











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Sclerotium rolfsii es un hongo fitopatógeno presente en el suelo, considerado como el agente causal de la pudrición de los tubérculos de papa (Solanum tuberosum). También provoca la pudrición de tallos, raíces, pecíolos, hojas y frutos en diversos cultivos. El hongo ha causado pérdidas de hasta del 15 % de la producción en el cultivo de papa en Italia (Garibaldi et al., 2007). Los síntomas inician con pudrición seca en la base del tallo, además necrosis en el tejido vascular; las hojas de las plantas infectadas presentan clorosis, conforme la enfermedad avanza las plantas se marchitan y los tubérculos se pudren. El hongo produce micelio blanco en forma de abanico, sobre el tejido afectado, cuando este madura se producen esclerocios, los cuales constituyen la principal fuente de inóculo primario, y se dispersan por medio del agua, residuos vegetales, suelo y animales (Kator et al., 2015; Roca et al., 2016). En Sinaloa, México, la enfermedad ha sido subestimada por los productores de papa, pero se estima que ésta ha causado pérdidas de hasta 20 % de la producción en predios con suelos que retienen alta humedad (arcillosos) y coinciden con el incremento de la temperatura ambiental superiores a los 30 °C, además los daños se incrementan cuando el productor pospone la cosecha, lo que propicia que el hongo incremente el daño a los tubérculos que permanecen en el suelo.
Los fungicidas sintéticos se utilizan para combatir diferentes especies de hongos fitopatógenos; no obstante, la aplicación excesiva de plaguicidas ha ocasionado problemas ambientales y de salud humana (Brauer et al., 2019). Por otro lado, para el control de S. rolfsii se han utilizado otros métodos con enfoque ecológico, como es el uso de agentes de biocontrol, lo cual resulta en una prometedora alternativa (Mishra et al., 2015; Khan et al., 2020). En este sentido, las diferentes especies de Trichoderma son las más utilizadas como biocontroladores, ya que presentan diferentes mecanismos de acción (competencia por el sustrato, micoparasitismo y antibiosis) que permiten manejar diversas enfermedades causadas por fitopatógenos (Infante et al., 2009; Adnan et al., 2019; Garrido y Vilela, 2019; Andrade-Hoyos et al., 2020; Pérez et al., 2020; Asad, 2022). Aun cuando estudios de antagonismo in vitro entre especies de Trichoderma y S. rolfsii en papa son nulos; otros estudios en otros hospedantes se han consignado buen control al utilizar Trichoderma spp. como antagonista contra S. rolfsii obtenidos de plantas de tomate (Solanum lycopersicum), frijol (Phaseolus vulgaris), repollo (Brassica oleracea), betel (Piper betle), lenteja (Lens esculenta), garbanzo (Cicer arietinum), calabaza (Cucurbita pepo), entre otros (Díaz-Nájera et al., 2018; Kushwaha et al., 2018;Chandra-Sekhar et al., 2020; Kamel et al., 2020; Andrade-Hoyos et al., 2023). T. hamatum, T. harzianum, T. viride, T. virens y T. asperellum producen distintos metabolitos secundarios, que activan los mecanismos de defensa, promueven el desarrollo y crecimiento de las plantas; además de inhibir el desarrollo de los patógenos (Lee et al., 2016; Chandra et al., 2020; Vineela et al., 2020; Safari et al., 2022; Guzmán et al., 2023). Los metabolitos que producen pueden ser volátiles y no volátiles; entre los metabolitos volátiles se encuentran el ácido acético, 2-pentilfuran, 6-pentil-2H-piran-2-ona, 2-metil-1-butanol entre otros, los cuales inhiben a S. rolfsii (Phoka et al., 2020; Sridharan et al., 2020).
En México, S. rolfsii ha mostrado altos niveles de incidencia en lotes comerciales de papa de Sonora y Sinaloa; sin embargo, la evaluación de efectividad biológica de las especies de Trichoderma contra este fitopatógeno bajo condiciones controladas, invernadero y campo, no se ha determinado, por lo que es importante generar alternativas ecológicas para el manejo de este hongo en el corto plazo. Por lo anterior, los objetivos de este estudio fueron determinar la capacidad antagónica in vitro de 26 aislados de Trichoderma mediante confrontaciones duales, así como, identificar las interacciones hifales y evaluar el efecto de los metabolitos volátiles contra el crecimiento micelial de aislados de S. rolfsii obtenidos de suelos sometidos a la siembra de papa en los estados de Sonora y Sinaloa, México.
Materiales y Métodos
Obtención de aislados. En el estudio, se utilizaron 16 aislados de T. asperellum (TAM), cinco de T. asperelloides (TES), cuatro de T. afroharzianum (TAF), uno de T. azevedoi (TAI) y dos de S. rolfsii (Scr4 y Scr17). Dichos aislados fueron proporcionados por el banco microbiológico del Laboratorio de Diagnóstico Fitosanitario de la Junta Local de Sanidad Vegetal del Valle del Fuerte. Los aislados se obtuvieron de los municipios Altar y Caborca, pertenecientes al estado de Sonora, y Ahome, Guasave, El Fuerte y Sinaloa de Leyva, del estado de Sinaloa, durante los ciclos agrícolas del 2019-2021. Las secuencias de los aislados se depositaron en el GenBank, con los códigos OR521159- OR521184 para Trichoderma y OR514113 y OR514124 para Sclerotium rolfsii (Cuadro 1).
Efecto inhibitorio in vitro de 26 aislados de Trichoderma spp. contra Sclerotium rolfsii. El antagonismo in vitro de 26 aislados de cuatro especies de Trichoderma con tres días de crecimiento se determinó contra dos aislados de S. rolfsii (Scr4 y Scr17) de siete días de crecimiento en medio de cultivo Papa Dextrosa Agar (PDA) (BD Bioxon, Becton, Dickinson de México, S.A. de C.V.). La evaluación se llevó a cabo, mediante la técnica de cultivo dual (Dennis y Webster, 1971). Discos miceliales de 5 mm de diámetro fueron obtenidos tanto de las colonias puras de los aislados antagónicos como del hongo. Los discos de S. rolfsii, se colocaron en la periferia de la caja Petri de 90 mm de diámetro y de Trichoderma spp. en el extremo opuesto. Los tratamientos se distribuyeron en un diseño completamente al azar con cuatro réplicas (cuatro cajas Petri) por cada combinación de S. rolfsii-Trichoderma. El experimento se desarrolló en dos ocasiones.
Cajas Petri con PDA que contenían cultivos individuales tanto de S. rolfsii como de Trichoderma sirvieron como control (Pacheco et al., 2016). Las cajas Petri selladas con Parafilm, se incubaron a 25 ± 1 °C en oscuridad constante en una cámara bioclimática Thermo Scientific. Para determinar la capacidad antagónica de Trichoderma spp., se midió el crecimiento micelial, cada 24 h después de la siembra y terminó cuando S. rolfsii llenó las cajas Petri del grupo control. El porcentaje de inhibición del crecimiento micelial del patógeno de prueba (I) se calculó con la siguiente ecuación: Porcentaje de inhibición: I =C-T/C × 100, donde: C= Crecimiento del patógeno control y T= Crecimiento del patógeno en tratamiento (Kotasthane et al., 2014). La efectividad biológica de las especies de Trichoderma en la inhibición de Sclerotium rolfsii se determinó mediante la escala de clases del 1 al 5 de Bell et al. (1982) donde: clase 1) Trichoderma cubrió por completo la superficie del medio de cultivo, clase 2) Trichoderma creció al menos el 65 % del medio, clase 3) Trichoderma y S. rolfsii crecieron en un 50 % del medio, clase 4) S. rolfsii creció al menos el 65 % del medio y clase 5) S. rolfsii creció completo sobre la superficie del medio de cultivo.
Cuadro 1 Origen e identificación de aislados de Sclerotium rolfsii y especies de Trichoderma.
| Especie/aislado | Año de muestreo | Localidad | Código en Gen Bank |
|---|---|---|---|
| Sclerotium rolfsii /Scr4 | 2019 | Ahome, Sinaloa | OR514113 |
| Sclerotium rolfsii /Scr17 | 2019 | Ahome, Sinaloa | OR514124 |
| T. asperelloides /TES19 | 2020 | Caborca, Sonora | OR521159 |
| T. asperelloides /TES20 | 2020 | Altar, Sonora | OR521160 |
| T. afroharzianum /TAF21 | 2020 | Altar, Sonora | OR521161 |
| T. asperellum /TAM22 | 2020 | Caborca, Sonora | OR521162 |
| T. asperelloides /TES23 | 2020 | Caborca, Sonora | OR521163 |
| T. asperelloides /TES24 | 2020 | Caborca, Sonora | OR521164 |
| T. asperelloides /TES26 | 2020 | Caborca, Sonora | OR521165 |
| T. asperellum/TAM27 | 2019 | Guasave, Sinaloa | OR521166 |
| T. asperellum/TAM30 | 2019 | El Fuerte, Sinaloa | OR521167 |
| T. asperellum/TAM31 | 2019 | Guasave, Sinaloa | OR521168 |
| T. afroharzianum /TAF33 | 2020 | Sinaloa de Leyva, Sinaloa | OR521169 |
| T. asperellum/TAM35 | 2020 | Ahome, Sinaloa | OR521170 |
| T. asperellum/TAM37 | 2020 | Guasave, Sinaloa | OR521171 |
| T. afroharzianum/TAF38 | 2020 | Guasave, Sinaloa | OR521172 |
| T. asperellum/TAM57 | 2021 | Ahome, Sinaloa | OR521173 |
| T. asperellum/TAM59 | 2021 | Guasave, Sinaloa | OR521174 |
| T. asperellum/TAM64 | 2021 | Ahome, Sinaloa | OR521175 |
| T. asperelloides/TES65 | 2021 | Ahome, Sinaloa | OR521176 |
| T. asperellum/TAM67 | 2021 | Ahome, Sinaloa | OR521177 |
| T. asperellum/TAM68 | 2021 | Ahome, Sinaloa | OR521178 |
| T. asperellum/TAM69 | 2021 | Ahome, Sinaloa | OR521179 |
| T. asperellum/TAM70 | 2021 | Ahome, Sinaloa | OR521180 |
| T. azevedoi /TAI73 | 2021 | Ahome, Sinaloa | OR521181 |
| T. asperellum/TAM74 | 2021 | Ahome, Sinaloa | OR521182 |
| T. asperellum/TAF75 | 2021 | Ahome, Sinaloa | OR521183 |
| T. asperellum/TAM76 | 2020 | Caborca, Sonora | OR521184 |
Interacción hifal de Trichoderma spp. contra S. rolfsii. La interacción hifal de 26 aislados de Trichoderma spp. contra S. rolfsii (Scr4) se evaluó mediante la metodología de cultivos duales (Dennis y Webster, 1971). La observación de la interacción se llevó a cabo al momento del contacto entre los hongos, de los cuales se tomaron tres muestras de la zona de interacción. Para esto, se colocó un fragmento de cinta adhesiva transparente sobre las hifas del hongo y de Trichoderma spp., enseguida la cinta se colocó sobre una gota de lactofenol-azul de algodón en un portaobjetos y se observó al microscopio óptico para identificar vacuolización, granulación, enrollamiento, adhesión, lisis o penetración causados por los aislados de Trichoderma spp. (Martínez et al., 2018).
Efecto de metabolitos volátiles de especies de Trichoderma con actividad antifúngica contra S. rolfsii. Para determinar el efecto inhibitorio de metabolitos volátiles de los 26 aislados de Trichoderma spp. contra un aislado de S. rolfsii (Scr4), éstos se sembraron en PDA y se incubaron a 25 ± 2 °C por tres y siete días, respectivamente. Posteriormente, discos del mismo medio de 5 mm de diámetro con micelio activo de S. rolfsii y 26 aislados de Trichoderma spp. se transfirieron al centro de cajas Petri con PDA. El inóculo de las especies de Trichoderma, se transfirió 24 h antes que el de S. rolfsii. Para confrontar los aislados, las bases de las cajas Petri se ensamblaron y se sellaron con Parafilm (Rajani et al., 2021). En la parte inferior se ubicó a Trichoderma y en la parte superior S. rolfsii (Dennis y Webster, 1971; Mokhtar y Dehimat, 2012). Los tratamientos se distribuyeron en un diseño completamente al azar con cuatro réplicas por cada combinación de S. rolfsii-Trichoderma. El experimento se desarrolló en dos ocasiones. Como control se incluyeron cajas Petri con PDA con cultivos individuales de S. rolfsii como de Trichoderma, las cuales se incubaron a 25 ± 2 °C. El efecto de inhibición de Trichoderma spp. se determinó una vez que S. rolfsii cubrió el medio de cultivo en el tratamiento control sin Trichoderma para lo cual se consideró el porcentaje de inhibición del crecimiento de S. rolfsii mediante la fórmula descrita por Kotasthane et al. (2014), donde: Porcentaje de inhibición: I =C-T/C × 100, donde: C= Crecimiento del patógeno testigo, T= Crecimiento del patógeno en tratamiento.
Análisis estadísticos de datos. Los datos sobre inhibición micelial in vitro mediante confrontaciones en cultivos duales de los 26 aislados de Trichoderma contra dos aislados de S. rolfsii; de igual forma, los datos de inhibición del crecimiento micelial de un aislados de S. rolfsii por los metabolitos volátiles de 26 aislados de Trichoderma spp., se analizaron mediante estadística no paramétrica (Kruskal-Wallis). La separación de medias de inhibición del crecimiento micelial se realizó mediante los procedimientos de Conover (1999) con (P ≤ 0.05).
Resultados
Inhibición in vitro de aislados de Trichoderma spp. sobre S. rolfsii. Los resultados mostraron diferencias significativas (P ≤ 0.05) en el efecto inhibitorio in vitro de S. rolfsii entre los 26 aislados de Trichoderma evaluados, ya que inhibieron los dos aislados de S. rolfsii (Scr4 y Scr17), con porcentaje de inhibición de 21.0 a 75.4 % en Scr4 y de 23.6 a 77.1 % en Scr17.
El aislado TAI73 de T. azevedoi mostró el mayor efecto inhibitorio in vitro en el crecimiento micelial del aislado Scr4 de S. rolfsii (Figura 1A) con porcentaje de inhibición de 75.4 %, seguido de los aislados TAM74 (68.9 %), TAM30 (66.6 %), TAF75 (66.0 %), TES65 (65.0 %), TES24 (64.9 %), TAM68 (64.9 %) y TAM37 (64.8 %). Todos los niveles de inhibición se ubicaron en la clase 2, sin diferencias significativas entre éstos (P ≥ 0.05) pero sí con el resto de los aislados de Trichoderma. Mientras que el aislado TAF33 mostró el menor porcentaje de inhibición (21.0 %) (Figura 1B) y correspondió a la clase 4, seguido de TAM70 (56.7 %), TAF21 (57.2 %), TAM76 (57.3 %), TAM57 (57.3 %) y TAM31 (59.0 %), ubicándose todas en la clase 3, sin diferencias significativas entre tratamientos (P ≥ 0.05) (Cuadro 2).
Por otro lado, el aislado de Trichoderma TAI73 mostró el mayor nivel de inhibición en el crecimiento micelial de Scr17 con 77.1 % (clase 2), seguido de TAM69 (68.8 %), TES24 (67.9 %) (Figura 1C), TAM30 (67.9 %), TES65 (67.3 %), TAF75 (67.0 %),
TAM74 (66.4 %), TAM35 (66.2 %) y TAM37 (66.1 %) en clase 2, sin diferencias significativas entre ellos (P ≥ 0.05) (Cuadro 2). En cambio, el aislado de Trichoderma TAF33 (Figura 1D) (clase 4) con 23.6 % mostró el menor porcentaje de inhibición contra Scr17, seguido de TAF38 (52.8 %), TAM31 (57.4 %), TAM68 (59.8 %), TAM59
(60.1 %), TAM64 (60.3 %) y TES20 (60.3 %) todas en clase 3, sin diferencias significativas entre ellos (P ≥ 0.05) (Cuadro 2).
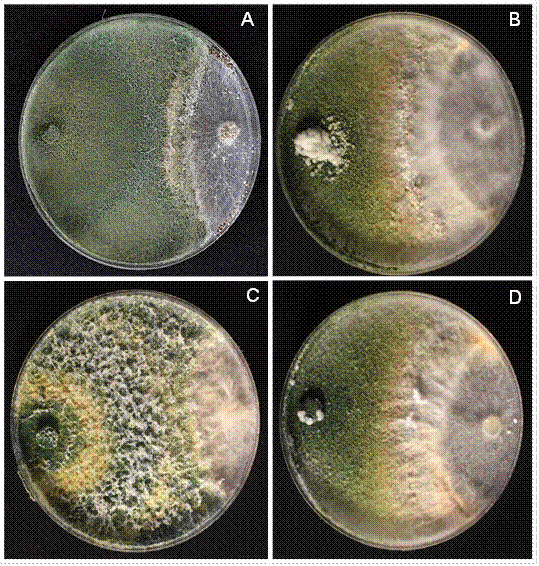
Figura 1 Antagonismo in vitro de Trichoderma spp. (izquierda) contra S. rolfsii (derecha). (A) Aislado de Trichoderma TAI73 contra S. rolfsii Scr4, (B) Trichoderma TAF33 contra S. rolfsii Scr4, (C) Trichoderma TES24 contra S. rolfsii Scr17 y (D) Trichoderma TAF33 contra S. rolfsii Scr17.
El aislado de T. azevedoi (TAI73) presentó el mayor porcentaje de inhibición del crecimiento micelial in vitro del aislado Scr4 de S. rolfsii, con 75.4 % de inhibición (Cuadro 2); mientras que el mismo aislado de Trichoderma ejerció una inhibición de
77.1 % en el aislado Scr17 de S. rolfsii. En contraste, T. afroharzianum (TAF33) tuvo el menor efecto inhibitorio sobre los dos aislados de S. rolfsii (Cuadro 2). Se observó una marcada variación en el efecto de los aislados de Trichoderma sobre la inhibición de S. rolfsii con diferencias significativas entre ellos (P ≤ 0.05) (Cuadro 2).
Cuadro 2 Antagonismo in vitro de 26 aislados de Trichoderma spp. contra dos aislados de Sclerotium rolfsii.
| Aislado Scr4 de S. rolfsii | Aislado Scr17 de S. rolfsii | ||||
|---|---|---|---|---|---|
| Aislado de Trichoderma | % de inhibición | Escala | Aislado de Trichoderma | % de inhibición | Escala |
| Testigo | 0.0 az | 5 | Testigo | 0.0 a* | 5 |
| TAF33 | 21.0 ab | 4 | TAF33 | 23.6 ab | 4 |
| TAM70 | 56.7 abc | 3 | TAF38 | 52.8 ab | 3 |
| TAF21 | 57.2 abcd | 3 | TAM31 | 57.4 abc | 3 |
| TAM76 | 57.3 abcd | 3 | TAM68 | 59.8 abc | 3 |
| TAM57 | 57.3 abcd | 3 | TAM59 | 60.1 abcd | 3 |
| TAM31 | 59.0 abcde | 3 | TAM64 | 60.3 abcd | 3 |
| TES26 | 60.3 bcdef | 2 | TES20 | 60.3 abcde | 3 |
| TAM67 | 60.5 cdefg | 2 | TAF21 | 60.4 bcde | 2 |
| TAF38 | 60.5 cdefg | 2 | TES26 | 60.5 bcde | 2 |
| TAM64 | 60.5 cdefg | 2 | TAM57 | 61.9 cdef | 2 |
| TAM59 | 60.9 defgh | 2 | TAM23 | 61.9 cdef | 2 |
| TAM23 | 61.9 efghi | 2 | TAM70 | 62.6 defg | 2 |
| TAM27 | 62.0 efghi | 2 | TAM67 | 63.0 defg | 2 |
| TAM22 | 62.7 fghij | 2 | TAM22 | 63.4 efgh | 2 |
| TAM69 | 63.2 fghij | 2 | TAM27 | 63.4 efgh | 2 |
| TES20 | 63.3 fghij | 2 | TAM76 | 64.7 fghi | 2 |
| TES19 | 63.4 ghijk | 2 | TES19 | 65.0 fghi | 2 |
| TAM35 | 63.6 hijk | 2 | TAM37 | 66.1 ghij | 2 |
| TAM37 | 64.8 ijkl | 2 | TAM35 | 66.2 ghij | 2 |
| TAM68 | 64.9 ijkl | 2 | TAM74 | 66.4 ghij | 2 |
| TES24 | 64.9 ijkl | 2 | TAF75 | 67.0 hij | 2 |
| TES65 | 65.0 jkl | 2 | TES65 | 67.3 hij | 2 |
| TAF75 | 66.0 jkl | 2 | TAM30 | 67.9 ij | 2 |
| TAM30 | 66.6 kl | 2 | TES24 | 67.9 ij | 2 |
| TAM74 | 68.9 l | 2 | TAM69 | 68.8 j | 2 |
| TAI73 | 75.4 l | 2 | TAI73 | 77.1 j | 2 |
zMedias con una letra común no son significativamente diferentes (P ≤ 0.05), Kruskal-Wallis.
Determinación de interacciones hifales de Trichoderma spp. y S. rolfsii. La zona de encuentro hifal entre aislados de Trichoderma spp. y S. rolfsii (Scr4) mostró varios tipos de interacción; donde 23 aislados del antagonista causaron lisis, (Figura 2A), 20 produjeron granulación (Figura 2B), 20 mostraron adhesión (Figura 2C), 19 presentaron enrollamiento (Figura 2D), 13 indujeron vacuolización (Figura 2E) y uno mostró penetración (Figura 2F). El aislado TES65 de T. asperelloides fue el único que penetró las hifas de S. rolfsii (Cuadro 3, Figura 2F). Los aislados de Trichoderma spp. TAM37, TAM59, TES65, TAI73 y TAM76 mostraron hasta cinco interacciones al confrontarlo con S. rolfsii. Mientras que los aislados TES20, TAF21, TAM23, TES24, TES26, TAM30, TAM31, TAF33, TAM35, TAM57, TAM68, TAM70 y TAF75 del hongo antagónico mostraron tres interacciones (Cuadro 3).
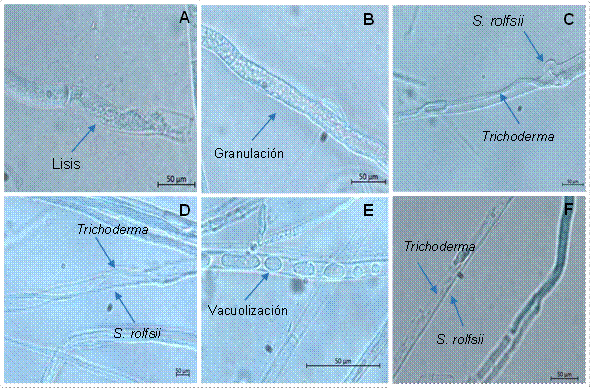
Figura 2 Interacciones hifales entre aislados de Trichoderma spp. y Sclerotium rolfsii. (A) Lisis (TAM67), (B) granulación (TES65), (C) adhesión (TAM69), (D) enrollamiento (TAM59), (E) vacuolización (TAM76) y (F) penetración (TES65).
Cuadro 3 Interacción hifal in vitro de 26 aislados de Trichoderma spp. y Sclerotium rolfsii (Scr4).
| Aislado de Trichoderma | Tipos de interacciones hifales | |||||
|---|---|---|---|---|---|---|
| Lisis | Granulación | Adhesión | Enrollamiento | Vacuolización | Penetración | |
| TES19 | x | x | x | x | ||
| TES20 | x | x | x | |||
| TAF21 | x | x | x | |||
| TAM22 | x | x | x | x | ||
| TAM23 | x | x | x | |||
| TES24 | x | x | x | |||
| TES26 | x | x | x | |||
| TAM27 | x | x | x | x | ||
| TAM30 | x | x | x | |||
| TAM31 | x | x | x | |||
| TAF33 | x | x | x | |||
| TAM35 | x | x | x | |||
| TAM37 | x | x | x | x | x | |
| TAF38 | x | x | x | x | ||
| TAM57 | x | x | x | |||
| TAM59 | x | x | x | x | x | |
| TAM64 | x | x | x | x | ||
| TES65 | x | x | x | x | x | |
| TAM67 | x | x | x | x | ||
| TAM68 | x | x | x | |||
| TAM69 | x | x | x | x | ||
| TAM70 | x | x | x | |||
| TAI73 | x | x | x | x | x | |
| TAM74 | x | x | x | x | ||
| TAF75 | x | x | x | |||
| TAM76 | x | x | x | x | x | |
Nota: El montaje de la zona de interacción se tiñó con lactofenol-azul cuando las hifas de los hongos interactuaron.
Inhibición in vitro de metabolitos volátiles de Trichoderma contra S. rolfsii. Los metabolitos volátiles de 26 aislados de Trichoderma spp. inhibieron el crecimiento micelial de S. rolfsii (Scr4) in vitro. El porcentaje de inhibición varió de 26 a 81.4 %. El mayor porcentaje de inhibición lo ejerció el aislado TAI73 de T. azevedoi con un 81.4 %, mientras que los porcentajes de inhibición de los aislados TAM68 (Figura 3A), TES65, TAM74, TAM57, TAF75, TAM70 y TAM37 variaron de 78.2 a 70.9 %, sin diferencias significativas entre ellos (P ≥ 0.05) (Cuadro 4). Los aislados TAM59, TAM35, TES26, TES19, TAF38, TAM27 y TAM30 (Figura 3B), ejercieron una inhibición de 49.6 a 57.1
% contra S. rolfsii. El aislado TAF33 de T. afroharzianum ejerció el menor porcentaje de inhibición equivalente a 26 %. La variación del efecto inhibitorio de los aislados de Trichoderma se reflejó con diferencias significativas entre aislados (P ≤ 0.05) (Cuadro 4).
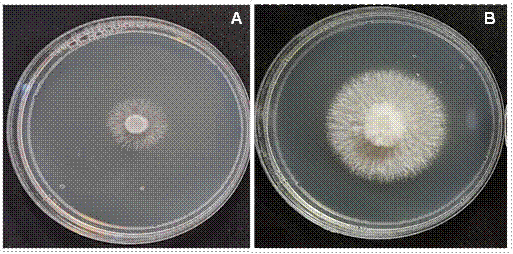
Figura 3 Efecto inhibitorio de metabolitos volátiles de Trichoderma spp. contra Sclerotium rolfsii. (A) Aislado de Sclerotium rolfsii (Scr4) expuesto a metabolitos volátiles de Trichoderma TAM68 y (B) S. rolfsii (Scr4) expuesto a metabolitos volátiles de Trichoderma TAM30.
Cuadro 4 Efecto de metabolitos volátiles de aislados de Trichoderma contra Sclerotium rolfsii Scr4.
| Aislado de Trichoderma | % de inhibición | Aislado de Trichoderma | % de inhibición |
|---|---|---|---|
| Testigo | 00.0 az | TAM31 | 62.5 efghij |
| TAF33 | 26.0 a | TAM64 | 63.0 fghijk |
| TAM59 | 49.6 ab | TAM76 | 65.8 ghijk |
| TAM35 | 50.3 ab | TAM67 | 68.1 hijkl |
| TES26 | 54.1 abc | TAF21 | 69.2 ijklm |
| TES19 | 55.6 abcd | TAM37 | 70.9 ijklmn |
| TAF38 | 56.3 abcde | TAM70 | 72.0 jklmn |
| TAM27 | 56.9 abcdef | TAF75 | 73.5 klmn |
| TAM30 | 57.1 abcdef | TAM57 | 73.5 klmn |
| TES20 | 58.4 bcdefg | TAM74 | 76.3 lmn |
| TES24 | 58.7 bcdefg | TES65 | 76.3 lmn |
| TAM22 | 59.5 cdefgh | TAM68 | 78.2 mn |
| TAM69 | 61.4 defghi | TAI73 | 81.4 n |
| TAM23 | 62.2 efghij |
zMedias con una letra común no son significativamente diferentes (P ≤ 0.05), Kruskal-Wallis.
Discusión
El uso de Trichoderma como agente de control biológico en el cultivo de papa es limitado, pues los productores prefieren el uso de fungicidas sintéticos, argumentando que la eficacia de este tipo de sustancia es conocida. En México, no hay evidencia científica sobre el uso de diferentes especies de Trichoderma para controlar la pudrición blanda de la papa.
En este estudio, se encontró que los aislados de T. azevedoi, T. asperellum, T. asperelloides y T. afroharzianum provenientes de suelos sometidos al cultivo de papa, mostraron diferentes niveles de inhibición sobre el crecimiento micelial de S. rolfsii in vitro. T. azevedoi presentó el mayor efecto inhibitorio, con un 75.4 % contra S. rolfsii (Scr4) y un 77.1 % contra la misma especie (Scr17). Estos resultados coinciden con los reportados por Kamel et al. (2020), quienes evaluaron seis especies de Trichoderma contra tres aislados de S. rolfsii provenientes de cultivos de frijol, tomate y repollo en Egipto. En este estudio, T. viride mostró el mayor efecto inhibitorio (85.6 %) contra S. rolfsii obtenido de tomate, mientras T. harzianum inhibió en 79.3 y 77.8 % al mismo patógeno aislado de repollo y frijol, respectivamente. De manera similar Vineela et al. (2020) reportaron que T. hamatum inhibió el crecimiento micelial de S. rolfsii en un 75.2%, seguido de T. harzianum (74.7 %) y T. viride (71.9 %), obtenidos de cultivos de cacahuate (Arachis hypogaea) en la India. Por otro lado, Safari et al. (2022) evaluaron 30 hongos antagonistas mediante cultivo dual y reportaron que T. virens mostró la mayor capacidad de supresión del crecimiento micelial de S. rolfsii (90.8 %) aislado de cultivos de cacahuate en Irán, seguido de T. viride con un 66.8 %. De manera similar Kushwaha et al. (2018) determinaron que T. harzianum ejerció un efecto inhibitorio de 63.5 % in vitro, seguido de T. virens con un 51.5 % y T. viride con 50.8 % contra S. rolfsii aislado de cultivos de lenteja (Lens culinaris) en la India. En contraste Chandra-Sekhar et al. (2020) reportaron una baja inhibición in vitro del crecimiento micelial de S. rolfsii obtenido de tomate en la India, donde T. harzianum inhibió un 50 %, T. viride 39.7 % y T. asperellum 33.8 %.
En el presente estudio, se encontró que, de los 26 aislados de Trichoderma evaluados contra S. rolfsii (Scr4), 20 de ellos (73.0 %) se ubicaron en la clase 2 de acuerdo a la escala de Bell (Bell et al., 1982); mientras que 17 aislados del antagonista (73.0 %) se clasificaron en la clase 2 en cuanto a la inhibición del crecimiento micelial de S. rolfsii (Scr17). Estos resultados sugieren que su potencial de control debería de evaluarse en campo. Por otro lado, Kotasthane et al. (2014) evaluaron 20 aislados de T. harzianum, T. aureoviride, T. viride y T. virens procedentes de cultivos de arroz (Oryza sativa) en la India, de los cuales 19 se ubicaron en la clase 1 y 2, mientras que solo uno se clasificó en clase 3. Estos resultados contrastan con los obtenidos por Bell et al. (1982), quienes evaluaron 77 aislados de Trichoderma. En su estudio, el antagonismo contra S. rolfsii no fue evidente, ya que 88.0 % de los aislados se clasificaron en las clases 3, 4 y 5 con acción limitada, mientras que sólo el 1.0 % se ubicó en la clase 2. Es importante recalcar que la competencia por espacio y nutrientes en las especies de Trichoderma en procesos antagónicos contra fitopatógenos es una característica deseable, ya que está asociada a su crecimiento rápido y producción de metabolitos antimicrobianos (Chandra et al., 2020). Por ello, es necesario realizar estudios adicionales para identificar los metabolitos secundarios, especialmente en T. azevedoi, que mostró el mayor antagonismo contra los dos aislados de S. rolfsii en este trabajo.
Con relación a la interacción hifal, los aislados de Trichoderma incluidos en el presente estudio provocaron vacuolización, granulación, enrollamiento, adhesión, lisis y penetración de las hifas de S. rolfsii, lo que causó la desintegración del micelio del hongo, resaltando T. azevedoi, que presentó el mayor número de interacciones hifales. Resultados similares fueron reportados por Kamel et al. (2020), quienes observaron que la mayoría de las especies de Trichoderma se adhirieron, enrollaron y provocaron la lisis de las hifas del patógeno en la India. En nuestro estudio destaca la acción de T. azevedoi, donde en las diferentes interacciones pueden atribuirse la capacidad de producción de enzimas hidrolíticas como la glucanasa y quitinasa, los cuales afectan la integridad celular de las hifas, provocando el colapso del micelio de S. rolfsii (Kotasthane et al., 2014; Kamel et al., 2020).
Los resultados del presente estudio indican variabilidad en la inhibición del crecimiento micelial de S. rolfsii (26.0 a 81.4 %), resaltando T. azevedoi en su capacidad inhibitoria; esto coincide con lo reportado por Sangle et al. (2016), quienes evaluaron 30 aislados de T. harzianum T. viride, T. hamatum, T. reesei, T. piluliferum y T. virens contra S. rolfsii proveniente de cultivo de lenteja en la india, donde T. harzianum presentó el mayor efecto inhibitorio (84.5 %) del crecimiento micelial del patógeno. De igual manera, Amin et al. (2010), evaluaron T. harzianum, T. virens y T. viride contra S. rolfsii procedentes de diferentes cultivos en la India, obteniendo porcentajes de inhibición menores a los reportados en el presente estudio (20 al 40.0 %). Por otro lado, Ruangwong et al. (2021) analizaron el efecto inhibitorio de T. asperelloides, proveniente de flores de flamenco (Anthurium sp.) en Tailandia, contra S. rolfsii, el cual inhibió el crecimiento micelial del 41.2 %. En su estudio identificaron 17 tipos de metabolitos volátiles, entre los cuales destacó la presencia del 2-feniletanol, un compuesto con propiedades antifúngicas. Sridharan et al. (2020) encontraron que T. longibrachiatum, procedente del cultivo de arroz en la India, inhibió el crecimiento de S. rolfsii y Macrophomina phaseolina en 57 a 35.0 %, respectivamente. Además, este antagonista causó alteraciones en la estructura micelial, pues redujo el grosor del micelio y disminuyó el número de esclerocios en comparación con el patógeno en ausencia del antagonista. Estos resultados, evidencian la capacidad de los metabolitos volátiles producidos por aislados de Trichoderma para inhibir el crecimiento micelial de S. rolfsii sin necesidad de contacto directo entre estos hongos; esta característica reviste importancia en la protección de las plantas de papa contra S. rolfsii (Sangle et al., 2016). Los resultados del presente estudio abren nuevas líneas de investigación relacionadas con la identificación de los mecanismos moleculares involucrado en la inhibición del crecimiento micelial, así como para evaluar la estabilidad de estos efectos en condiciones de campo. Los resultados del presente estudio son relevantes, ya que la mayoría de los aislados de Trichoderma mostraron potencial antagónico, destacando T. azevedoi como potencial agente de control biológico contra S. rolfsii en el cultivo de papa.
Conclusiones
El aislado TAI73 de T. azevedoi presentó el mejor potencial antagónico para el control de S. rolfsii en todas las pruebas in vitro realizadas en el presente estudio. T. azevedoi inhibió en un 75.4 y 77.1 % el crecimiento micelial de los aislados Scr4 y Scr17 de S. rolfsii, respectivamente y se ubicó en la clase 2 contra ambos aislados en la escala de Bell, lo que muestra su alta capacidad antagónica. Este antagonista también causó vacuolización, granulación, enrollamiento, adhesión y lisis de las hifas de S. rolfsii (Scr4). Además, T. azevedoi produjo metabolitos volátiles, los cuales inhibieron en 84.1 % el crecimiento micelial in vitro del hongo. Estos resultados abren nuevas líneas de investigación relativas a la determinación de la efectividad biológica de T. azevedoi como potencial agente de biocontrol en campo.

