudomonassp. LSK25 isolated from Signy Island, Antarctica. Molecules, 24: 1-14. [ Links ]
Sánchez-Ceja MG, Loeza-Lara PD, Carballar-Hernández S, Jiménez-Mejía R y Medina-Estrada RI 2023. Aislamiento de bacterias nativas con potencial en la promoción del crecimiento de maíz criollo mexicano (Zea maysL.). Biotecnia, 26: e2125. 10.18633/biotecnia.v26.2125 [ Links ]
SENASICA 2020. Ficha técnicaFusariumspp.. Dirección General de Sanidad Vegetal, SADER. https://www.gob.mx/cms/uploads/attachment/file/633031/Fusarium_spp ma_z 2020.pdf [ Links ]
Sharma K, Singh N, Vaishnav A, Singh V y Singh S 2023. Fenugreek associated bacteriumPriestia endophyticaSK1 induces defense response againstfusariumwilt by accumulation of secondary metabolites. South African Journal of Botany, 160: 229-234. [ Links ]
Stead D 1984. Evaluation of a fluorimetric assay on the lipases from strains of milk psychrotrophic bacteria. Journal of Dairy Research, 51: 123-130. [ Links ]
Sumerell B, Laurence M, Liew E y Leslie J 2010. Biogeography and phylogeography ofFusarium: a review. Fungal Diver, 44: 3-13. 10.1007/s13225-010-0060-2 [ Links ]
Teather RM y Wood PJ 1982. Use of Congo red-polysaccharide interactions in enumeration and characterization of cellulolytic bacteria from the bovine rumen. Applied and Environmental Microbiology, 43: 777-780. [ Links ]
Velarde F, Valdez N, Zamora F, López R, Melgoza C et al. 2018. Identificación molecular deFusariumspp. aislados de maíz en Sinaloa, México. Revista Mexicana de Ciencias Agrícolas, 9 : 1675-1689. 10.29312/remexca.v9i8.756 [ Links ]
Received: January 15, 2026; Accepted: April 25, 2026
 Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
