










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de fitopatología
versión On-line ISSN 2007-8080versión impresa ISSN 0185-3309
Rev. mex. fitopatol vol.44 no.1 Texcoco ene. 2026 Epub 23-Jun-2026
https://doi.org/10.18781/r.mex.fit.2407-3
Phytopathological notes
Morphological and molecular characterization of Trichoderma strains isolated from the BioHumisol ® biofertilizer
11Facultad de Ciencias Químico-Biológicas, Universidad Autónoma de Sinaloa, Calzada de las Américas, Blvd. Universitarios, Cd. Universitaria, C.P. 80040, Culiacán, Sinaloa, México;
2Facultad de Biología, Universidad Autónoma de Sinaloa, Calzada de las Américas, Blvd. Universitarios, Cd. Universitaria, C.P. 80040, Culiacán, Sinaloa, México;
3Centro de Investigación en Alimentación y Desarrollo, A. C. Coordinación Culiacán, Carretera Culiacán-El dorado Km 5.5, Campo El Diez, C.P. 80110, Culiacán, Sinaloa, México.
Background/Objetive.
Biofertilizers exist that contain microorganisms beneficial to plants and the environment. However, some commercial products provide little information about the species they use. Therefore, this study aimed to identify, culturally, morphologically, morphometrically, and molecularly, the Trichoderma species present in the BioHumisol® fertilizer used in agriculture in Culiacán and Navolato, Sinaloa, México.
Experimental development.
Fungal colonies were isolated and purified from the BioHumisol® liquid biofertilizer. For cultural, morphological, and morphometric identification, three Petri dishes were used, each containing one fungal isolate. The following characteristics were recorded: color, mycelium shape and arrangement in the culture medium, days of sporulation, and the size and shape of conidia, phialides, and chlamydospores. The mycelial growth rate was also recorded. At the molecular level, the ITS region and the TEF-1 α gene were amplified. The sequences were registered in GenBank with the accession numbers: PP956784, PP956785 for ITS; PP968400, PP968399 for TEF-1 α. Phylogenetic analysis of the concatenated sequences was performed using the maximum likelihood and Bayesian inference method with a bootstrap of 1000 replicates.
Results.
The fungal colonies presented hyaline, branched, non-whorled conidiophores; phialides either single or in groups with terminal branches worth average measurements of 4.1-6.2 × 9.7-15.3 µm; unicellular ovoid conidia with average dimensions of 3.6-5.09 × 3.27-6.41 µm and subglobose intercalary and terminal chlamydospores measuring 21.41 × 21.72 µm consistent with Trichoderma. Currently accurate identification requires cultural, morphological and morphometric characterization, complemented by molecular characterization. BLAST analysis of partial ITS sequences showed 99% homology, and the TEF -1 α gene showed 100% homology with T. asperellum. Phylogenetic analyses confirm genetic proximity to species of the Viride clade.
Conclusion.
Based on the cultural, morphological, morphometric and molecular characterization, the species found in the liquid biofertilizer BioHumisol® is Trichoderma asperellum.
Keywords: Trichoderma asperellum; ITS; TEF -1 α; Biocontrol
Antecedentes/Objetivo.
Existen biofertilizantes, que contienen microorganismos benéficos para la planta y el ambiente. Sin embargo, algunos productos comerciales proporcionan poca información de las especies que emplean en sus productos; por lo que este trabajo tuvo el objetivo de identificar cultural, morfológica, morfométrica y molecularmente las especies de Trichoderma presentes en el biofertilizante BioHumisol® de uso agrícola en Culiacán y Navolato, Sinaloa, México.
Desarrollo experimental.
A partir del biofertilizante líquido BioHumisol® se aislaron y purificaron colonias fúngicas. Para la identificación cultural, morfológica y morfométrica se emplearon tres cajas Petri con crecimiento fúngico por aislado, se registraron las características de color, forma del micelio y disposición en el medio de cultivo, días de esporulación, tamaño y forma de conidios, fiálides y clamidosporas. Además, se registró la tasa de crecimiento micelial. A nivel molecular se amplificó la región ITS y el gen TEF- 1 α, las secuencias se registraron en GenBank con los números de acceso: PP956784, PP956785 para ITS; PP968400, PP968399 para TEF-1 α, y se realizó el análisis filogenético de las secuencias concatenadas mediante el método de máxima verosimilitud e inferencia Bayesiana con un bootstrap de 1000 réplicas.
Resultados.
Las colonias fúngicas presentaron conidióforos hialinos, ramificados, no verticilados; fiálides simples o en grupos con ramificaciones terminales con medidas promedio de 4.1-6.2 × 9.7-15.3 µm; conidios unicelulares, ovoides con medidas promedio de 3.6-5.09 × 3.27-6.41 µm y clamidosporas subglobosas intercalares y terminales cuyo tamaño fue de 21.41 × 21.72 µm, lo que coincide con Trichoderma. Actualmente, para una correcta identificación se requiere la caracterización cultural, morfológica, morfométrica completada con la caracterización molecular. El análisis BLAST de las secuencias parciales de ITS mostraron una homología del 99 % y del gen TEF -1 α una homología del 100 % con T. asperellum, el análisis filogenético confirma cercanía genética con especies del clado Viride.
Conclusión.
De acuerdo con la caracterización cultural, morfológica, morfométrica y molecular la especie que se encuentra en el biofertilizante líquido BioHumisol® es Trichoderma asperellum.
Palabras clave: Trichoderma asperellum; ITS; TEF -1 α; Biocontrol
Introduction
Agriculture in Mexico relies on fertilizers that supply nutrients to crops to increase both yield and product quality. This enables producers to achieve higher productivity and, consequently, greater economic income (SAGARPA, 2025). In Sinaloa, chemical fertilizers are used in approximately 98% of agricultural production. However, beyond the economic cost of their acquisition, the excessive use of chemicals has ecological and environmental repercussions (Debnath et al., 2020).
Biofertilizers contain microorganisms capable of synthesizing chemical compounds that promote plant growth, help fix atmospheric nitrogen, and solubilize or mobilize soil nutrients; an example is the fungus Trichoderma (Shahwar et al., 2023). The benefits of using Trichoderma spp. include promoting plant growth and development through growth factors, lowering soil pH, solubilizing components of plant metabolism, and reducing soil pollution through bioremediation (Rodríguez-García and Vargas-Rojas, 2022). Various mechanisms have been proposed to explain how Trichoderma species promote plant growth. Among these are the biosynthesis of secondary metabolites, which promotes the production of enzymes such as xylanase, cellulase, and glucanase; the solubilization of phosphorus; the synthesis of indole-3-acetic acid; and the stimulation of root and stem growth, as well as the enhancement of carbohydrate metabolism, photosynthesis, and plant defense mechanisms (Contreras-Cornejo et al., 2024).
In addition, Trichoderma functions as a biological control agent by combating phytopathogenic bacteria, fungi, and nematodes through various mechanisms, including competition for substrate and space, antibiosis, antagonism, mycoparasitism, recognition, chemotrophic growth, lytic activity, and the promotion of induced systemic resistance in plants (Yao et al., 2023; Sánchez-Rivera et al., 2025). In Mexico, T. harzianum, T. viride,
T. koningii, T. asperellum, T. longibrachiatum, T. virens, T. lignorum, and T. aureoviridae are reproduced and commercialized as biological control agents (Allende-Molar et al., 2022; SENASICA, 2025). Certified Trichoderma species used for biotechnological purposes in biofertilizer formulations include T. harzianum, T. viride, T. asperellum, T. virens, T. atroviride, T. gamsii, T. hamatum, and T. polysporum. Most biofertilizers are prepared using T. viride, T. virens, and predominantly T. harzianum (Hernández-Melchor et al., 2019; Yao et al., 2023).
The company Humisol Orgánico S.A. de C.V. produces and commercializes the liquid biofertilizer BioHumisol®, which, according to its label, contains anaerobic and nitrifying bacteria; however, the Trichoderma species present in the commercial product are unknown. The BioHumisol® biofertilizer is used in crops such as berries, asparagus, and vegetables. Since no certified strain is available and the Trichoderma species present in this biofertilizer are unknown, cultural, morphometric, and molecular characterization was necessary (BioHumisol, 2025). Therefore, the aim of this study was to identify, at the cultural, morphological, morphometric, and molecular levels, the Trichoderma species present in the commercial liquid biofertilizer BioHumisol®, used in agriculture in Culiacán and Navolato, Sinaloa, Mexico.
Experimental development
Sampling, isolation, purification, and preservation. Sampling was conducted at the company Humisol Orgánico S.A. de C.V., located in Cofradía de San Pedro, Navolato, Sinaloa, Mexico, where the liquid biofertilizer BioHumisol® (registration RSCO-381/X/21) is produced and commercialized in the municipalities of Culiacán and Navolato, Sinaloa. For fungal isolation, a stock solution of the final product was prepared by placing 50 mL of the liquid biofertilizer into a glass flask containing 450 mL of sterile distilled water and shaking it for 30 s (stock solution). Subsequently, six serial dilutions (10⁻¹ to 10⁻⁶) were prepared by transferring 1 mL of the previous dilution into a glass tube containing 9 mL of sterile distilled water, followed by 30 s of vortex mixing (Cis-Lab, Mexico) to homogenize the solution (Suárez-Palacios et al., 2023). From this suspension, serial dilutions were made up to 1 × 10⁻⁶. Additionally, 100 µL of the conidial suspension were spread on Petri dishes containing potato dextrose agar (PDA, Bioxon®) supplemented with lactic acid (PDA-LA). The conidial suspension was homogenized using sterile 0.8 mm glass beads. Petri dishes were inoculated in triplicate for each dilution and incubated at (±) 28 °C for four days in an incubator (Ecoshel, USA) (Savín-Molina et al., 2021). Purification was performed using the hyphal-tip technique (Cuervo-Parra et al., 2024). Once pure hyphal-tip cultures were obtained, 5 mm mycelial disks were taken with a cork borer and preserved in 1.5 mL tubes containing sterile distilled water (Castellani method), then stored at 4 °C until further use (Fernández et al., 2013).
Cultural, morphological, and morphometric identification. For cultural, morphological, and morphometric identification, three Petri dishes with fungal growth per isolate were used. The following characteristics were recorded: color, mycelial form and arrangement on the culture medium, sporulation time, and the size and shape of conidia, phialides, and chlamydospores (Ynfante-Martínez et al., 2023). The structures were observed using a digital camera (Dino-Lite) attached to a compound microscope, CxL series (Labomed®) (Sánchez-Hernández et al., 2018). Measurements of conidia, phialides, and chlamydospores (n = 50) were taken using the Dino Capture 2.0 software (Zainudin et al., 2023). Cultural, morphological, and morphometric characteristics were compared with the taxonomic features reported by Barnett and Hunter (1998) and Samuels and Hebbar (2015). In addition, the mycelial growth rate of the strains was determined by continuous measurement of Petri dishes containing PDA-LA culture medium incubated at (±) 28 °C under 12 h light / 12 h dark conditions for four days. The mycelial growth rate was calculated by measuring the radial growth diameter in perpendicular directions with a vernier caliper every 24 h for four days. Growth rate was expressed in mm/day (Samaniego et al., 2018).
Molecular identification. Three Petri dishes with contamination-free fungal growth per isolate were used. Mycelium was scraped with a sterile spatula from Petri dishes containing seven-day-old fungal cultures grown on PDA-LA medium. Subsequently, 150 mg of mycelium were transferred into sterile 1.5 mL tubes. Fungal DNA extraction was then performed following the manufacturer’s instructions for the commercial ZR Fungal/Bacterial DNA Miniprep® Kit (Zymo Research, USA) (Manfredini et al., 2025). To verify DNA concentration and purity, samples were analyzed using a spectrophotometer (QIAxpert, QUIAGEN®), and their integrity was confirmed by electrophoresis on a 1% agarose gel. DNA samples were stored in an ultrafreezer at −80 °C until further use (Matas- Baca et al., 2022). The Internal Transcribed Spacer (ITS) region was amplified using the primers ITS4 (5′TCCTCCGCTTATTGATATGC3′) and ITS5 (5′GGAAGTAAAAGTCGTAACAAGG3′), under the following amplification conditions:
initial denaturation at 94 °C for 5 min, followed by 29 cycles of denaturation at 94 °C for 30 s, annealing at 60 °C for 30 s, and extension at 72 °C for 45 s, with a final extension at 72 °C for 5 min (Ríos et al., 2016).
In addition, the Elongation Factor 1 alpha (TEF-1α) gene of isolates ATV and ATB was amplified using the primers EF1-728 (5′CATCGAGAAGTTCGAGAAGG3′) and TEF1R (5′GCCATCCTTGGGAGATACCAGC3′), under the following amplification conditions: initial denaturation at 94 °C for 2 min, followed by 29 cycles of denaturation at 94 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 1 min, with a final extension at 72 °C for 10 min (Lee et al., 2020). Amplification was performed in a Biorad C1000 thermocycler with a final volume of 25 µL (Samuels et al., 2002; García-Nuñez et al., 2017). The integrity of the PCR products was verified by electrophoresis on a 1% agarose gel stained with ethidium bromide, and the gel was visualized using a UV photodocumentation system (Axygen™, USA) (Lee et al., 2020).
The PCR products obtained were purified and sequenced using the Sanger method and sent to the Advanced Genomics Unit of LANGEBIO, Cinvestav, Irapuato. The resulting sequences were analyzed with the BioEdit software (García-Nuñez et al., 2017) and submitted to GenBank under the following accession numbers: PP956784, PP956785, PP968400, and PP968399. Subsequently, they were compared with other sequences available in the GenBank database of the National Center for Biotechnology Information (www.ncbi.nlm.nih.gov/) using the BLAST (Basic Local Alignment Search Tool) algorithm (NCBI, 2024).
Phylogenetic analysis. Partial nucleotide sequences of the ITS region and the TEF-1α gene were aligned with reference sequences obtained from GenBank, and phylogenetic relationships were inferred based on multiple sequence alignment using the MAFFT program (Vences et al., 2022). The sequences were concatenated with the Mesquite software, version 3.8. The phylogenetic tree was constructed using the maximum likelihood and Bayesian inference methods. To determine confidence values for the clades within the resulting tree, a bootstrap analysis with 1000 replicates was performed using the TPM2+F+G4 substitution model in IQTREE, and the tree was visualized in FigTree version 1.4.4 (Zhao et al., 2024). The sequences used in the phylogenetic analysis are shown in Table 1. A sequence of Fusarium oxysporum (GenBank accession numbers: OR734798 and ON316841) was used as the outgroup.
Table 1 Species and GenBank accession numbers of the sequences used in the phylogenetic analysis
| Specie | Strain number | GenBank number | GenBank number |
|---|---|---|---|
| (ITS) | (TEF 1-α) | ||
| T. atroviride | IMI206040 | OR975627.1 | MK644115.1 |
| T. composticola | CBS133497 | MT187974.1 | DQ841716.1 |
| T. asperellum z | ATV | PP956784 | PP968400 |
| T. asperellum z | ATB | PP956785 | PP968399 |
| T. asperellum | CBS43397 | OR770586.1 | OL825045.1 |
| T. asperellum | CBS43397 | MN727373.1 | MW457023.1 |
| T. flagellatum | CBS130626 | MH865822.1 | FJ763158.1 |
| T. orientale | CBS130428 | PP464121.1 | OL757487.1 |
| T. bissettii | CBS137447 | MW295457.1 | HG931271.1 |
| T. virens | FT333 | ON357718.1 | OQ702629.1 |
| T. crassum | CBS33693 | PP860359.1 | KJ871144.1 |
| T. alni | CBS120633 | MK459321.1 | KJ665356.1 |
| T. amazonicum | CBS126898 | MH864268.1 | HM142378.1 |
z Sequences from this study.
Cultural, morphological, and morphometric identification. This study provides the cultural, morphological, morphometric, and molecular identification of two isolates with characteristics like Trichoderma, designated ATV and ATB, obtained from the agricultural liquid biofertilizer BioHumisol®. After four days, the colonies consisted of white, powdery-textured mycelium that turned green with age; abundant sporulation and radial growth were observed. The average daily radial growth rate of the two isolates over four days was 26, 57, 84, and 90 mm. The fungal colonies exhibited morphological characteristics such as hyaline, branched, non-verticillate conidiophores; phialides simple or in terminally branched groups; turquoise, unicellular, ovoid conidia; and subglobose intercalary and terminal chlamydospores (Figure 1). Conidia (n = 50) measured 3.6-5.09 ×
3.27-6.41 µm, phialides (n = 50) measured 4.1-6.2 × 9.7-15.3 µm, and chlamydospores measured 21.41 × 21.72 µm. Based on cultural, morphological, and morphometric characteristics, both isolates were tentatively identified as Trichoderma according to the taxonomic keys of Barnett and Hunter (1998) and Samuels and Hebbar (2015).
The size of the conidia coincides with that reported for T. asperellum (Calle-Cheje et al., 2023); however, in this study, the conidia were larger than those described in other morphological characterizations of T. asperellum (Sebumpan et al., 2022; Andrade-Hoyos et al., 2023). Trichoderma is widely used as a biofertilizer and biopesticide in Mexican agriculture for various economically important crops due to its multiple mechanisms of action. T. asperellum is employed in bioformulations in Mexico and exhibits morpho- cultural variability (Andrade-Hoyos et al., 2023). Furthermore, Foyate (2023) reported diversity in morphological traits such as pigmentation, and the size and shape of conidia and phialides. This morphological variability may be related to the type of formulation used in commercial products. On the other hand, morpho-cultural variability makes precise identification of Trichoderma sp. difficult, thus requiring the use of different taxonomic keys and molecular identification (Ynfante-Martínez et al., 2023).
Molecular identification. The sequences of isolates ATV and ATB were deposited in GenBank (ITS: PP956784, PP956785; TEF 1-α: PP968400, PP968399). The BLAST
analysis of the partial ITS sequences (549 bp and 588 bp) showed 99% similarity with T. asperellum isolates (KY750369, KY750373), while the partial TEF 1-α sequences (631 bp and 624 bp) showed 100% similarity with T. asperellum (JQ040494, KP747448). The phylogenetic tree obtained from the concatenated sequences of this study and other Trichoderma species from different clades indicate that the isolates obtained from the BioHumisol® biofertilizer belong to the Viride clade (Figure 2).
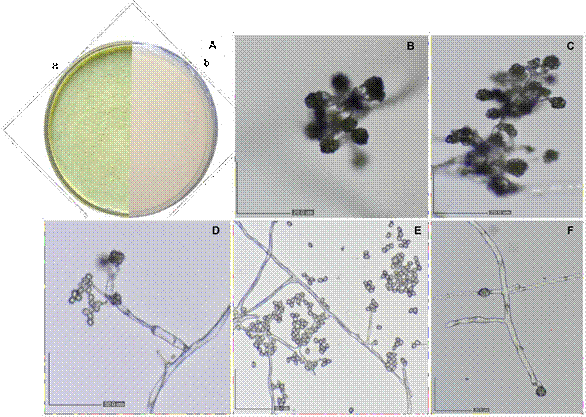
Figure 1 Morphology of Trichoderma asperellum isolates. A) Petri dish after seven days of growth on PDA- LA: a) front view, b) reverse view; B-D) conidiophores with phialides and clustered conidia; E) hyphae and conidia; F) chlamydospores.
Currently, accurate identification requires cultural, morphological, and morphometric characterization complemented by molecular characterization. Several studies have addressed the molecular characterization of T. asperellum using the ITS gene region (Ma et al., 2020; Sehim et al., 2023). In the present study, the percentage of homology with T. asperellum using the ITS region was 99%. Reports of molecular studies employing the ITS region have mentioned a 98% similarity (GenBank accession number: MN950427) for a native Trichoderma asperellum species isolated from soil in Guasave, Sinaloa, Mexico (Matas-Baca et al., 2022). In Mexico, the molecular identification of T. asperellum is primarily based on the ITS region and the TEF-1α gene (Allende-Molar et al., 2022). The universal marker for molecular identification in fungi is the ITS region (Maldonado-Bonilla et al., 2024); however, it does not allow differentiation among closely related species and provides low resolution within the Trichoderma genus (Samuels et al., 2002). Consequently, this genus is considered notoriously difficult to identify, and the use of more than one gene is recommended to increase specificity and provide greater robustness to molecular identification (Samuels and Hebbar, 2015).
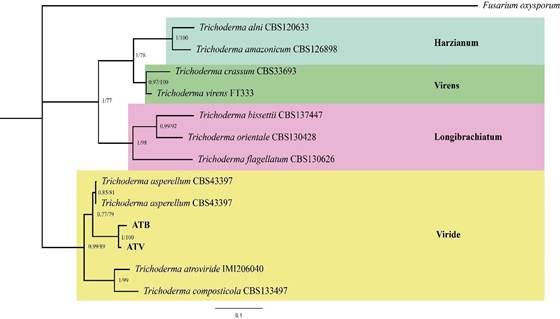
Figure 2 Phylogenetic tree based on concatenated partial ITS and TEF 1-α sequences of Trichoderma species. The tree was constructed using the maximum likelihood and Bayesian inference methods with 1000 replicates. Bayesian posterior probabilities (PP > 0.5) and bootstrap values (BS > 50) are shown at the nodes. The sequences from this study are shown in bold. Fusarium oxysporum (GenBank accession numbers: OR734798 and ON316841) was used as the outgroup.
A widely used gene for the molecular identification of Trichoderma is TEF-1α, as it allows for the assessment of the accuracy and robustness of genetic markers used in fungi, aiding in the identification of previously undescribed species. The TEF-1α gene has been reported to possess high phylogenetic efficacy due to its stronger association with the identification of unknown species and its alignment among Trichoderma species (Hewedy et al., 2020). According to García-Nuñez et al. (2017), the identity percentage for the molecular identification of T. asperellum using the TEF-1α region was 99%, and they suggest that phylogenetic analysis is more accurate when both the ITS region and the TEF- 1α gene are used. This analysis should be complemented with morphological characterization for reliable identification. Molecular identification of Trichoderma requires the use of one or more markers-ITS, TEF1, ACT, CAL, ACL1, CHI18-5, and RPB2-along with phylogenetic analysis. Based on this approach, the genus is grouped into 24 clades (Samuels and Hebbar, 2015), with the Harzianum, Viride, Virens, and Longibrachiatum clades being particularly notable (Rodríguez et al., 2021).
The Viride clade encompasses the largest number of species (54), which are morphologically similar, including T. asperellum, T. asperelloides, and T. viride (Samuels and Hebbar, 2015; Allende-Molar et al., 2022). The phylogenetic tree based on ITS and TEF-1α markers confirms that T. asperellum isolated from the Biohumisol® biofertilizer produced by Humisol Orgánico S.A. de C.V. belongs to the Viride clade-a species previously reported within this group (Samuels and Hebbar, 2015). The isolates obtained in this study are genetically related to other T. asperellum species when compared with other species grouped within the same clade (T. atroviride and T. composticola). According to Sánchez-Miranda et al. (2021), phylogenetic analysis allows the association of various native Trichoderma species, including T. asperellum, isolated from different crops and localities. The genetic proximity among Trichoderma sequences isolated from distinct geographical regions may be due to the genetic regions used, which are highly conserved among species, resulting in high homology even in geographically distant isolates. Moreover, this genus is cosmopolitan and exhibits high ecological adaptability, which facilitates its global dispersal.
Conclusions
Cultural, morphological, and morphometric characterization (with average measurements of phialides: 4.1-6.2 × 9.7-15.3 µm; conidia: 3.6-5.09 × 3.27-6.41 µm; and chlamydospores: 21.41 × 21.72 µm), together with molecular characterization of Trichoderma strains isolated from the agricultural liquid biofertilizer Biohumisol® in Culiacán and Navolato, Sinaloa, Mexico, enabled the identification of Trichoderma asperellum. The biotechnological importance of this species supports the effectiveness of the Biohumisol® liquid biofertilizer in regional agriculture. Moreover, the integration of morphological and molecular methods ensures precise identification, which is essential for developing sustainable agricultural management strategies based on native microorganisms with high adaptive capacity.
Acknowledgments
The authors thank the Program for the Promotion and Support of Research Projects (PROFAPI 2022) for financial support of project code PRO_A7_078, which contributed to strengthening collaboration between the scientific and private sectors. The authors also acknowledge the technical assistance of undergraduate thesis students Arlen Yadira Partida Durán and Diego López Silvas, as well as Biol. Víctor Manuel Angulo Medina and Biol. Nahomy Monserrat Escalera Mares.
REFERENCES
Allende-Molar R, Báez-Parra K, Salazar-Villa E y Rojo-Báez I. 2022. Biodiversidad de Trichoderma spp. en México y su uso potencial en la agricultura. Agroecosistemas tropicales y subtropicales 25 (https://www.google.com/search?q=%5Bissueno%5D3%5B/issueno%5D). http://dx.doi.org/10.56369/tsaes.4297 [ Links ]
Andrade-hoyos P, Rivera-Jiménez MN, Landero-Valenzuela N, Silva-Rojas HV, Martínez-Salgad M, et al. 2023. Beneficios ecológicos y biológicos del hongo cosmopolita Trichoderma spp. en la agricultura: una perspectiva en el campo mexicano. Revista Argentina de Microbiología 55 (https://www.google.com/search?q=%5Bissueno%5D4%5B/issueno%5D): 366-377. https://doi.org/10.1016/j.ram.2023.06.005 [ Links ]
Barnett HL and Hunter BB. 1998. Ilustrated genera of imperfect fungi. EE. UU. Burgess Publishing Company. 241pp. http://dx.doi.org/10.2307/3757954 [ Links ]
Biohumisol 2025. Humisol orgánico, S. A de C. V. https://www.humisolorganico.com.mx/ (Consulta, junio 2025). [ Links ]
Calle-Cheje YH, Aguilar-Anccota R, Rafael-Rutte R y Morales-Pizarro A. 2023. Formulación y conservación del hongo antagonista Trichoderma asperellum como polvo mojable y emulsionable. Idesia (Arica) 41 :43-53. https://doi.org/10.4067/S0718-34292023000400043 [ Links ]
Contreras-Cornejo HA, Schmoll M, Esquivel-Ayala BA, González-Esquivel CE, et al. 2024. Mechanisms for plant growth promotion activated by Trichoderma in natural and managed terrestrial ecosystems. Microbiological Research 281: 127621. https://doi.org/10.1016/j.micres.2024.127621 [ Links ]
Cuervo-Parra JA, Romero-Cortes T, Román AR, Valle HJ y Aparicio-Burgos HE. 2024. Identificación morfológica de hongos aislados de plantas de Garcinia mangostana. Revista Mexicana de Ciencias Agrícolas 15 : e3575. https://doi.org/10.29312/remexca.v15i5.3575 [ Links ]
Debnath S, Chakraborty G, Dutta SS, Chaudhruri SR, Das P and Saha AK. 2020. Potential of Trichoderma species as biofertilizer and biological control on Oryza sativa L. cultivation. Biotecnología vegetal 20 :1-16. http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S2074-86472020000100001 [ Links ]
Fernández-Andreu CM, Díaz-Suárez LA, Illnait-Zaragozí MT, Aragonés-López C, Martínez-Machín G, et al. 2013. Conservación de cultivos de hongos de importancia médica en agua destilada. Revista Cubana de Medicina Tropical 65 : 361-369. http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S037507602013000300009 [ Links ]
Foyate MS. 2023. Morphological Variations Observed in Trichoderma asperellum Isolated from Commercial Products. International Multidisciplinary Research Journal 13: 6-9. https://doi.org/10.25081/imrj.2023.v13.8230 [ Links ]
García-Núñez HG, Martínez-Campos AR, Hermosa- Prieto MR, Monte-Vázquez E, Aguilar-Ortigoza CJ, et al. 2017. Morphological and molecular characterization of native isolates of Trichoderma and its potential biocontrol against Phytophthora infestans. Revista Mexicana de Fitopatología 35: 58-79. https://doi.org/10.18781/R.MEX.FIT.1605-4 [ Links ]
Hernández-Melchor DJ, Ferrera-Cerrato R y Alarcón A. 2019. Trichoderma: Importancia agrícola, biotecnológica y sistemas de fermentación para producir biomasa y enzimas de interés industrial. Chilean Journal of Agricultural & Animal Science 35 :98-112. https://dx.doi.org/10.4067/S0719-38902019005000205 [ Links ]
Hewedy OA, Abdel-Lateif KS, Seleiman MF, Shami A, Albarakaty FM, et al. 2020. Phylogenetic diversity of Trichoderma strains and their antagonistic potential against soil-borne pathogens under stress conditions. Biology 9 : 189. https://dx.doi.org/10.3390/biology9080189 [ Links ]
Lee SH, Jung HJ, Hong SB, Choi JI and Ryu JS. 2020. Molecular markers for detecting a wide range of Trichoderma spp. That might potentially cause green mold in Pleurotus eryngii. Microbiology 48 (https://www.google.com/search?q=%5Bissueno%5D4%5B/issueno%5D): 313-320. https://doi.org/10.1080/12298093 [ Links ]
Ma J, Tsegaye E, Li M, Wu B and Jiang X. 2020. Biodiversity of Trichoderma from grassland and forest ecosystems in Northern Xinjiang, China. 3 Biotech, 10 : 362. https://doi.org/10.1007/s13205-020-02301-6 [ Links ]
Maldonado-Bonilla LD, Sánchez-Espinosa AC y Villarruel-Ordaz JL. 2024. Identificación de hongos mediante códigos de barras de ADN. Ciencia ergo sum 31, e249. https://doi.org/10.30878/ces.v31n0a33 [ Links ]
Manfredini A, Pugliese M, Valfre P and Canfora L. 2025. Advancing strain-specific TaqMan assay for Trichoderma asperellum detection in commercial agricultural settings. Biological Control 202:105723. https://doi.org/10.1016/j.biocontrol.2025.105723 [ Links ]
Matas-Baca MÁ, Urías-García C, Pérez-Álvarez S, Flores-Córdova MA, Escobedo-Bonilla CM, et al. 2022. Morphological and molecular characterization of a new autochthonous Trichoderma sp. isolate and its biocontrol efficacy against Alternaria sp. Saudi Journal of Biological Sciences 29 :2620-2625. http://dx.doi.org/10.1016/j.sjbs.2021.12.052 [ Links ]
National Center for Biotechnology Information (NCBI). 2024. Bethesda (MD): National Library of Medicine (US). https://www.ncbi.nlm.nih.gov/. (Consulta, junio 2024) [ Links ]
Ríos VC, Caro CJM, Berlanga RDI, Ruiz CMF, Ornelas PJJ, et al. 2016. Identificación y actividad antagónica in vitro de aislados de Bacillus spp. y Trichoderma spp. contra hongos fitopatógenos comunes. Revista Mexicana de Fitopatología 34 : 85-99. http://dx.doi.org/10.18781/R.MEX.FIT.1507-1 [ Links ]
Rodríguez-García D y Vargas-Rojas J. 2022. Efecto de la inoculación con Trichoderma sobre el crecimiento vegetativo del tomate (Solanum lycopersicum). Agronomía Costarricense 46 : 47-60. http://dx.doi.org/10.15517/rac.v46i2.52045 [ Links ]
Rodríguez MC, Evans HC, Abreu LM, Macedo DM, Ndacnou MK, et al. 2021. New species and records of Trichoderma isolated as mycoparasites and endophytes from cultivated and wild coffee in Africa. Scientific reports 11:5671. https://doi.org/10.1038/s41598-021-84111-1 [ Links ]
SAGARPA. 2025. Servicio de Información Agroalimentaria y Pesquera. https://www.siap.gob/idex. (Consulta, junio, 2025). [ Links ]
Shahwar D, Mushtaq Z, Mushtaq H, Alqarawi AA, Park Y, et al. 2023. Role of microbial inoculants as biofertilizers for improving crop productivity: A review. Heliyon 9 : e16134. https://doi.org/10.1016/j.heliyon.2023.e16134 [ Links ]
Samaniego-Fernández LM, Harouna M, Corbea O, Rondón-Castillo AJ y Placeres-Espinosa I. 2018. Aislamiento, identificación y evaluación de cepas autóctonas de Trichoderma spp. antagonistas de patógenos del suelo. Revista de Protección Vegetal 33 . http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S1010-27522018000300003 [ Links ]
Samuels GJ, Dodd SL, Gams W, Castlebury LA and Petrini O. 2002. Trichoderma species associated with the green mold epidemic of commercially grown Agaricus bisporus. Mycologia 94 :146-70. https://doi.org/10.1080/15572536.2003.11833257 [ Links ]
Samuels GJ and Hebbar PK. 2015. Trichoderma: Identification and agricultural applications. St. Paul, Minnesota. APS Press. https://my.apsnet.org/APSStore/Product-Detail.aspx?iProductCode=44846 [ Links ]
Sánchez-Hernández L, Arias-Mota RM, Rosique-Gil JE y Pacheco-Figueroa CJ. 2018. Diversidad del género Trichoderma (Hypocraceae) en un Área Natural Protegida en Tabasco, México. Acta Botánica Mexicana (123): 167-182. https://doi.org/10.21829/abm123.2018.1269 [ Links ]
Sánchez-Miranda MD, Moreno-Mayorga LF y Páramo-Aguilera LA. 2021. Identificación morfológica y molecular de especies autóctonas Trichoderma spp, aisladas de suelos de importancia agrícola. Revista Científica de Ciencia y Tecnología El Higo 11 : 26-42. https://doi.org/10.5377/elhigo.v11i1.11715 [ Links ]
Sánchez-Rivera G, Flores-Moctezuma HE, Castrejón-Ayala F, Gutiérrez-Rojas M y Ruiz-Juárez D. 2025. Análisis de estudios de Trichoderma spp. como agente de control de fitopatógenos del suelo. Tropical and Subtropical Agroecosystems 28-061. http://doi.org/10.56369/tsaes.5914 [ Links ]
Savín-Molina J, Hernández-Montiel LG, Ceiro-Catasú W, Ávila-Quezada GD, Palacios-Espinosa A, et al. 2021. Caracterización morfológica y potencial de biocontrol de especies de Trichoderma aisladas de suelos del semiárido. Revista Mexicana de Fitopatología 39 : 435-451. https://doi.org/10.18781/r.mex.fit.2106-7 [ Links ]
Sebumpan R, Guiritan KR, Suan M, et al. 2022. Morphological and molecular identification of Trichoderma asperellum isolated from a dragon fruit farm in the southern Philippines and its pathogenicity against the larvae of the super worm, Zophobas morio (Fabricius, 1776) (Coleoptera: Tenebrionidae). Egyptian Journal of Biological Pest Control 32, 47. https://doi.org/10.1186/s41938-022-00548-0 [ Links ]
Sehim AE, Hewedy OA, Altammar KA, Alhumaidi MS and Abd ERY. 2023. Trichoderma asperellum empowers tomato plants and suppresses Fusarium oxysporum through priming responses. Frontiers of Microbiology 14: 1140378. https://doi.org/10.3389/fmicb.2023.1140378 [ Links ]
Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria (SENASICA) 2025. Catálogo de laboratorios reproductores y comercializadores de agentes de control biológico en México. https://www.gob.mx/senasica/documentos/directorio-de-laboratorios-reproductores-y-comercializadores-de-agentes-de-control-biologico (Consulta, junio 2025). [ Links ]
Suárez-Palacios C, Remache-Sánchez N, Pico-Rosado J, Paredes-Puga E, Jiménez-Cumbicus J, et al. 2023. Aislamiento y evaluación de cepas nativas de Trichoderma spp., como promotor de desarrollo radicular. Revista Ciencia UNEMI 16 (42): 45-54. https://doi.org/10.29076/issn.2528-7737vol16iss42.2023pp45-54p [ Links ]
Vences M, Patmanidis S, Kharchev V and Renner SS. 2022. Concatenator, a user-friendly program to concatenate DNA sequences, implementing graphical user interfaces for MAFFT and FastTree. Bioinformatics advances 2 : vbac050. https://doi.org/10.1093/bioadv/vbac050 [ Links ]
Yao X, Guo H, Zhang K, Zhao M, Ruan J and Chen J. 2023. Trichoderma and its role in biological control of plant fungal and nematode disease. Frontiers Microbiology 14:1160551. https://doi.org/10.3389/fmicb.2023.1160551 [ Links ]
Ynfante-Martínez D, Martínez-Coca B, Peteira-Delgado B, Reyes-Duque Y, Gil K, et al. 2023. Caracterización morfo-cultural y variabilidad genética y molecular de aislamientos de Trichoderma. Revista de Ciencias Biológicas y de la Salud XXV (https://www.google.com/search?q=%5Bissueno%5D1%5B/issueno%5D):194-203. https://doi.org/10.18633/biotecnia.v25i2.1890 [ Links ]
Zainudin NAIM, Azhar N, Rosli MNH and Nordin NAM. 2023. Identification and characterization of fungi associated with leaf spot disease of rubber trees (Hevea brasiliensis) in Pahang, Malaysia. Journal Proteksi Tanaman 7 : 89-102. https://doi.org/10.25077/jpt.7.2.89-102.2023 [ Links ]
Zhao R, Chen KY, Mao LJ and Zhang CL. 2024. Eleven new species of Trichoderma (Hypocreaceae, Hypocreales) from China. Micology 16, 180-209. https://doi.org/10.1080/21501203.2024.2330400 [ Links ]
Received: July 28, 2024; Accepted: December 10, 2025
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
