











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Las plantas ornamentales son importantes en los diferentes nichos ecológicos. Se caracterizan por su belleza, color y aroma. A través de distintas investigaciones se ha reportado que además de su valor estético, poseen una influencia sobre la microbiología y las características del suelo (Francini et al., 2022). Tal es el caso de Euphorbia mili y Calendula officinalis, utilizadas en la biorremediación del suelo, al disminuir metales pesados como el Cromo, Cadmio y Plomo (Afrousheh et al., 2015), lo que, a su vez, influye sobre los microorganismos presentes en el suelo al provocar efectos positivos o negativos en su supervivencia. Además, se ha comprobado que este tipo de plantas favorecen la absorción de Nitrógeno y Potasio al mejorar sus formas asimilables e incrementan el contenido de materia orgánica (Sari y Küçük, 2019). Así mismo, diversos estudios han confirmado que el efecto de las plantas ornamentales en las propiedades físico-químicas del suelo, están asociadas a la diversidad de la microbiota edáfica (Zhang et al., 2023), donde destaca la comunidad fúngica, la cual juega un papel clave en la regulación de la relación diversidad-productividad de las plantas (Mommer et al., 2018), al ser uno de los microorganismos fundamentales en la descomposición de materia orgánica favoreciendo la estructura, fertilidad y salud del suelo (Frac et al., 2018). Al mismo tiempo, se ha establecido que la diversidad de estos organismos puede ser afectada por algunas especies vegetales, las cuales liberan metabolitos secundarios con acción antimicrobiana como las saponinas, la cuales alteran negativamente la membrana celular de los hongos, aumentan los niveles de Ca2+ y especies reactivas de oxígeno (Turner et al., 2013). Además, se ha comprobado que las cumarinas, triterpenos, flavonoides, benzoxazinoides y fitohormonas, inhiben el crecimiento de los microorganismos rizosféricos (Voges et al., 2019). Debido a la importancia de la relación planta- microbioma, actualmente existen numerosas investigaciones al respecto; sin embargo, son escasos los estudios relacionados al efecto de las plantas ornamentales sobre la comunidad fúngica del suelo, así como su correlación con las características edáficas. Por lo anterior el objetivo del presente estudio fue evaluar la respuesta biológica de plantas ornamentales en la diversidad y riqueza del microbioma fúngico rizosférico y las propiedades del suelo en Baja California Sur.
Materiales y Métodos
Área de estudio. La investigación se llevó a cabo en el ciclo 2022-2023 en el municipio de La Paz, Baja California Sur, México (24°7′0″ N, 110°43′0″ W y altitud de 0 a 27 msnm). La región se caracteriza por una precipitación total anual promedio menor a 200 mm y una temperatura media anual de 18 a 22 °C (INEGI, 2021). El clima es desértico cálido (BWh) y semiárido cálido (BSh) según la clasificación de Köppen. El suelo es considerado franco-arenoso. La vegetación predominante son los matorrales.
Obtención de plantas ornamentales. Seis especies de plantas ornamentales fueron seleccionadas para la presente investigación considerando características tales como; demanda comercial, baja necesidad hídrica, requerimientos de luminosidad intensos, ciclo biológico corto y ser una especie introducida al estado. Dos ornamentales fueron adquiridas a partir de esquejes: Corona de cristo (Euphorbia mili: Malpighiales- Euphorbiaceae) y Petunia mexicana (Ruellia simplex: Lamiales-Acanthaceae). Mientras que las cuatro ornamentales restantes se adquirieron mediante semillas de lotes comerciales (Vita®): Teresita (Catharanthus roseus: Gentianales-Apocynaceae), Pincel (Centaurea cyanus: Asterales-Asteraceae), Cempasúchil (Tagetes erecta: Asterales- Asteraceae) y Caléndula (Calendula officinalis: Asterales-Asteraceae).
Diseño experimental. El trabajo experimental se llevó a cabo a nivel de campo agrícola (con más de 10 años sin siembra de hortalizas), mediante el establecimiento de siete parcelas, con una superficie de 81 m2, constituida cada una por tres surcos de 15 m de largo x 80 cm de ancho. Las semillas del segundo grupo de plantas ornamentales fueron sembradas directamente en los surcos a una profundidad al doble de su tamaño (1-2 cm) con un marco de plantación de 30 cm. Mientras que los esquejes de 15 cm de largo, se trasplantaron a una profundidad de 10 cm, con un marco de plantación de 0.75 cm y 1.5
m. Los tratamientos consistieron en seis especies ornamentales y un control (sin cultivo). Cada tratamiento contó con tres repeticiones donde una repetición correspondía a un surco. Todos los tratamientos fueron arreglados bajo un diseño en bloques completos al azar.
Muestreo, aislamiento, cuantificación e identificación de aislados fúngicos. Nueve muestras de suelo rizosférico fueron obtenidas al azar de cada unidad experimental a 30 cm de profundidad, antes, durante y al final del experimento (n=27 muestras/tratamiento) por un periodo de ocho meses (Septiembre-abril). El aislamiento de hongos, fue mediante diluciones seriadas (103) y siembra en Papa-Dextrosa-Agar (PDA) (MCD LAB, Oaxaca, México) a pH 7. 10 días después de la siembra (dds) se determinó la población en UFC g -1 de suelo. A partir de aislados puros, se realizaron preparaciones semipermanentes en portaobjetos y se visualizaron bajo microscopio óptico (LABOMED-40x) para la identificación morfológica fúngica, la cual se llevó a cabo mediante la caracterización macroscópica (morfología y color de colonias) y microscópica (forma de micelio, esporas y cuerpo fructífero) y se compararon con claves taxonómicas (Barnett y Hunter 1972; Kirk et al., 2008; Samson et al., 2014).
Diversidad y riqueza de especies. La diversidad fúngica en cada una de las plantas ornamentales se determinó a través de la frecuencia de ocurrencia de cada especie, calculada al dividir el número de colonias de una especie dada entre el número total de comunidades aisladas en cada muestra, expresada en porcentaje. Mientras que la Riqueza (S = número total de especies) fue determinada mediante la fórmula propuesta por Kumari et al. (2016) a través del índice de la diversidad de Shannon (H´= -Σ pi Ln pi, donde pi es la proporción de individuos de las especies i) y Simpson (D = Σ(n/N) 2, donde n es el número total de organismos de una especie en particular y N es el número total de organismos de todas las especies).
Evaluación de las características edafológicas. El análisis edafológico del suelo se llevó a cabo antes del establecimiento de las plantas y al final del experimento por un período de ocho meses. El efecto de las plantas ornamentales sobre las propiedades del suelo fue evaluado mediante la determinación de pH, Materia orgánica (MO.) Fósforo (P), Potasio (K), Calcio (Ca), Magnesio (Mg) y Conductividad Eléctrica (CE). El pH del suelo se obtuvo a través de un potenciómetro (VWR scientific products modelo sp20). Mientras que la medición de Materia orgánica fue con el método de descomposición húmeda Walkley-Black actualizado (Walkley, 1942) Así mismo, la determinación de Fósforo se obtuvo a través del método OLSEN. Potasio por el método PECH, así como Calcio y Magnesio por el método EDTA. Todas las mediciones de macro y microelementos se realizaron en el espectrofotómetro (Marca spectronic instruments, modelo spectronic 20 GENESYS) y la conductividad electica se determinó por el método puente de wheatsone con ayuda de un conductímetro (Bridge model-31).
Análisis estadístico. Los datos se organizaron en Microsoft Excel 2019 para su procesamiento. Para identificar las diferencias significativas, en la diversidad, índices de Simpson y Shannon, así como las propiedades químicas del suelo, se uso un análisis de varianza de una vía y un rango de confianza de 0.05 con Tukey con el programa estadístico Prism. Así mismo, se analizó la comparación de los coeficientes de correlación de Pearson entre la diversidad de hongos y las propiedades del suelo en las plantas ornamentales.
Resultados y Discusión
Población. Las poblaciones fúngicas en las especies ornamentales presentaron variaciones a través del tiempo, donde en la mayoría de los casos la población disminuyó en el mes de octubre, lo cual se relacionó con los cambios en la temperatura y la permanencia de humedad en el suelo. C. cyanus aumentó la población fúngica al pasar de 2,833.33 a 8,096.66 UFC g -1 de suelo (Figura 1). Mientras que en E. mili el aumento de la población fue menor (de 3,159 a 3,559 UFC g -1 de suelo). Por su parte la especie C. roseus presentó una población de 3,126.66 UFC g -1 de suelo y al final su población incrementó a 5,086.66 UFC g -1. Por otro lado, C. officinalis inició con 1,060 UFC g -1 de suelo y aumentó considerablemente a 6,633.33 UFC g -1 de suelo al finalizar el experimento. De la misma manera R. simplex mantuvo una población inicial de 1,686.66 UFC g -1 de suelo y finalizó con 5,720 UFC g -1. En T. erecta su población fue de 2, 323.33 a 3,333.33 UFC g -1 de suelo al inicio y final respectivamente. Por último, en el tratamiento control se presentó una variación significativa en la población fúngica, al iniciar con 720 UFC g -1 de suelo y finalizó con 12,483.33 UFC g -1 de suelo, superando la población en las plantas ornamentales (Figura 1). En general, la mayoría de las plantas ornamentales presentaron un aumento en la población fúngica del suelo al final del experimento, pero ninguna superó al control, donde el microbioma incremento significativamente.
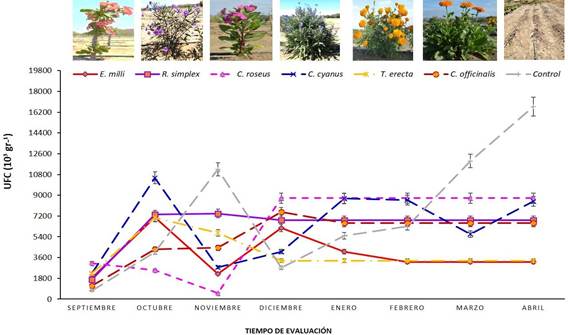
Figura 1 Población fúngica (Unidades Formadoras de Colonias) de la rizósfera de las plantas ornamentales.
Esta respuesta, podría estar asociada tan solo a la condición de humedad en el suelo para activar la población microbiana, y donde las plantas ornamentales poseen una influencia en la regulación de la población fúngica posiblemente debido a los metabolitos secundarios que secretan sus raíces. Esto concuerda con Ma et al. (2022) quienes reportan que los exudados radiculares contienen carbohidratos, aminoácidos, flavonoides y ácidos fenólicos, los cuales poseen diferentes efectos sobre los microorganismos de las raíces. Entre ellos, los aminoácidos y carbohidratos proporcionan fuentes eficientes de carbono y nitrógeno que influyen en el número y distribución de especies de las poblaciones microbianas del suelo. Así mismo, Sasse et al. (2018) añaden que los exudados de las raíces alteran gradualmente el microbioma al modificar las propiedades fisicoquímicas del suelo, al liberar grandes cantidades de ácidos orgánicos, lo que reduce el pH del suelo y afecta el crecimiento y la colonización microbiana. Además, los exudados de las raíces no solo solubilizan el fosfato, sino que también regulan la absorción microbiana de fósforo en la rizósfera. Además, la concentración y tipos de exudados dependen de la etapa fenológica de las plantas, lo que puede estar relacionado con la variación poblacional fúngica que se observó a través del tiempo en cada planta ornamental.
Caracterización morfológica. Un total de 25 aislados fúngicos fueron obtenidos con morfología colonial variada, al presentar texturas de tipo algodonosa, granulada y/o plana de forma circular o irregular en su crecimiento, donde éste último en algunas especies era rápido, moderado o lento. Además, presentaron distintas tonalidades de color los cuales variaron desde el verde oliva, verde ligero al verde intenso. Así como coloraciones blancas, amarillo, morado, naranja, negro, salmón, café y rosa (Cuadro 1). Dentro de la caracterización microscópica, los hongos presentaron formas diferenciales en sus esporas, micelio y cuerpo fructífero (Figura 2, 3 y 4), donde a partir de estas características se identificaron 11 especies del género Aspergillus, seis de Trichoderma, tres de Penicillium,
dos de Rhizopus, una de Morteriella spp., Fusarium oxysporum y Geotrichum spp. La mayoría de los hongos identificados pertenecen a los grupos de tipo saprófitos donde destacan los géneros Aspergillus spp., Penicillium spp., Trichoderma spp. y Rhizopus spp. Esto concuerda con lo reportado por Nicoletti y Trincone (2016), que mencionan que Aspergillus y Penicillium representan respectivamente el primero y segundo género de mayor abundancia de hongos filamentosos reportados. Fue evidente la variabilidad de colores de cada hongo, donde éstos están asociadas a la resistencia ambiental y al tipo de alimentación del microorganismo en su nicho ecológico. De acuerdo con Pombeiro et al., (2017), los pigmentos fúngicos relacionados a las melaninas se asocian al color gris, antraquinonas hidroxiantraquinonas y quinonas al color granate, bronce y naranja, azafilonas y oxopolieno al color amarillo, morado y rojo, carotenoides al naranja y amarillo y naftoquinonas al rojo y morado.
Cuadro 1 Morfología de hongos rizosféricos asociados a plantas ornamentales.
| Especie | Colonia | Micelio | Esporas | ||
|---|---|---|---|---|---|
| Anverso | Reverso | Textura | Crecimiento | ||
| Aspergillus C-1 | Morado | Morado | Plana | Lento | Globosas |
| Aspergillus C-3 | Verde | Amarillo | Plana | Moderado | Globosas |
| A. orizae | Verde | Verde | Plana | Moderado | Globosas |
| A. terreus | Salmon | Amarillo | Plana | Moderado | Globosas |
| A. niger C-1 | Negro | Negro | Plana | Moderado | Globosas |
| A. niger C-2 | Negro | Negro | Granulada | Moderado | Globosas |
| A. candidatus | Blanca | Café | Algodonosa | Moderado | Globosas |
| A. flavus | Verde oliva | Blanco | Plana | Moderado | Globosas |
| A. ochraceus | Amarillo | Crema | Granulado | Moderado | Globosas |
| A. flavipes | Blanca | Crema | Estriada | Lento | Globosas |
| Trichoderma spp. | Amarilla | Amarillo | Granular | Rapido | Globosas |
| T. atroviride | Verde ligero | Blanco | Algonosa | Rapido | Subglobosas |
| T. harzianum | Verde | Blanco | Algodonosa | Rápido | Globosas |
| T. atroviride C-1 | Verde | Verde | Plana | Rapido | Globosas |
| T. atroviride C-2 | Verde ligero | Blanco | Algodonosa | Rapido | Globosas |
| T. asperellum | Verde ligero | Blanco | Algodonosa | Rapido | Globosas |
| Penicillium C-1 | Naranja | Naranja | Plana | Moderado | Oblongas |
| Penicillum C-2 | Amarilla | Naranja | Plana | Moderado | Globosas |
| P. digitatum | Verde ligero | crema | Plana | Moderado | Globosas |
| Rhizopus spp. | Cafe | Cafe | Algodonosa | Rapido | Globosas |
| R. stolonifer | Gris | Gris | Plana | Rapido | Subglobosas |
| Morteriella spp. | Blanca | Blanco | Plana | Lento | Globosas |
| Geotrichum spp. | Blanca | Crema | Plana | Rapido | Bacilares |
| F. oxysporum | Rosa | Rosa | Plana | Moderado | Catenuladas |
Diversidad y riqueza fúngica del suelo. La diversidad fúngica indicó la presencia de dos filos (Ascomycota y Mucoromycota), seis familias (Trichocomaceae, Nectriaceae, Morteriellaceae, Hypocreaceae, Endomycetaceae y Mucoraceae) y siete géneros. El filo Ascomycota fue de mayor predominancia (71 % de frecuencia), comparado al filo Mucoromycota (29 %). Los resultados revelaron que las comunidades fúngicas en las muestras de suelo colectadas de cada planta ornamental fueron diferentes en el tiempo de evaluación (inicial y final), y se observó un cambio no solo en la diversidad de la microbiota de las ornamentales en la evaluación inicial y final, sino también en la frecuencia de ocurrencia, modificando la densidad poblacional por especie de hongo.

Figura 2 Microbiota correspondiente al género Aspergillus presente en la rizósfera de plantas ornamentales.

Figura 3 Microbiota correspondiente al género Trichoderma y Penicillium de la rizósfera en plantas ornamentales.

Figura 4 Microbiota correspondiente a los géneros de menor diversidad de la rizósfera en plantas ornamentales.
En el caso de C. cyanus, registró la mayor diversidad de especies fúngicas al final del experimento, al presentar siete aislados al inicio y 13 aislados al final del estudio, donde destacaron los géneros Aspergillus, Penicillium, Rhizopus y Trichoderma. Le siguió en orden de importancia, E. mili al registrar al inicio siete aislados y posteriormente 12. Así mismo, C. officinalis registró siete aislados y al final aumentó a nueve aislados. En R. simplex y C. roseum el número de aislados se mantuvo en siete y nueve aislados respectivamente. Mientras que en Tagetes (12-8 aislados) y el tratamiento control (9-6 aislados), la diversidad disminuyó (Figura 5). La diversidad presente en este tipo de especies reporta que, a pesar de las características en zonas áridas, como altas temperaturas, baja cantidad de materia orgánica, así como baja humedad, no impactaron en los géneros Aspergillus, Trichoderma y Penicillium. Esta diversidad es relevante para las plantas establecidas, como lo señala Reinhart y Callaway (2006) al mencionar que las interacciones entre las plantas y los microorganismos del suelo son de naturaleza dinámica y se basan en presiones coevolutivas. Además, Batten et al. (2006) señalan que las comunidades microbianas rizosféricas difieren entre las especies de plantas, entre los genotipos dentro de las especies y entre las diferentes etapas de desarrollo de las plantas, aunque no está claro si esto se debe a una mayor heterogeneidad del hábitat, el aumento de la biomasa vegetal comúnmente observado con comunidades de plantas muy diversas o una mayor diversidad de sustratos de carbono y compuestos de señalización proporcionada por las plantas. Fitzpatrick et al. (2018), señalan que el microbioma fúngico, se divide en tres categorías (mutualistas, patógenos y saprófitos) en función del tipo de planta, la cual a la vez depende de las interacciones antagónicas y sinérgicas dentro del microbioma del suelo y está confirmado que las plantas que crecen en un suelo con un microbioma común pueden alterar la abundancia relativa de mutualistas, patógenos y saprófitos en ese suelo.
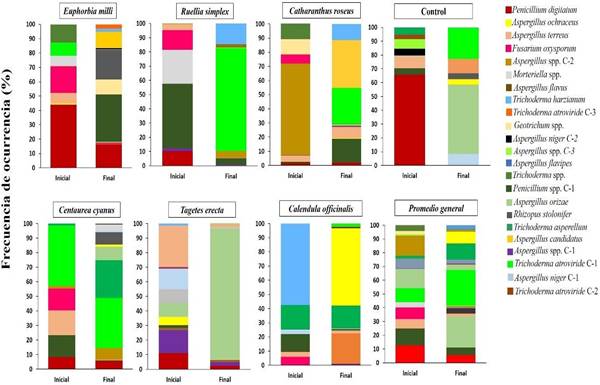
Índice de Shannon y Simpson. Las estimaciones de diversidad en las plantas ornamentales se presentan en el Cuadro 2. Los índices de Shannon y Simpson fueron utilizados para revelar la diversidad y riqueza presente en la rizósfera de las plantas ornamentales. E. mili y C. cyanus presentaron 14 aislados diferentes, mientras que T. erecta mostró ocho especies identificadas. Además, se registró que en E. mili las especies que presentaron mayor proporción fueron Penicillium spp. C-1 (33 %), Rhizopus stolonifer (20.8 %) y Penicillium digitatum (16.5 %). Mientras que en C. cyanus se registraron especies de Trichoderma atroviride C-1 (34 %) y Trichoderma asperellum (25 %). En T. erecta, se registró la mayor proporción en la especie Aspergillus oryzae (90
%). La diversidad calculada a través del índice de Shannon mostró similitud en las especies E. mili y C. cyanus mientras que T. erecta mostró el valor más bajo comparado con el resto de ornamentales. Mientras que C. officinalis y C. roseus presentaron al final una diversidad de aislados similares (13 aislados). Por su parte la dominancia de especies que fue medida por el índice de Simpson mostró que C. cyanus registró el mayor valor, mientras que T. erecta mostró el valor mínimo. De acuerdo con Margalef (1972) el índice de Shannon comprende valores de 1-5 donde valores menores a dos indican una diversidad baja. Asimismo, Silva et al. (2021) mencionan que el índice Shannon incrementa conforme se presenten un mayor número de sus especies y la proporción de individuos sea más homogéneo. En el índice de Simpson Soler et al. (2012) indican que este crece o disminuye según aumente o decrezca la diversidad y es en realidad un índice de dominancia que sobrevalora las especies más abundantes de la riqueza total. Magurran (1998) añade que entre más cercano a cero este el valor, existe mayor posibilidad de dominancia de una especie y de una población, y cuanto más cercano a la unidad hay menor dominancia. Como fue en R. simplex y T. erecta, donde en ambas se registró baja diversidad y baja dominancia de especies.
Cuadro 2 Diversidad y riqueza fúngica en la rizósfera de plantas ornamentales.
| Especie ornamental | Índice de Shannon | Índice de Simpson |
|---|---|---|
| Euphorbia mili | 1.8774 | 3.9072 |
| Ruellia simplex | 0.7976 | 1.6496 |
| Catharanthus roseus | 1.6670 | 4.4036 |
| Centaurea cyanus | 1.8823 | 4.8416 |
| Tagetes erecta | 0.4890 | 1.2312 |
| Calendula officinalis | 1.2973 | 2.6898 |
| Tratamiento control | 1.4187 | 3.0936 |
Propiedades químicas del suelo. No se presentaron diferencias significativas en cuanto al contenido del fósforo tanto en el análisis de suelo inicial como en el análisis final asociado a las seis especies de ornamentales y el control. En Materia orgánica C. officinalis mostró los valores máximos (1.07 %), mientras que en R. simplex fue menor (0.45 %). Así mismo, E. mili, R. simplex y C. roseus mostraron diferencia significativa con respecto a C. cyanus, T. erecta, C. officinalis y el control. En Potasio se mostró que
R. simplex mostró el menor contenido respecto al resto de ornamentales (33.11 ppm,) y el control (101.4 ppm). Además, se observó que R. simplex mostró diferencia significativa con las especies C. cyanus, T. erecta, C. officinalis y tratamiento control mientras que las especies E. mili y C. cyanus no mostraron diferencia significativa respecto al resto de los tratamientos. En el contenido de Fósforo, Calcio, Magnesio y Conductividad eléctrica no se presentaron diferencias significativas. En el caso R. simplex, E. mili y C. roseus donde la materia orgánica fue menor, esto se puede asociar a que éstas se encuentran en constante producción de biomasa asociada a hojas, ramas y flores (Figura 6). Esto concuerda con Adamczyk (2021) quien indica que una mayor proporción de raíces puede mejorar la cantidad y diversidad de compuestos orgánicos como el nitrógeno, lo que mejora la composición, el aumento y estabilidad de la materia orgánica del suelo, debido a que este tipo de compuestos secretados por las raíces ayudan a desdoblar las moléculas y/o iones asociados a elementos químicos presentes en la materia orgánica del suelo. Así mismo, Coskun et al. (2017) señalan que ciertos exudados pueden inhibir la nitrificación al inhibir selectivamente la actividad de bacterias nitrificantes del suelo, lo que ralentiza la velocidad de reacción a la que el amonio se convierte en nitrato en el suelo.
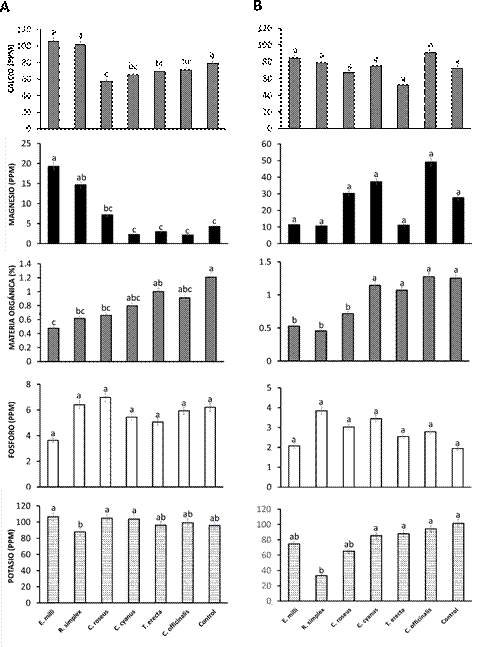
Figura 6 Efecto de plantas ornamentales en los nutrientes del suelo. A) Análisis inicial y B) final. Los valores son medias ± desviación estándar. a b c d; las medias seguidas por la misma letra para un factor dado no son significativamente diferentes (p < 0.05; prueba HSD de Tukey).
En el contenido de potasio Calendula officinalis presentó el mayor valor, esto posiblemente se asoció a los metabolitos secundarios exudados por las raíces de esta planta, tal como señala Ma et al. (2022) quienes indican que los exudados radiculares ricos en ácidos orgánicos pueden ejercer un efecto liberador de potasio sobre los minerales que contienen potasio. Esta respuesta en la variación del contenido de Potasio, también puede deberse a que algunas especies ornamentales utilizaron en mayor demanda este elemento para la resistencia a la sequía, la constante formación de raíces, así como la continua floración que se presentó en cada una de las especies ornamentales. Al respecto, Srivastava et al. (2019) indican que el potasio ayuda en la producción de azúcares, así como en la formación y síntesis de proteínas y reduce la pérdida de agua. Por su lado Anschütz et al. (2014) explican que el potasio ayuda en la resistencia a sequía en las plantas, así como en el estrés a las altas temperaturas. En el caso del resto de los parámetros no se mostraron cambios debido posiblemente a que se necesitaba mayor tiempo de evaluación o bien que todas las especies ornamentales no demanden una cantidad mayor de estos elementos como fue el caso de materia orgánica y potasio.
Análisis de correlación entre la diversidad fúngica y las propiedades físico-químicas del suelo
Los patrones encontrados en los diagramas de correlación de Person indican que, tanto en el índice de Shannon como en el índice de Simpson, las características edafológicas del suelo (Figura 7 y 8), se correlacionaron negativamente con el tipo de planta ornamental. Resultados similares de correlación negativa fueron reportados por Pacasa- Quisbert et al. (2017), al evaluar el efecto del tipo de uso del suelo, tipo de vegetación nativa y tipo de cultivo sobre la diversidad y número de hongos relacionados con parámetros físicos y químicos del suelo (pH, textura, materia orgánica), donde concluyeron que la abundancia microbiana está altamente relacionada con otros factores como; el tipo de suelo, características climáticas, otros microorganismos y hasta el tipo de comunidades de plantas.
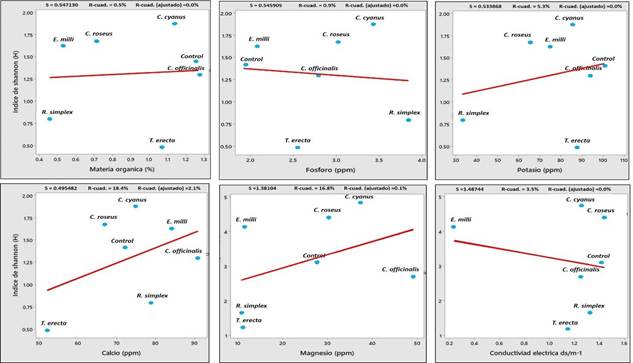
Figura 7 Correlación entre el índice de diversidad fúngica de Shannon y las características del suelo asociadas a plantas ornamentales.
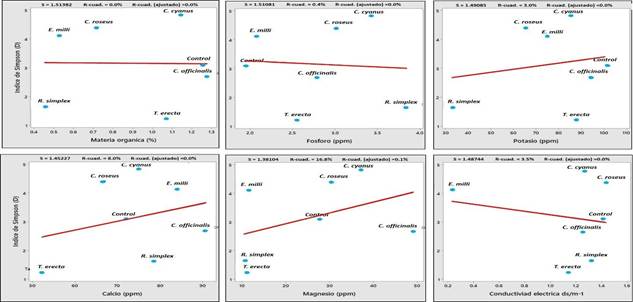
Conclusiones
Se determina que el tipo de especies de plantas ornamentales influyeron en la diversidad y riqueza de la microbiota fúngica, así como en algunas propiedades fisicoquímicas del suelo. Sin embargo, no se observó una relación positiva en la correlación de las tres variables. Ruellia simplex fue la planta ornamental con menor diversidad de hongos (7 aislados). Mientras que la planta asociada a C. cyanus presentó el mayor aumento en la diversidad y riqueza fúngica (13 aislados). Así mismo, las plantas ornamentales R. simplex y T. erecta favorecieron la presencia de hongos benéficos en el suelo al presentarse mayor cantidad de los géneros Trichoderma spp. y Aspergillus spp. Por lo que se puede señalar que algunas plantas ornamentales influyen positivamente en la riqueza y diversidad fúngica.

