











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkJustificación
Especies fitopatógenas del género Alternaria revisten importancia a nivel económica, ambiental, agronómica y fitopatológica en el mundo; son causantes de pérdidas económicas ya que causan enfermedades denominadas mancha y tizón foliar en alrededor de 4 000 especies de plantas de cualquier edad. El síntoma principal es una mancha de color oscuro con anillos concéntricos en las hojas de distintos hospedantes como hortalizas, frutales, cereales, crucíferas y plantas ornamentales (Ogada et al., 2021).
Por otra parte, algunas especies de Alternaria producen metabolitos secundarios como fitotoxinas asociadas a la patogenicidad y producen micotoxinas, como el ácido tenuazónico (TeA), alternariol (ALT), patulina y alternariol monometil éter (AME), que pueden afectar el 50 % de los productos agrícolas y comprometen la seguridad alimentaria. Además, son difíciles de remover durante el procesamiento de alimentos, por lo que una baja concentración de micotoxinas presentan actividad cancerígena, mutagénica y citotóxica. Aunado a ello, se consideran alérgenos y se asocian a infecciones en el tracto respiratorio causando asma en humanos (Patriarca, 2016; Bacha et al., 2023; He et al., 2024).
Debido al efecto negativo que pueden causar las especies de Alternaria en la salud de la planta y por ende en la salud humana, es de suma importancia explorar nuevas estrategias de manejo de especies patógenas; no obstante, en México la información al respecto es escasa, por lo que el objetivo de esta revisión fue describir la situación actual del género Alternaria en México mediante una recopilación de estudios publicados acerca de las especies fitopatógenas reportadas, las enfermedades que causan en plantas, así como las diversas estrategias de control.
Marco teórico y experimental
Análisis bibliográfico. Se realizó una revisión sistemática de la literatura científica obtenida a la fecha de la redacción de esta revisión sobre caracterización, enfermedades asociadas y estrategias de manejo de especies del género Alternaria en México. Para la búsqueda de artículos científicos en inglés y español se consultaron bases de datos como: PubMed, Redalyc, Scopus, SciELO, Google Scholar. La consulta se realizó considerando publicaciones científicas durante el periodo de 1990 a 2025. Las palabras clave utilizadas incluyeron “Alternaria” AND “identificación” AND “México”, “Alternaria” AND “enfermedad” AND “México”, “Alternaria” AND “control” AND “México”.
Caracterización morfológica de Alternaria. Alternaria fue descrito originalmente por Ness en 1816, con la especie tipo A. tenuis. Las descripciones iniciales fueron recopiladas en la clave taxonómica de Simmons (2007) en la que se reconocieron 275 especies. El hongo se caracteriza por tener colonias de color gris, marrón y negro, con hifas incoloras, de color marrón u oliva; ausencia de setas y rara vez forma estroma. Los conidióforos son de color marrón, solitarios o en fascículos con conidios ovoides, cilíndricos, elipsoides u obclavadas de color olivo a marrón, presentan septos transversales y pueden presentar o no, septos longitudinales u oblicuos; en promedio los conidios presentan medidas de 45(-70.5) × 6.5-15.5 (-17) µm (Figura 1) (Watanabe, 2002; Simmons, 2007; Woudenberg et al., 2013).
La caracterización morfológica de Alternaria, inicia con el aislamiento y siembra en medios selectivos como Agar Extracto de Malta (MEA), Agar Sabouraud y no selectivos como Papa Dextrosa Agar (PDA), Agar Papa Zanahoria (PCA), Agar Harina de Maíz (CMA) y Agar V8 (Rivas y Mhulhauser, 2014; Blagojevic et al., 2020; Yessimseitova et al., 2025).Las características morfológicas macroscópicas que se consideran para su identificación son: color de la colonia, margen de la colonia, textura y forma del micelio. Para las características microscópicas se requiere observación directa mediante microscopía óptica, donde se considera la forma de conidióforos, células conidiógenas y conidios, ramificación y disposición de conidióforos, disposición de septos, clamidosporas y, finalmente se requieren las características morfométricas como el tamaño de conidios, número de septos transversales y longitudinales, longitud de conidióforos y clamidosporas (Barnett y Hunter, 1998; Simmons, 2007).
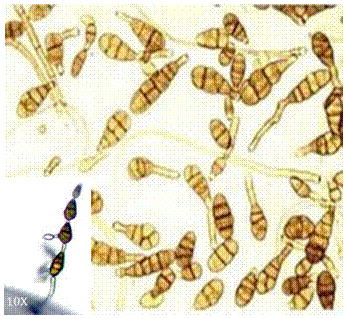
De acuerdo con los caracteres morfológicos de este género, anteriormente se clasificaron en dos categorías, de esporas grandes y esporas pequeñas, estas últimas incluyen a la sección Alternaria, donde se encuentran A. arborescens y A. alternata (De Mers, 2022). Se han adoptado los términos de formae specialis y patotipos para algunas de estas especies, en el caso de A. alternata, se han descrito por lo menos siete patotipos, ya que cada uno genera toxinas específicas dependiendo del hospedero (Akimitsu et al., 2014). Actualmente, las características morfológicas no son suficientes para la identificación de organismos debido a la dificultad de identificar las especies debido a los polimorfismos intraespecíficos y similitudes interespecíficas, agravadas por su sensibilidad a las variaciones ambientales (Ghoneem et al., 2023).
Caracterización molecular de Alternaria . En la actualidad se requiere del uso de técnicas de análisis de ADN como la técnica basada en PCR y secuenciación masiva, además de la amplificación de genes multilocus y análisis filogenético para caracterizar molecularmente a las especies de Alternaria (Ghoneem et al., 2023). Los principales genes multilocus empleados como biomarcadores para la identificación molecular de especies de Alternaria son: La subunidad nuclear pequeña del ARN ribosomal (SSU), la subunidad nuclear grande del ARN ribosomal (LSU), el espaciador interno transcrito (ITS), Gliceraldehido-3-fosfato- deshidrogenasa (GAPDH), ARN polimerasa II, la segunda subunidad mayor de la ARN polimerasa II (RPB2), Factor de elongación 1 alfa (TEF 1 alfa), Alergeno mayor de Alternaria (Alt A1), Endopoligalacturonasa (Endo PG), Región genética anónima (OPA 102), Calmodulina (CAL) y grupo ortólogo eucariota (KOG) (Woudenberg et al., 2015; Jayawardena et al., 2019).
La filogenia molecular es una valiosa herramienta para dilucidar taxones parafiléticos dentro de clados de Alternaria, esto implica que no siempre existe una correlación entre los grupos de especies basados en las características morfológicas (He et al., 2024). Por lo tanto, se emplean distintos métodos para determinar las relaciones filogenéticas de las especies estudiadas como por ejemplo Máxima Parsimonia, Máxima verosimilitud y/o Inferencia Bayesiana (Tovar-Pedraza et al., 2024). Debido al desafío de distinguir morfoespecies con base en secuencias génicas, actualmente se pueden aplicar tecnologías como la secuenciación del genoma completo para buscar genes que puedan servir como biomarcadores que permitan distinguir estas especies (Dang et al., 2015). En el Centro Nacional para la Información Biotecnológica (NCBI) se encuentran dos genomas de Alternaria disponibles públicamente: A. brassicicola, secc. Brassicicola (BioProject PRJNA34523), y A. arborescens, secc. Alternaria (BioProject PRJNA78243) (Woudenberg et al., 2015).
Complejo de especies de Alternaria. Los géneros Alternaria, Chalastospora, Crivellia, Embellisia, Nimbya, Stemphylium, Ulocladium, Undifilum y Sinomyces se encuentran dentro del complejo de especies de Alternaria, las cuales están estrechamente relacionadas; sin embargo, después de una revisión taxonómica, todos los géneros resultan en sinonimia con Alternaria, a excepción de Stemphylium. Dentro de este complejo de especies se encuentran especies saprófitas, endófitas y patógenas (Woudenberg et al., 2013). Por otro lado, debido a actualizaciones taxonómicas, por estudios moleculares, especies de Chalastospora han cambiado a Alternaria (e.g. Alternaria cetera syn. Chalatospora cetera). La sección Chalatospora se caracteriza por tener conidióforos simples o ramificados con conidios de color marrón, elipsoides u ovoides, ausencia de septos transversales, y rara vez septos longitudinales, estos se observan solos o en cadena (Woudenberg et al., 2013), se encuentra asociado a dermatitis granulomatosa nodular en animales domésticos y especies vegetales (Norris et al., 2021).
La sección Crivellia se caracteriza por presentar conidióforos primarios rectos o curvos, simples o ramificados con conidios rectos, curvos, con septos transversales, pueden presentar microesclerocios o clamidosporas (Woudenberg et al., 2013; Lawrence et al., 2016). Dentro de esta sección se encuentra A. penicillata, estado sexual de Brachycladium penicillatum.
Dentro de la sección Embellisia se encuentra A. embellisia, causante del cancro de bulbo en ajo (Allium sativum), se describen los conidióforos como simples o ramificados con conidios de color marrón con dos a seis septos transversales y uno o dos longitudinales y clamidosporas en pares o cadenas (Delgado-Ortiz et al., 2019). En el caso del género Nimbya, este se encuentra dentro de la sección Alternantherae, con especies que actualmente son del género Alternaria (e.g. A. alternantherae syn. N. alternantherae) (Woudenberg et al., 2013), sin embargo, existe la sección Nimbya, en la que se encuentra A. cypericola, que presenta hifas ramificadas y septadas, con conidios rectos o curvos, ovoides a elipsoides, de color marrón, generalmente solitarios o pueden formar cadenas de dos a tres conidios, los conidióforos son macronematosos, solitarios, rectos o ligeramente curvos con septos y de color marrón; se encuentra asociadas a plantas de la familia Juncaceae y Cyperaceae (Ahmadpour et al., 2021). El género Stemphylium se encuentra en diferentes secciones del complejo de especies de Alternaria, este se encuentra asociado a enfermedades en cultivos como cebolla, causando lesiones de color marrón en las hojas, provocando defoliación y pérdida de productividad por su incapacidad fotosintética (Hay et al., 2021). Este género es el estado anamorfo de Pleospora spp. (Simmons, 1985). La especie S. vesicarium presenta hifas con septos, ramificadas, hialinas, con paredes lisas, con conidióforos solitarios, simples, rectos o ligeramente curvados, lisos y rugosos en la célula apical, con conidios de color marrón, oblongos, ovalados, redondeados en el ápice y verrugosas (Kádasi et al., 2024).
Ulocladium, es un género que de acuerdo con Kidd y colaboradores (2022), incluye especies saprófitas, que afectan a frutos y causante de enfermedades en plantas, raramente afecta a humanos. Las colonias pueden formarse de color negro a oliva o gris, con conidióforos geniculados que genera conidios multicelulares que generalmente se encuentran solos, de forma obovada, de color marrón y paredes rugosas. Se ha probado la especie U. atrum como antagonista de Sclerotinia sclerotiorum, con capacidad de crecimiento sobre el patógeno, sin embargo, no parasitó los esclerocios (Li et al., 2003). La sección Undifilum incluye especies endófitas, los conidios presentes son ovadas a elipsoides largos, los septos formados pueden ser gruesos, comparte caracteres con Embellesia; sin embargo, se caracteriza por la formación de tubos germinativos ondulados que se ramifican, además, producen el compuesto tóxico swaisonina (Woudenberg et al., 2013), se puede encontrar en Astragalus sp. y Oxytropis sericea (Cook et al., 2009; Baucom et al., 2012).
Distribución y principales síntomas causados por Alternaria en México
Se han reportado diversas especies de Alternaria de interés agrícola y ornamental, causando enfermedades como mancha y tizón foliar (Palemón-Alberto et al., 2024). Dentro de las especies afectadas se encuentra la cebolla (Reyes-Tena et al., 2023), crisantemo (Chrysanthemum morifolium) (Domínguez-Serrano et al., 2016) y papayo (Carica papaya) (Vásquez-López et al., 2012). En cactáceas se asocia a la enfermedad de la mancha dorada y mancha negra (Montiel-Salero et al., 2022; Chavarría-Cervera et al., 2024). Oficialmente se conocen al menos 10 especies diferentes de Alternaria en México en 23 hospederos de importancia económica, que están distribuidos en 15 estados (Cuadro 1). No obstante, se tiene desconocimiento de su amplia distribución del hongo en cultivos de interés, así como en hospedantes alternos.
Cuadro 1 Especies fitopatógenas de Alternaria reportadas en cultivos en México.
| Especie reportada | Hospedero | Área geográfica | Referencia |
|---|---|---|---|
| A. alternata, A. tenuissima | Chrysanthemum morifolium | Yucatán | Dominguez-Serrano et al., 2016 |
| A. limicola | Citrus paradisi, C. aurantium, C. macrophylla y C. sinensis | Colima | Palm y Civerolo, 1994 |
| A. alternata | Triticum spp. | Sonora | Mata-Santoyo et al., 2018 |
| A. tomato | Helianthus annuus | Yucatán | Poudel et al., 2019 |
| A. alternata | Fragaria×ananassa | Guanajuato | Mariscal-Amaro et al., 2017 |
| A. alternata | Carica papaya | Guerrero | Vásquez-López et al., 2012 |
| A. tenuissima, A. alternata | Brassica oleracea | Guanajuato | Fraire-Cordero et al., 2010 |
| A. alternata | Opuntia matudae | Hidalgo | Montiel-Salero et al., 2022 |
| A. alternata | Avena sativa | Tlaxcala | Leyva-Mir et al., 2014 |
| A. embellisia | Allium sativum | Coahuila | Delgado-Ortiz et al., 2019 |
| A. alternata | Brassica oleracea var. italica | Guanajuato | Arratia-Castro et al., 2022 |
| A. solani | Solanum lycopersicum | Chiapas | Quiroga-Madrigal et al., 2007 |
| A. tenuissima | Salix bonplandiana | Estado de México | González-Díaz et al., 2011 |
| A. chrysanthemi | Chrysanthemum morifolium | Yucatán | Villanueva-Couoh et al., 2004 |
| A. alternata | Jatropha curcas | Sinaloa | Espinoza-Verduzco et al., 2012 |
| A. alternata | Solanum lycopersicum | Sinaloa | Félix-Gastélum y Gálvez-Figueroa, 2002; Troncoso- Rojas et al., 2005 |
| A. tenuissima | Malus x domestica | Chihuahua | Madrid-Molina et al., 2023 |
| A. solani | Capsicum chinense | Yucatán | Cristóbal et al., 2006 |
| A. alternata | Allium cepa | Michoacán | Reyes-Tena et al., 2023 |
| A. alternata | Ficus carica | Morelos | Saavedra et al., 2020 |
| A. alternata | Opuntia ficus-indica | Colima | Chavarría-Cervera et al., 2024 |
| A. arborescens | Capsicum pubescens | Guerrero | Palemón-Alberto et al., 2024 |
| Alternaria sp. | Vaccinium sp. | Michoacán | Mondragón-Flores et al., 2012 |
| A. solani,A. alternata | Capsicum annuum | Chihuahua | Guigón-López et al., 2001 |
| A. alternata | Cyamopsis tetragonoloba | Sinaloa | García-León et al., 2024 |
| Alternaria spp. | Capsicum annuum | Sinaloa | Guerrero-Santana y Vega-Camargo, 2024 |
| A. burnsii A destruens | Plumeria obtusa | Sinaloa | Márquez-Licona et al., 2025 |
| A. alternata | Vaccinium corymbosum | Sinaloa | Nuñez-García et al., 2025 |
Su importancia de este hongo depende de la incidencia y severidad que puede generar en los diferentes cultivos. Por ejemplo, se ha determinado la incidencia de la mancha foliar causada por Alternaria en cultivos de crisantemo de hasta el 30 %, con síntomas en forma de manchas en el margen de las hojas de forma circular u ovaladas de color café a negro con halos amarillos y otras hojas completamente amarillas (Domínguez-Serrano et al., 2016). El tizón foliar en cultivo de cebolla (Allium cepa), se reporta incidencia del 20 % en verano, con síntomas en forma de manchas de color café y morado en las hojas, mientras que en invierno la incidencia puede llegar hasta el 80 %, con manchas foliares moradas y lesiones secas. Estos síntomas mercan el crecimiento del bulbo (Reyes-Tena et al., 2023).
Sinaloa, es un estado predominante por su alta producción de cultivos como maíz, frijol, chile y jitomate, el cual ha registrado problemas por este hongo. Se ha reportado el moho negro en frutos maduros de tomate (Solanum lycopersicum), causada por A. alternata, donde la severidad varía de acuerdo a las condiciones de humedad prevalecientes durante la madurez en frutos. Cuando se presentan períodos prolongados de rocío, lluvia o alta humedad relativa, los conidios de A. alternata germinan en respuesta a nutrientes solubles presentes en la superficie de los frutos. Los síntomas de enfermedad varían desde pequeñas lesiones superficiales de color café claro hasta lesiones necróticas hundidas, y se produce una capa negra de conidios sobre el tejido infectado (signo). Esto representa una pérdida económica para los productores cuando la incidencia de la enfermedad supera el 8 % (Félix-Gastélum y Gálvez-Figueroa, 2002). Adicionalmente, se ha reportado A. alternata ocasionando pudrición blanda y lesiones hundidas en frutos de tomate, mancha foliar en guar (Cyamopsis tetragonoloba) (Félix- Gastélum y Gálvez-Figueroa, 2002; García-León et al., 2024); Alternaria spp. También ocasiona mancha foliar en ajonjolí (García-Espinoza, 2022) y pudrición de fruto en chile (Guerrero-Santana y Vega-Camargo, 2024).
Alternaria no solo afecta el follaje y frutos de los cultivos, también puede causar el tizón de inflorescencias en Jatropha curcas ocasionada por A. alternata en Sinaloa, donde se observan lesiones pequeñas de color marrón oscuro que al fusionarse forman lesiones necróticas más grandes, lo que provoca la caída de las flores (Espinoza-Verduzco et al., 2012). En almácigos y semillas de chile (Capsicum annuum) induce la secadera temprana (damping-off) en almácigos y semillas en Aguascalientes y Zacatecas, México, con asociaciones con Alternaria, Fusarium y Rhizoctonia (Velásquez-Valle et al., 2007).
Por otro lado, las malezas son de importancia al ser hospedantes de este hongo al encontrarse asociados o cerca de cultivos de relevancia económica. Por ejemplo, en Sinaloa se ha reportado el tizón foliar (Alternaria spp.) en tabaco silvestre (Nicotiana glauca) el cual crece al costado de los canales de riego y drenes. En las hojas se observan síntomas como lesiones irregulares de color café claro a oscuro en forma irregular. La severidad es mayor cuando los periodos diarios de follaje mojado varían de 15 a 18 h y la temperatura oscila entre 14 y 28 °C. Por su parte, especies de Alternaria de la Sección Alternata se relacionan con el tizón foliar en tabaquillo (Nicotiana trigonophylla) y chichiquelite (Solanum nigrum) y Alternaria ricini se ha asociado como el tizón foliar en higuerilla silvestre (Ricinus comunis) y lesiones foliares en tabacón (Nicotiana glauca) y tabaco silvestre (Figura 2D-E) (Félix-Gastélum et al., 2023).
Generalmente, este hongo se ha caracterizado por síntomas característicos; sin embargo, se ha reportado con asociaciones con otros hongos, como la mancha negra en nopal (Opuntia ficus-indica) es causada por una asociación de Alternaria alternata, Corynespora cassiicola y Neocytalidium dimidiatum; entre estos, A. alternata, ha registrado incidencia de hasta el 60 %, con síntomas de ablandamiento de los cladodios, así como lesiones de color negro de forma circular e irregular (Figura 2C) (Chavarría-Cervera et al., 2024). El mismo patógeno es causante de la mancha dorada, descrita en México en O. matudae (Xoconostle), que genera lesiones en cladodios de color amarillo o clorosis cerca de las areolas, que posteriormente se forma una mancha dorada que evoluciona a necrosis del tejido (Figura 2A) (Montiel-Salero et al., 2022).

Figura 2 Diversidad de enfermedades asociadas a Alternaria spp. A) Síntomas de mancha dorada en Opuntia matudae causadas por A. alternata (Tomado de Montiel-Salero et al., 2022); B) Síntoma de pudrición del florete de brócoli por A. alternata (Tomado de Arratia-Castro et al., 2022); C) Síntoma de mancha negra en Opuntia ficus-indica por A. alternata (Tomado de Chavarría-Cervera et al., 2024); D) Síntomas causados por Alternaria sp. en tabaco silvestre y E) en tabaco (Nicotiana glauca) (Tomado de Félix-Gastélum et al., 2023).
Otro caso de asociación con otros hongos es la pudrición del florete en el cultivo de brócoli (Brassica oleracea var. italica), que reduce la calidad y rendimiento del cultivo, cuando se tiene alta humedad relativa (70 %) y temperatura de 30 °C, las pérdidas superan el 40 % (Arratia-Castro et al., 2022). Alternaria tenuissima, A. alternata y Fusarium oxysporum se asociaron como los causantes de la enfermedad en variedades comerciales Marathon, Patron y Monaco en Guanajuato (Fraire-Cordero et al., 2010). Mientras que Arratia-Castro et al. (2022) reportaron la asociación de A. alternata, F. verticillioides y F. oxysporum como agentes causales de la pudrición del florete de brócoli (Figura 2B).
Estrategias de manejo
Para establecer medidas de control, se requiere hacer un diagnóstico del agente causal de la enfermedad con el fin establecer estrategias de manejo acorde al cultivo. Con el objetivo de realizar un manejo integrado, al menos se establecen tres técnicas, las cuales se basan en el control químico, cultural, físico, biológico. De este último, como el uso de agentes de control biológico, extractos vegetales y microalgas. Aunque el control químico es la alternativa más eficaz, el uso inadecuado de los mismos ha ocasionado contaminación en los agroecosistemas, aunado al riesgo en salud pública por el manejo de estos productos (Zavaleta-Mejía, 1999; Zepeda-Jazo, 2018).
Control químico. Es uno de los métodos más utilizados por productores agrícolas por tener una gran eficacia y resultados rápidos. Los principales grupos químicos utilizados para el control de Alternaria spp. son los triazoles, en este grupo se encuentra el propiconazol, este fungicida sistémico inhibe el crecimiento radial micelial y la germinación de los conidios, al igual que la mezcla de azoxystrobin + ciproconazol, del grupo de los metoxiacrilatos y triazoles (Ramírez-Jiménez et al., 2025). La mezcla de ambos grupos químicos también se evaluó con azoxystrobin + epoxiconazol, que retrasa la senescencia de plantas de trigo e indujo un aumento de la enzima antioxidante superóxido dismutasa y reducción de los niveles de O2 (Wu y Tieddmann, 2001). Del grupo de las carboxamidas, Captan inhibe el crecimiento micelial al interferir en la respiración celular, dificultando el desarrollo del micelio y se transloca a diversos tejidos a través del tratamiento a semillas o suelo; así como los ditiocarbamatos, mancozeb, modifica e inactiva las proteínas involucradas en la traducción y transcripción del ADN (Zarate-Ramos et al., 2022). Los cloronitrilos (Clorotalonil), reducen el crecimiento micelial ya que este compite con el gliceraldehido-3-fosfato por el sitio activo de la gliceraldehido-3-fosfato deshidrogenasa y reduce las moléculas de glutatión fúngico a formas alternativas; así mismo se ha observado la degradación a subproductos que se unen a sitios objetivo (Fairchild et al., 2013).
Aunque en algunos casos se emplea la combinación de fungicidas del mismo modo de acción para el mismo cultivo o patógeno, estudios han demostrado que especies de Alternaria han mostrado resistencia cruzada con mancozeb, tebuconazol, iprodiona, fludioxonil y ciprodinil en Grecia (Malandrakis et al., 2015) y mancozeb y difenoconazol en China, sugiriendo que el uso de fungicidas no específicos del sitio puede generar menor sensibilidad en los patógenos y mayor capacidad de generar enfermedades, lo que representa un riesgo en la producción agrícola (Yang et al., 2019).
También se utiliza el fungicida sistémico Amistar GS® (azoxystrobin) en 500 ppm contra
A. chrysanthemi, el cual puede reducir intensidad del daño foliar en un 50 % y además de asociarse con una menor tasa de infección con respecto a otros fungicidas sintéticos (Villanueva-Couoh et al., 2004). En frutos postcosecha e in vitro se ha utilizado el isotiocianato de bencilo, con resultados favorables al utilizar 0.1 mg mL-1 y 0.56 mg mL-1; siendo un controlador eficiente en frutos sin afectar la calidad (Troncoso-Rojas et al., 2005). No obstante, es importante destacar del tiempo de aplicación de los agroquímicos en la floración o formación del fruto y la permanencia de estos en los frutos para evitar la residualidad en poscosecha.
De la misma manera, se han evaluado in vitro Fosetyl aluminio (800 ppm), Azoxystrobin (500 ppm), Imazalil (750 ppm), Procloraz (450 ppm), Benomilo (500 ppm) y Thiabendazol (600 ppm); sin embargo, solo se demostró que Imazalil y Procloraz presenta una efectividad al 100 % sobre Alternaria sp. (Herrera et al., 2011). Esto sugiriere la posible resistencia de especies de Alternaria contra algunos agroquímicos. Por ello, es pertinente estudiar en el país sobre la resistencia que han adquirido las especies de Alternaria de los productos disponibles en el mercado.
En Sinaloa se evaluó in vitro Azoxystrobin a dosis de 7 y 10 ppm, aunque inhibió hasta 99.5
% la germinación de conidios de Alternaria alternata no redujo el crecimiento micelial (Félix- Gastélum y Gálvez-Figueroa, 2002). No obstante, el control químico no es amigable con el ambiente por lo que genera contaminación ambiental y daño a la salud humana a los consumidores por residuos tóxicos en los alimentos y en los trabajadores que se encuentran en contacto directo con los productos químicos (Rangel-Ortíz et al., 2023), por lo que es pertinente la interacción de otras estrategias de manejo.
Control químico basado en la fenología del cultivo. Si bien en México existe escasa información sobre sistemas de predicción basados en la fenología del cultivo, variables ambientales y densidades del inóculo para el inicio de aplicaciones preventivas de los fungicidas químicos en el manejo de enfermedades causadas por especie de Alternaria, existe información en otros países sobre el uso eficiente de fungicidas. En México se han determinado etapas de mayor incidencia y severidad causados por Alternaria en cultivos de cebolla; sin embargo, existe escasa información sobre la aplicabilidad de esta información (Reyes-Tena et al., 2023). En Israel se determinaron los periodos de mayor susceptibilidad de A. alternata en el cultivo de manzana mediante el desarrollo de una estrategia de manejo fenológico, al sincronizar las aplicaciones de fungicidas con la etapa máxima de susceptibilidad del fruto es una estrategia efectiva para controlar la pudrición de Alternaria, con esto se puede optimizar el uso de fungicidas, reducir costos y minimizar problemas de resistencia (Lior et al., 2018).
Por otra parte, es importante considerar la carga de inóculo en el manejo preventivo con fungicidas químicos. Por ejemplo, la carga de inóculo influye en la severidad de la mancha foliar causada por A. brassicae en el cultivo de coliflor; para un manejo efectivo de la enfermedad es importante controlar la cantidad de inóculo en el suelo, el monitoreo constante y tomar en cuenta la carga de inóculo como factor clave para interpretar la eficacia del control químico (Sunitha y Jha, 2023).
Control biológico. Diversos microorganismos se han evaluado como agentes de control biológico de hongos fitopatógenos, el género más estudiado es Trichoderma, en México las principales especies que se ha reportado como biocontrol contra especies fitopatógenas de Alternaria son T. harzianum, T. asperellum y T. viride (Allende-Molar et al., 2022). Este género tiene mecanismos para controlar el crecimiento y desarrollo de fitopatógenos como: 1) antibiosis, mediante metabolitos secundarios que alteran la permeabilidad de las membranas lipídicas de los fitopatógenos, 2) competencia por espacio y nutrientes, por su rápido crecimiento logra colonizar espacios por los que compiten otros microorganismos e incluso, coloniza los sitios de unión de los fitopatógenos a la planta, 3) micoparasitismo, a través del crecimiento quimiotrófico de Trichoderma, se adhiere y enrolla al patógeno, induce la producción de enzima líticas extracelulares (quitinasas, glucanasas y proteasas) que degradan la pared celular del microorganismo patógeno, e 4) inducción de resistencia, la activación de mecanismos de resistencia sistémica aceleran la respuesta frente a fitopatógenos al segregar proteínas con actividad enzimática (Companioni et al., 2019)
La evaluación in vitro es la vía principal para probar preliminarmente un organismo de control biológico. Se han realizado distintos ensayos de antagonismo con diversas especies de Trichoderma, por ejemplo se evaluó a T. asperellum contra Alternaria sp., mostrando un porcentaje de efectividad entre 50 y 93 % (Matas-Baca et al., 2022), y la evaluación in vivo del mismo antagonista (100 mL) en árboles de manzana, que mostró capacidad antagónica sobre el control (Madrid-Molina et al., 2023), se le atribuye su efectividad a la producción enzimática de glucanasas y quitinasas, que influyen en la respuesta de defensa contra Alternaria spp., degradando la pared celular (Infante et al., 2009; Camacho-Luna et al., 2021). En los últimos años se han realizado varias investigaciones enfocadas en el uso de agentes de control biológico para el control de una amplia diversidad de hongos fitopatógenos; no obstante, los estudios encausados en el control de Alternaria, aún son limitados (Cuadro 2).
Cuadro 2 Efectividad in vitro de microorganismos antagonistas contra Alternaria spp.
| Alternaria spp. | Microorganismo antagonista | Porcentaje de efectividad (%) | Referencia |
|---|---|---|---|
| Alternaria solani | Trichoderma harzianum | 62.9-73.8 | Michel-Aceves et al., 2008 |
| A. solani | Trichoderma longibrachiatum | 44.2-67.8 | Michel-Aceves et al., 2008 |
| A. solani | Trichoderma koningii | 81.3 | Michel-Aceves et al., 2008 |
| A. alternata | Trichoderma asperellum | 56-62 | Rios-Velasco et al., 2016 |
| A. alternata | Bacillus methylotrophicus | 63 | Rios-Velasco et al., 2016 |
| A. alternata | Bacillus amyloliquefaciens | 64 | Rios-Velasco et al., 2016 |
| A. alternata | Bacillus subtilis | 38.2-71.9 | Ruiz-Sánchez et al., 2016 |
| A. alternata | Pseudomonas fluorescens | 47 | Rodríguez-Romero et al., 2019 |
Bacillus subtillis también es de las especies bacterianas más utilizadas como biocontroladoras de fitopatógenos, se ha probado el producto comercial Probacil® (1 X 108 de células por mL) en combinación con ácido salicílico sobre A. solani en plantas de tomate, como resultado se redujo la severidad de la enfermedad, aumentó el crecimiento de la plantas y se obtuvo mayor rendimiento en los frutos (Figura 2) (Espinosa-Vázquez et al., 2019); con B. atrophaeus, contra A. alternata, se muestra antagonismo in vitro y 33.7 % de germinación de esporas, mismo que se le atribuye a la presencia de lipopéptidos; esta cepa se comparó con Brevibacterium frigoritolerans, que no muestra inhibición suficiente; sin embargo, la germinación de conidios es tan solo del 12 % (Chacón-López et al., 2021).
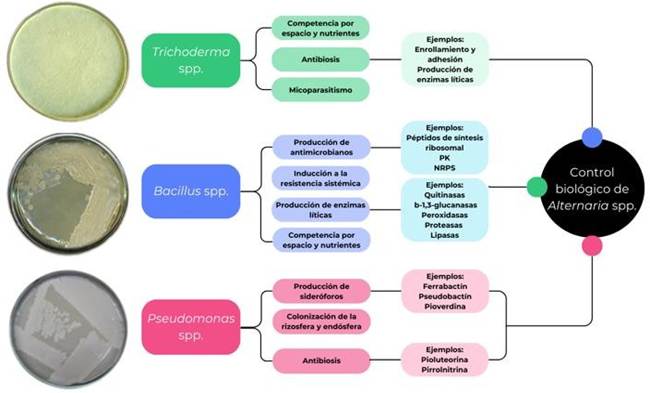
Figura 3 Mecanismos de acción de los principales agentes de control biológico (PK: Policétidos; NRPS: moléculas peptídicas de síntesis no ribosomal).
La eficacia de los agentes de control biológico se debe a la diversidad de formas de acción de estos microorganismos, mismos que se ilustran en la Figura 3 (Martínez et al., 2013; Álvarez-García et al., 2020; Pedraza et al., 2020). No obstante, el uso de microorganismos es un proceso que demora tiempo para que actúen correctamente debido al tiempo de crecimiento de estos, ya que se requiere de productos de rápida acción para el control de los patógenos; además, el costo del control biológico es más elevado que el control químico u otro tipo de control, por lo que actualmente se está trabajando en la búsqueda de nuevas alternativas para el control de fitopatógenos.
Otras alternativas de control. Aunado al control biológico, se han explorado nuevas alternativas, como el uso de extractos de plantas, aceites esenciales o biopolímeros. En el caso de los biopolímeros, es un área con gran potencial y sus estudios aún están en ascenso; sin embargo, se ha demostrado que el quitosano posee actividad antifúngica, además, proporciona un aumento en la producción de peroxidasa, catalasa y fenilalanina amonio liasa que mejoran la defensa vegetal (Rodríguez-Guzmán et al., 2019) y en combinación con biocontroladores como P. fluorescens, inhibe hasta el 60.2 % del crecimiento micelial y 100 % de inhibición de germinación de los conidios (Rodríguez-Romero et al., 2019).
Extractos vegetales. Los extractos vegetales resultan ser una alternativa benéfica como método preventivo; el uso del extracto de acetona de chilemecate (Salmea scandens), a concentraciones de 4 000 y 5 000 ppm, ha tenido resultados prometedores in vitro y en frutos de tomate contra A. solani, esto se le atribuye ya que el extracto tiene la presencia de ácidos carboxílicos, aldehídos, cetonas y compuestos aromáticos, mismos que pueden fungir como antagonistas (Salas-Marina et al., 2021). El extracto etanólico de residuos de mamey (Pouteira sapota) también muestra efecto inhibitorio de hasta el 46.4 % sobre Alternaria spp. (Rodríguez-Romero y Martinez-Ramírez, 2023). El aceite esencial de tomillo (Thymus vulgaris) (1 000 ppm) presenta capacidad antifúngica in vitro contra A. citri, causante de pudrición en cítricos, se le atribuye su capacidad antagonista a que los compuestos del aceite esencial son borneol, timol y carvacrol (Soto et al., 2006).
La elaboración de productos eficientes de extractos contra fitopatógenos es una tarea complicada; sin embargo, la constante evaluación de diferentes extractos ha dado resultados prometedores contra fitopatógenos, además, son seguros para los consumidores y para el ambiente (Villa-Martinez et al., 2015), y, aunque resultan tener una buena efectividad, los bioplaguicidas requieren regulación de instituciones como la Secretaría de Salubridad y Asistencia (SSA) en coordinación con la Secretaría de Agricultura y Desarrollo (SADER) y Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT), ya que se debe emitir un permiso para su elaboración y fabricación.
Microalgas. El uso de las microalgas ha incrementado debido a su bajo costo, fácil producción y sus características; convierten el dióxido de carbono en biomasa, misma que es utilizada para la creación de alimentos para humanos y animales, cosméticos, biocombustibles, tratamiento de aguas residuales y como bioestimulante para plantas; entre los géneros con mayor importancia comercial se encuentran Chaetoceros, Chlorella, Dunaliella e Isochrysis (Ortiz- Moreno et al., 2019). En un estudio realizado por Schmid y colaboradores (2022), evaluaron diferentes microalgas contra diversos fitopatógenos, entre estos, A. alternata, las microalgas fueron Nannochloropsis sp., Phaeodactylum tricornutum, Scenedesmus obliquus, Chlorella vulgaris y Spirulina sp., y se determinó que únicamente la microalga P. tricornutum mostró capacidad antagonista de los hongos, con excepción de A. alternata.
Nanopartículas. Las nanopartículas poseen actividad antimicrobiana y antifúngica, lo que las convierte en nanoplaguicidas más eficientes que los plaguicidas tradicionales (Wang et al., 2017). Estas partículas pueden interactuar directamente con las membranas celulares de los patógenos vegetales, causando alteraciones estructurales. Debido a su reducido tamaño, son capaces de penetrar las paredes celulares de bacterias y hongos, generando despolarización de la membrana, pérdida del contenido celular interno y, finalmente, la lisis celular (Sirelkhatim et al., 2015). Este efecto se debe a la liberación gradual de iones de zinc (Zn²⁺) al disolverse las ZnO NPs en el entorno, los cuales interfieren con procesos enzimáticos esenciales, la síntesis proteica y la replicación del ADN, afectando gravemente a las células microbianas (Mishra et al., 2025).
Se ha evidenciado la aplicación de nanopartículas de zinc (ZnO); Mishra et al. (2025) reportaron que las nanopartículas de óxido de zinc micogénico (elaboradas con filtrado de cultivo de Trichoderma harzianum) inhibió el crecimiento micelial de Alternaria brassicae en un 91.48 % a 200 μg mL-1 en comparación con las nanopartículas de zinc sintetizadas químicamente a 200 μg mL-1 (79.62%) y Mancozeb al 02 % (82.96 %). Por otra parte, las nanopartículas de plata (AgNP) también tienen potencial en la actividad antifúngica, Ansari et al. (2023) sugieren que las nanopartículas de plata tienen potencial para mejorar el crecimiento y rendimiento de plantas de tomate y a su vez brinda protección contra Alternaria solani.
Por otro lado, Cerna-Chavez et al. (2024) evaluaron in vitro el efecto inhibitorio de nanopartículas de dióxido de silicio (NPs SiO2) y grafeno (NPs-Graf) mezcladas con extractos de Bacillus amyloliquefaciens (EcBa) sobre el desarrollo micelial y formación de estructuras reproductivas de A. alternata; el tratamiento con nanopartículas de silicio presentó mejor efecto inhibitorio ya que logró inhibir el crecimiento micelial y disminuyó la producción de esporas y esclerocios de 84 hasta 100 %.
En México existen escasos estudios sobre el uso de nanopartículas contra especies de Alternaria. Hernández-López et al. (2018) desarrollaron nanopartículas de quitosano con α- pineno (P-CSNP) y un recubrimiento comestible nanoestructurado (EC-PCSNP) y evaluaron estas nanopartículas en pimiento morrón (Capsicum annuum) inoculado con A. alternata en refrigeración para evaluar su calidad poscosecha; observaron que la aplicación de nanopartículas de quitosano previno la incidencia y la severidad del fitopatógeno durante 21 días de almacenamiento.
Conclusiones y Perspectivas
El complejo de especies de Alternaria abarca diferentes especies que representan un riesgo para los cultivos, plantas ornamentales y al ser productoras de micotoxinas, son un riesgo potencial en salud humana. Su taxonomía es compleja, y actualmente el análisis filogenético está en constantes cambios. Alternaria spp. están ampliamente estudiadas en cuanto a importancia económica, en esta revisión, se detallan 10 especies en 23 cultivos, distribuidos en diferentes estados de México, lo que muestra la gran adaptabilidad a diferentes condiciones ambientales que tiene este género. Alternaria genera daños de hasta el 80 % del total en cultivos, de ahí la importancia del estudio de estos organismos; sin embargo, aún es probable que aún haya hospedantes cultivables por reportar y otras especies vegetales que pueden funcionar como reservorio del patógeno, tal es el caso de las plantas ornamentales y especies nativas, que al no generar un impacto económico directo, no son foco de atención por parte de la comunidad científica, que a su vez, pueden llegar a generar un impacto ecológico mayor.
Debido a la importancia de generar estrategias de control sobre Alternaria spp., se incluyen la utilización de control químico; no obstante, estos causan consecuencias negativas en los consumidores del cultivo, y al aplicarlo, pueden afectar de manera colateral a microorganismos benéficos del suelo, además, el manejo adecuado de las enfermedades podría reducir la capacidad de los patógenos de generar estructuras de resistencia y permanecer latentes en el suelo.
La utilización de métodos preventivos como el uso de productos biológicos y naturales y las prácticas agrícolas como la rotación de cultivos, desinfección de maquinaria y materiales podrían reducir la incidencia y severidad de los patógenos. El uso de control biológico como una estrategia contra fitopatógenos, resulta ser una vía favorable para mejorar la calidad del suelo que repercute positivamente en la salud vegetal, aunque suelen ser métodos con acción a corto y mediano plazo, tienen mejores beneficios a largo plazo. En los ejemplos presentados en esta revisión que son más utilizados en la agricultura, como Trichoderma spp. y Bacillus spp., que además de ser antagonistas eficaces, tienen la capacidad de fijar elementos esenciales para los cultivos, son promotores del crecimiento vegetal e inducen a la resistencia sistémica de las plantas hacia los patógenos (González-León et al., 2022). Esto se ha probado en diversos estudios aplicados al control in vitro e in vivo contra Alternaria spp. (Ríos-Velasco et al., 2016), con resultados prometedores y de aplicación directa para los productores (González-Chingate et al., 2020). Además, se han explorado otras alternativas naturales, como el uso de extractos vegetales o de microalgas, aceites esenciales y el quitosano, ya que estos producen compuestos que le confieren su capacidad antagónica, además, no dañan la microbiota del suelo.
La aplicación del control biológico y orgánico para controlar Alternaria spp., o cualquier otro fitopatógeno presenta dificultades debido al diagnóstico incorrecto de los patógenos, además, aún existen vacíos en cuanto a la información existente de las especies reportadas y a las costumbres sobre las prácticas agrícolas; por lo tanto es imperativo enfatizar la utilización de estrategias preventivas en el área agronómica con productos biológicos o naturales ya que tendría mejores resultados en cuanto a la fitosanidad y por ende, a la salud humana.
El complejo de especies de Alternaria tiene constantes cambios taxonómicos, incluye especies patógenas que representan pérdidas económicas en cultivos agrícolas. Oficialmente, se han reportado nueve especies en 23 hospederos cultivables; sin embargo, aún existen vacíos en distintos cultivos, en especies ornamentales, forestales y nativas. Las dificultades sobre el control adecuado de las enfermedades causadas por Alternaria spp. y otros patógenos, surge debido al diagnóstico incorrecto de las enfermedades. En esta revisión, se dilucidan los tipos de control con algunos ejemplos para México y el estado actual sobre la utilización y avances sobre vías alternas al control químico, a través de pruebas de antagonismo mediante microorganismos benéficos como Trichoderma spp., Bacillus spp., Brevibacterium spp. y Pseudomonas fluorescens, que son especies ampliamente estudiadas y viables para utilizarse como agentes de control biológico, debido a sus interacciones benéficas, además del uso de control natural con quitosano, extractos vegetales y de microalgas, que presentan antagonismo sobre Alternaria spp., y actualmente el uso de nanopartículas, que pueden ser utilizados como métodos preventivos para el control de enfermedades.

