











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
La selva tropical caducifolia (STC) abarca cerca del 40% de los bosques tropicales a nivel mundial (FAO, 2001). Este tipo de selva se ubica principalmente en regiones cálidas caracterizadas por una variación climática estacional ampliamente marcada (Holdridge, 1967; Rzedowski, 2006). Durante la estación seca en la STC, la mayoría de los árboles pierden sus hojas generando espacios abiertos en el dosel y en consecuencia se origina una amplia variación microclimática. Mientras que, en la estación de lluvias, la cobertura del dosel se restablece provocando la estabilización de factores como la temperatura y la humedad relativa (Trejo, 1999). La variación en la estructura de la vegetación y las condiciones microclimáticas entre ambas estaciones, puede generar cambios en la disponibilidad de los recursos necesarios para los vertebrados. Por ejemplo, se ha señalado que los sitios de reproducción y regulación térmica, así como la disponibilidad de alimento pueden variar entre la estación de seca y húmeda (Mandujano & Gallina, 1995; García & Cabrera-Reyes, 2008).
Los anfibios y reptiles son algunos de los vertebrados que pueden responder a la variación estacional de la STC, debido a que son altamente sensibles a los cambios en sus microhábitats (Gibbons et al., 2000; Pough, 2007; Pyron, 2018) y dependen del microclima (Paternina, 2016). Asimismo, varias especies de la herpetofauna necesitan de cuerpos de agua temporales o permanentes para su reproducción, desplazamiento y alimentación (Wells, 2007; Vitt & Caldwell, 2013). Sin embargo, se ha reportado que las ranas, sapos, salamandras y cecilias pueden ser más sensibles que los reptiles a las modificaciones ambientales que genera la variación estacional, debido a su piel desnuda y permeable, y una mayor dependencia a la humedad (García, 2003; García & Cabrera-Reyes, 2008). Al presentar este tipo de piel, los anfibios pueden presentar altos niveles de estrés fisiológico cuando las temperaturas son altas durante la estación seca (Androne et al., 2001; Leyte-Manrique et al., 2016).
Las diferentes respuestas de la herpetofauna a la estacionalidad en las selvas secas pueden compararse a través del análisis de la diversidad taxonómica y sus componentes: riqueza de especies, abundancia y composición de especies (Duellman & Thomas, 1996; Peña-Joya et al., 2018; Roth-Monzón et al., 2018). Por ejemplo, en tres selvas tropicales caducifolias de México se encontró que la riqueza y la abundancia de los anfibios disminuyeron drásticamente en la estación seca en comparación con la estación de lluvias. Mientras que la riqueza y la abundancia de los reptiles registraron patrones contrastantes entre ambas estaciones (García & Cabrera-Reyes, 2008; Rioja-Paradela et al., 2013; Leyte-Manrique et al., 2016). Además, se ha reportado que dentro de cada grupo biológico también existen diferentes respuestas a la estacionalidad, asociadas principalmente a los rasgos morfológicos, fisiológicos o de historia de vida de las especies (Keinath et al., 2016).
Leyte-Manrique et al. (2016) reportaron la presencia de seis especies de anuros en la estación de lluvias en una STC de México, mientras que en la estación seca sólo reportaron una especie. En contraste, la riqueza de lagartijas puede ser semejante entre estaciones climáticas debido a su plasticidad y capacidad para explotar los recursos disponibles. Sin embargo, se ha detectado que la riqueza y abundancia de serpientes generalmente disminuye en la estación de secas, probablemente por la disminución de presas como mamíferos o aves (Aurioles et al., 2000; Marques et al., 2001; Vite-Silva et al., 2010; Roth-Monzón et al., 2018). Por lo tanto, estudiar la respuesta de la herpetofauna a la variación estacional, considerando los rasgos particulares de cada grupo biológico y cada especie, puede ayudar a comprender la dinámica temporal de las comunidades que habitan la STC.
Particularmente, la STC cubre aproximadamente el 12% del territorio mexicano (Ceballos et al., 2010). Sin embargo, la mayor parte de su cobertura vegetal ha sido deforestada para establecer zonas agrícolas y ganaderas (García Vázquez et al., 2006; Zepeda Gómez et al., 2017). Esta situación pone en riesgo a las comunidades de anfibios y reptiles, así como a sus interacciones ecológicas, de las que derivan importantes funciones ecológicas como: la dispersión de semillas, herbivoría, reducción de la turbidez del agua, y el flujo de energía a través de las cadenas tróficas (Cortes-Gómez et al., 2015). Por lo tanto, conocer la respuesta de la herpetofauna a la estacionalidad dentro de fragmentos de STC, puede ser importante para promover la conservación y recuperación de sus comunidades y funciones ecológicas. El objetivo de este estudio fue evaluar la respuesta de la diversidad taxonómica de los anfibios y reptiles a la variación estacional en una STC del centro de México. Particularmente, se comparó el número efectivo de especies, la abundancia, la equidad, y la composición de especies de cada grupo biológico entre la estación seca y la estación húmeda en una selva seca de la Sierra Mixteca de Puebla.
Materiales y métodos
Área de estudio. Se localiza en la localidad de La Sandía, municipio de Acatlán de Osorio dentro de la Sierra Mixteca de Puebla, México (18.20660° N, 98.147133° O; 1411 m s.n.m.; Fig. 1). La vegetación original corresponde a la STC, representada principalmente por especies de los géneros Bursera, Ipomoea, Tillandsia, Mimosa y Solanum (Rojas-Martínez & Flores-Olvera, 2019). El clima es cálido subhúmedo (Aw2) con una marcada estacionalidad climática (Guízar-Nolazco et al., 2010). Durante la estación de lluvias (junio - septiembre) la temperatura varía entre 15 y 27°C y la precipitación promedio es de 166 mm, mientras que en la estación seca (octubre - mayo) la temperatura varía entre 18 y 33°C y la precipitación promedio es de 13 mm (García, 1988; Guízar Nolazco et al., 2010; Martínez Espinosa, 2013). La STC se distribuye en fragmentos inmersos en zonas agrícolas de temporal (cacahuate, maíz, frijol y jamaica) y zonas de pastoreo (López Téllez et al., 2007; Guízar Nolazco et al., 2010).
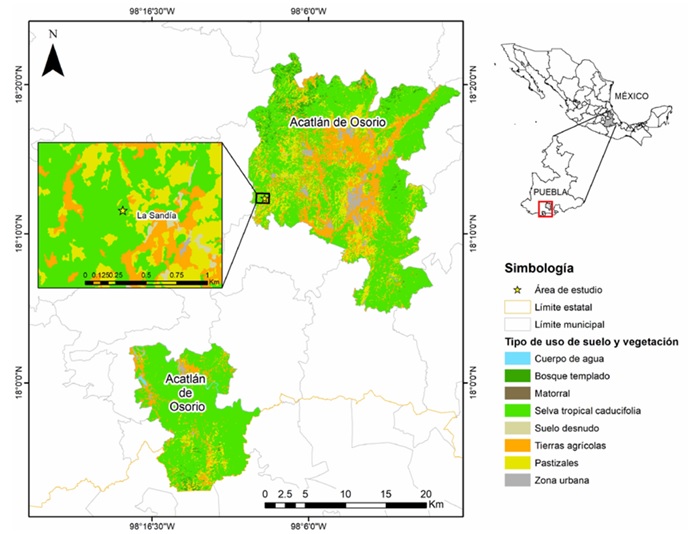
Figura 1 Ubicación del área de estudio, usos de suelo y tipos de vegetación en el municipio de Acatlán de Osorio, Puebla.
Registro de la herpetofauna. Para registrar a los anfibios y reptiles, seleccionamos un fragmento de ~2ha, el cual fue visitado cuatro veces durante la estación seca y cuatro veces durante la estación húmeda de 2017. En cada visita, durante cuatro días se realizaron tres recorridos al azar en el día (10:00 a 15:00 h) y tres en la noche (19:00 a 00:00 h). Las horas de muestreo definidas incluyeron los períodos de mayor actividad de cada grupo biológico en selvas secas tropicales (Carvajal-Cogollo & Urbina-Cardona, 2008; García & Cabrera-Reyes, 2008; Vite-Silva et al., 2010; Leyte-Manrique et al., 2016). En cada recorrido, de tres a cinco personas buscaron a los anfibios y reptiles utilizando la técnica de búsqueda libre con captura manual, la cual consiste en revisar los microhábitats idóneos de las especies incluyendo debajo de piedras u hojarasca, cuerpos de agua, árboles y arbustos, entre otros (Aguirre-León, 2011). El esfuerzo de muestreo en las visitas donde participaron tres personas (3 visitas) fue de 30 horas/persona (10 horas × 3 personas), mientras que en los muestreos donde participaron cinco personas (1 visita) fue de 50 horas/persona (10 horas × 5 personas), para un total de 140 horas/persona en cada estación climática. Los individuos capturados fueron identificados en campo mediante claves taxonómicas (Oliver López et al., 2009; Canseco-Márquez & Gutiérrez-Mayén, 2010). Después de la identificación a nivel de especie, todos los individuos fueron liberados en su sitio de captura. El manejo de individuos bajo estudio, no afectó en ningún momento la integridad ni la condición de vida de los organismos y de su entorno.
Obtuvimos el estado de conservación de cada especie registrada considerando la Lista Roja de la IUCN (2024), el índice de vulnerabilidad ambiental (Wilson et al. 2013a,b) y la Norma Oficial Mexicana NOM-059-SEMARNAT-2010 (SEMARNAT, 2010).
Análisis de datos. Todos los análisis de datos se realizaron de forma independiente para cada grupo biológico. Para realizar comparaciones válidas de los números de Hill entre las estaciones climáticas, primero calculamos la cobertura de la muestra (
Donde f 1 es el número de singletons, f 2 es el número de doubletons y n es el número de individuos (Chao & Jost, 2012). Cuando las coberturas de la muestra son cercanas al 100% y similares entre las comunidades, las comparaciones de los números de Hill pueden realizarse directamente con los valores observados (Cultid-Medina & Escobar, 2019). Después, calculamos los números de Hill para cada grupo biológico y por estación, estos números representan a la riqueza de especies (N0), el número de especies comunes (N1) y número de especies dominantes en las comunidades (N2; Jost, 2006; Moreno et al., 2011). Para comparar los números de Hill entre estaciones, se calcularon los intervalos de confianza al 95%, cuando los intervalos de confianza no traslapan se puede inferir diferencia estadística (Chao & Jost, 2012). Estos análisis fueron realizados en el paquete iNEXT (Hsieh et al., 2016) de R versión 2.0.9 (R Core Team, 2017).
Para comparar la abundancia de especies, calculamos el número total de individuos registrados en todos los muestreos en cada estación climática, y comparamos los promedios entre estaciones mediante modelos lineales generalizados con distribución Poisson y función de enlace logarítmica (Díaz-García et al., 2020). Para comparar la equidad de las comunidades entre estaciones climáticas, calculamos el índice de equidad relativa de primer orden (1 0 RLE) siguiendo la fórmula 1 0 RLE = ln(N1) / ln (N0), donde N1 = número de especies comunes y N0 = riqueza de especies (Jost, 2010). Para comparar la composición de las comunidades de anfibios y reptiles entre las dos estaciones climáticas construimos curvas de rango-abundancia, y revisamos los cambios en la posición de cada especie entre las curvas de cada estación climática (Moreno et al., 2011).
Resultados
En total registramos 86 individuos de 10 especies de anuros (seis ranas y cuatro sapos) y 114 individuos de 25 especies de reptiles (13 serpientes, 11 lagartijas, y una tortuga dulceacuícola). La cobertura de la muestra para cada una de las estaciones climáticas fue ≤90% en ambos grupos biológicos (Cuadro 1), por lo que pudimos realizar las comparaciones con los valores observados. De acuerdo con una revisión de colecciones científicas (Texas Cooperative Wildlife Collections, Texas A & M University [TCWC], Kansas University [KU] y University of Ilinois [UIMNH]), reportamos a Rhadinaea hesperia (Bailey, 1940), Sceloporus jalapae (Günther, 1890) y Tantilla bocourti (Günther, 1895) como nuevos registros para la herpetofauna del municipio de Acatlán de Osorio. Asimismo, registramos 25 especies endémicas a México, el 98% de las especies registradas se encontraron en la categoría de Preocupación menor de la Lista Roja de la IUCN (2024), y siete especies en la categoría de Alta vulnerabilidad ambiental de acuerdo con Wilson et al. (2013a, b; Cuadro 2).
Cuadro 1 Riqueza, abundancia y cobertura de la muestra de las comunidades de anfibios y reptiles en dos estaciones climáticas registradas en la localidad de La Sandía, Acatlán de Osorio, Puebla.
| Riqueza de especies | Abundancia | Cobertura de la muestra | |
| ANFIBIOS | |||
| Secas | 10 | 29 | 90% |
| Lluvias | 8 | 57 | 93% |
| REPTILES | |||
| Secas | 19 | 78 | 91% |
| Lluvias | 18 | 66 | 90% |
Cuadro 2 Diversidad, endemismo y estado de conservación de las especies de anfibios y reptiles registradas en la estación seca y en la estación de lluvias en la localidad de La Sandía, Acatlán de Osorio, Puebla. La X indica a las especies endémicas a México. La columna IUCN indica el estado de conservación de acuerdo a la Lista Roja de la Unión Internacional para la Conservación de la Naturaleza (IUCN, 2024): NE = No Evaluada, LC = Preocupación menor, y EN = En peligro. La columna EVS indica el estado de vulnerabilidad ambiental de anfibios y reptiles mexicanos (Wilson et al. 2013a; b): Baja = Vulnerabilidad baja (valor de riesgo de 3-9), Media = Vulnerabilidad media (valor de riesgo de 10-13), y Alta = Vulnerabilidad alta (valor de riesgo de 14-20).
| ID | Estación de secas | Estación de lluvias | Total | Especie endémica a México | IUCN | NOM 059 | EVS | |
| ANFIBIOS | ||||||||
| Orden Anura | ||||||||
| Bufonidae | ||||||||
| Incilius perplexus | Ip | 3 | 1 | 4 | X | LC | Media | |
| Incilius marmoreus | Im | 1 | 6 | 7 | X | LC | Media | |
| Incilius occidentalis | Io | 2 | 2 | X | LC | Media | ||
| Craugastoridae | ||||||||
| Craugastor augusti | Ca | 1 | 1 | LC | Baja | |||
| Eleutherodactylidae | ||||||||
| Eleutherodactylus nitidus | En | 5 | 4 | 9 | X | LC | Media | |
| Hylidae | ||||||||
| Dryophytes arenicolor | Da | 2 | 18 | 20 | LC | Baja | ||
| Exerodonta smaragdina | Es | 5 | 6 | 11 | X | LC | Pr | Media |
| Phyllomedusidae | ||||||||
| Agalychnis dacnicolor | Ad | 1 | 12 | 13 | X | LC | Media | |
| Scaphiopodidae | ||||||||
| Spea multiplicata | Sm | 5 | 2 | 7 | LC | Baja | ||
| Ranidae | ||||||||
| Rana spectabilis | Ls | 4 | 8 | 12 | X | LC | Media | |
| Riqueza de especies | 10 | 8 | 10 | |||||
| Abundancia | 29 | 57 | 86 | |||||
| REPTILES | ||||||||
| Orden Squamata | ||||||||
| Lacertilia | ||||||||
| Dactyloidae | ||||||||
| Anolis microlepidotus | Am | 2 | 1 | 3 | X | LC | A | Alta |
| Gekkonidae | ||||||||
| Hemidactylus frenatus | Hf | 1 | 1 | LC | ||||
| Iguanidae | ||||||||
| Ctenosaura pectinata | Cp | 7 | 2 | 9 | X | LC | A | Alta |
| Phyllodactylidae | ||||||||
| Phyllodactylus bordai | Pb | 3 | 3 | X | LC | Pr | Media | |
| Phrynosomatidae | ||||||||
| Urosaurus bicarinatus | Ub | 17 | 9 | 26 | X | LC | Media | |
| Sceloporus horridus | Sh | 12 | 4 | 16 | X | LC | Media | |
| Sceloporus jalapae | Sj | 1 | 1 | X | LC | Media | ||
| Phrynosoma taurus | Pt | 1 | 1 | 2 | X | LC | A | Media |
| Teiidae | ||||||||
| Aspidoscelis sackii | As | 1 | 1 | LC | Alta | |||
| Aspidoscelis deppii | Ad | 10 | 4 | 14 | LC | Baja | ||
| Aspidoscelis costata | Ac | 8 | 8 | X | NE | Pr | Media | |
| Ophidia | ||||||||
| Colubridae | ||||||||
| Trimorphodon tau | Tt | 2 | 2 | X | LC | Media | ||
| Trimorphodon biscutatus | Tbi | 1 | 2 | 3 | LC | Baja | ||
| Oxybelis aeneus | Oa | 5 | 1 | 6 | LC | Baja | ||
| Pseudoleptodeira latifasciata | Pl | 3 | 3 | X | LC | Pr | Alta | |
| Pseudoficimia frontalis | Pf | 1 | 1 | X | LC | Media | ||
| Masticophis mentovarius | Mm | 7 | 2 | 9 | LC | A | Baja | |
| Tantilla bocourti | Tb | 1 | 1 | X | LC | Baja | ||
| Rhadinaea hesperia | Rh | 1 | 1 | X | LC | Pr | Media | |
| Sonora michoacanensis | Sm | 2 | 1 | 3 | X | LC | Alta | |
| Drymarchon melanurus | Dm | 1 | 1 | LC | Baja | |||
| Salvadora mexicana | Sme | 2 | 2 | X | LC | Pr | Alta | |
| Elapidae | ||||||||
| Micrurus laticollaris | Ml | 1 | 1 | X | LC | Pr | Alta | |
| Leptotyphlopidae | ||||||||
| Rena maxima | Rm | 2 | 2 | 2 | X | LC | Media | |
| Orden Testudines | ||||||||
| Kinosternidae | ||||||||
| Kinosternon integrum | Ki | 1 | 22 | 23 | X | LC | Pr | Media |
| Riqueza de especies | 19 | 18 | 25 | |||||
| Abundancia | 78 | 66 | 144 |
Número efectivo de especies. No encontramos diferencias significativas en la riqueza de especies (N0), el número de especies comunes (N1) y el número de especies dominantes (N2) en las comunidades de anfibios (Fig. 2a) y de reptiles (Fig. 2b) registrados en la estación seca y la estación de lluvias.
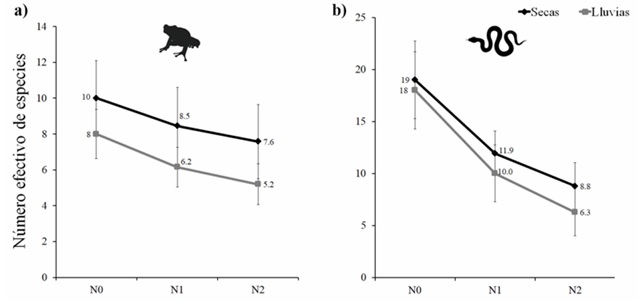
Figura 2 Riqueza de especies (N0), número de especies comunes (N1), y número de especies dominantes (N2) de las comunidades de anfibios (a) y reptiles (b) en dos estaciones climáticas en la selva tropical caducifolia de La Sandía, Acatlán de Osorio, Puebla. Los marcadores y líneas en color negro muestran los valores para la estación de secas, y en color gris para la estación de lluvias. Los marcadores indican los valores observados para cada orden de N, mientras que las barras de error indican los intervalos de confianza al 95%.
Abundancia. La abundancia de anfibios varió significativamente entre estaciones climáticas (Devianza = 9.28, grados de libertad = 1, p < 0.01), siendo mayor en la estación de lluvias que en secas (Fig. 3a). La abundancia de reptiles no varió significativamente entre estaciones climáticas (Devianza = 1.01, grados de libertad = 1, p > 0.05, Fig. 3b).
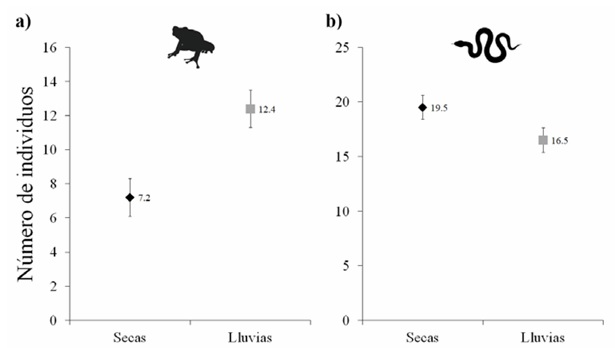
Figura 3 Abundancia de anfibios (a) y reptiles (b) registrada en la estación seca y en la estación de lluvias en la selva tropical caducifolia de La Sandía, Acatlán de Osorio, Puebla. Los marcadores y líneas en color negro muestran los valores para la estación de secas, y en color gris para la estación de lluvias. Para cada estación climática se muestra la media (marcadores) y el error estándar (barras de error).
Equidad y composición de especies. La comunidad de anfibios durante la estación seca presentó una distribución de las abundancias entre las especies más homogénea (1 0 RLE = 0.84) que durante la estación de lluvias (1 0 RLE = 0.67). En la estación seca no observamos ninguna especie dominante, pero en la estación de lluvias observamos dos ranas arborícolas dominantes, en primer instancia Dryophytes arenicolor (Cope, 1866), seguida por Agalychnis dacnicolor (Cope, 1864). La abundancia de estos dos anuros fue escasa en la estación seca. Se registró al sapo Incilius occidentalis (Camerano, 1879) y a la rana Craugastor augusti (Dugès, 1879) como especies exclusivas en la estación seca. En la estación de lluvias no se registraron especies exclusivas (Fig. 4).
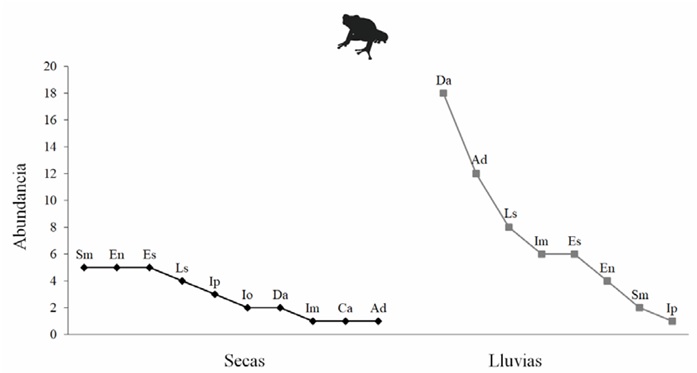
Figura 4 Curvas de rango abundancia de los anfibios registrados en la estación seca y en la estación de lluvias en la selva tropical caducifolia de La Sandía, Acatlán de Osorio, Puebla.
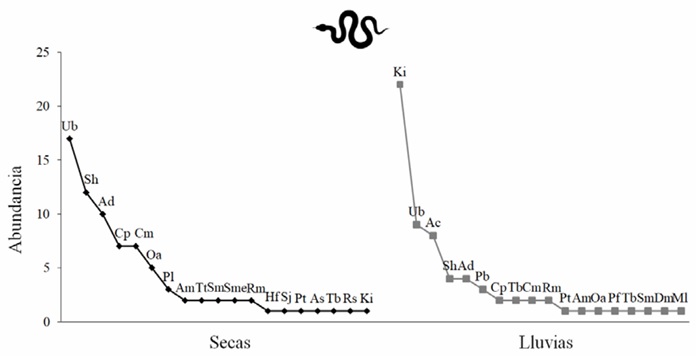
La comunidad de reptiles presentó una distribución heterogénea con respecto a la abundancia en ambas estaciones (Secas 1 0 RLE = 0.62; Lluvias 1 0 RLE = 0.55). Sin embargo, para cada especie se observaron cambios en la abundancia entre estaciones. En la estación seca observamos la dominancia de las lagartijas Urosaurus bicarinatus (Duméril, 1856), Sceloporus horridus (Wiegmann, 1834) y Aspidoscelis deppii (Wiegmann, 1834). En la estación de lluvias, la especie dominante fue la tortuga Kinosternon integrum (Le Conte, 1854). Esta especie de tortuga dulceacuícola sólo fue registrada con un individuo en la estación seca. Ambas estaciones compartieron once especies de reptiles y se encontraron especies únicas para cada estación. Las especies exclusivas en la estación seca fueron A. sackii (Wiegmann, 1834), Hemidactylus frenatus (Schlegel, 1836), Pseudoleptodeira latifasciata (Günther, 1894), Rena hesperia, Salvadora mexicana (Duméril, Bibron & Duméril, 1854), Sceloporus jalapae, y Trimorphodon tau (Cope, 1869). Las especies exclusivas en la estación de lluvias fueron A. costata (Cope, 1878), Drymarchon melanurus (Duméril, Bibron & Duméril, 1854), Micrurus laticollaris (Peters, 1869), Phyllodactylus bordai (Taylor, 1942), Pseudoficimia frontalis (Cope, 1864) y Tantilla bocourti (Fig. 5).
Discusión
Este estudio contribuye al conocimiento de la diversidad de los anfibios y reptiles en la Mixteca poblana reportada en otros estudios (Aurioles et al., 2000; Pérez & Feria, 2000; Chacón et al., 2002; Chong & Yanes, 2002; Navarro et al., 2002; Maceda-Cruz, 2005). La herpetofauna de la localidad de La Sandía representa aproximadamente el 55% de las especies documentadas en la Mixteca Poblana (García Vázquez et al., 2006). Asimismo, el 72% de las especies registradas en este estudio son endémicas de México, lo que resalta la importancia de esta localidad y tipo de ecosistema para su conservación. Se identificaron tres especies como nuevos registros para el municipio de Acatlán de Osorio: la lagartija S. jalapae y las serpientes T. bocourti y R. hesperia. Mientras que las dos primeras ya habían sido registradas en otras localidades de la Mixteca poblana (Vargas Orrego, 2014; López Vivanco, 2015), R. hesperia representa el primer registro para la región, aumentando el conocimiento de la distribución y diversidad de especies de herpetofauna a escala regional.
Los resultados de este estudio no revelaron diferencias significativas en la riqueza de especies de la herpetofauna entre las estaciones climáticas. Este hallazgo contrasta con los resultados de otros estudios en la misma región (Aurioles et al., 2002; Maceda-Cruz, 2005; Vargas-Orrego, 2014; López-Vivanco, 2015; Hernández-Ayotla, 2018), así como en áreas con vegetación similar en Guanajuato (Leyte-Manrique et al., 2018), Jalisco (García & Cabrera-Reyes, 2008; Suazo-Ortuño et al., 2011; Peña-Joya et al., 2018), Oaxaca (Rioja-Paradela et al., 2013) e Hidalgo (Vite-Silva et al., 2010; Roth-Monzón et al., 2018). Estos estudios muestran que la riqueza de anfibios y reptiles es mayor durante la estación de lluvias en comparación con la estación seca. Este patrón podría atribuirse a que realizamos un menor número de muestreos en cada estación climática con respecto a otros estudios, por lo que la probabilidad de encontrar especies con un estrecho período de actividad pudo estar limitada. Es necesario que futuros estudios consideren ampliar el período de muestreo, tomando en cuenta que las especies pueden presentar diferentes patrones de actividad relacionados con la amplia gama de condiciones que puede generar cada estación climática en la STC.
Las selvas bajas caducifolias, caracterizadas por su baja precipitación y altas temperaturas imponen un estrés fisiológico en ciertas especies, que puede influir en su actividad reproductiva y en consecuencia pueden verse afectadas sus poblaciones (García & Cabrera-Reyes, 2008; Luría-Manzano, 2012). Los resultados de este estudio sugieren que la abundancia de anuros puede ser más sensible en comparación con la abundancia de los reptiles a las variaciones ambientales generadas por la estacionalidad de la STC, un hallazgo consistente con estudios previos en la Mixteca poblana (Vargas Orrego, 2014; López Vivanco, 2015). Asimismo, el modelo de distribución de las abundancias de los anfibios varió entre temporadas, resaltando que durante la temporada de secas no se detectó algún patrón de dominancia, contrario a los modelos clásicos que se caracterizan por una distribución sesgada donde hay muchas especies raras y sólo unas pocas especies comunes (Verberk, 2011). Esto podría deberse a la elevada susceptibilidad de las ranas y sapos a los cambios en la disponibilidad de agua (Pough et al., 2007), dado que muchas especies registradas dependen de cuerpos de agua para su reproducción, especialmente aquellas de las familias Hylidae, Ranidae y Bufonidae (Gual Díaz & Rendón Correa, 2014). Además, la exposición directa y prolongada a la radiación solar y la limitada tolerancia a las fluctuaciones de humedad y temperatura atmosférica que supone la estación seca, podrían contribuir a la respuesta encontrada en la comunidad de anuros (Duellman & Trueb, 1994). Anteriormente, se ha reportado que los anfibios dependen de hábitats con un dosel cerrado y suelos cubiertos de hojarasca, como el que puede representar la STC en la estación de lluvias, debido a que estas características de la vegetación proporcionan condiciones de alta humedad atmosférica, temperaturas constantes y disponibilidad de presas (Díaz-García et al., 2020).
En el caso de los reptiles, no detectamos diferencias en la riqueza y abundancia entre estaciones climáticas. Esto puede atribuirse a que los escamosos no experimentan una restricción tan marcada por la humedad y disponibilidad de agua como los anuros (Leyte-Manrique et al., 2019). Además, su mayor tolerancia a las condiciones del hábitat podría derivar de las adaptaciones conductuales y fisiológicas que este grupo ha desarrollado (Pechmann & Wilbur, 1994; García & Cabrera-Reyes, 2008; Jellinek et al., 2014). Por ejemplo, se ha sugerido que en ambientes cálidos los reptiles tienen una mayor capacidad de termorregular que en ambientes fríos (Lara-Reséndiz, 2020). Además, tienen la capacidad de poner huevos protegidos contra la desecación, lo que les proporciona una ventaja en hábitats con temperaturas altas y contrastantes (Zug et al., 2001).
La variación estacional en las condiciones ambientales de La Sandía se vio reflejada principalmente en los cambios en la composición de cada grupo biológico. Por ejemplo, las especies de anfibios más abundantes en la estación de lluvias, D. arenicolor y A. dacnicolor, presentan una larva acuática de nado libre durante su ciclo de vida, por lo que son altamente dependientes de los cuerpos de agua (Duellman & Trueb, 1994; Canseco-Márquez et al., 2010; Leyte-Manrique et al., 2018), y su detectabilidad aumenta en esta estación (Soto-Sandoval et al., 2017). En la estación de secas registramos una alta abundancia de especies de hábitos generalistas como Spea multiplicata y Eleutherodactylus nitidus. Asimismo, el sapo I. occidentalis fue registrado únicamente en la estación de secas. Esto puede deberse a que las condiciones ambientales que genera la sequía representan un filtro ambiental donde pocas especies se pueden ver favorecidas. En las regiones áridas y semiáridas de Puebla, los adultos de I. occidentalis prefieren habitar en los matorrales áridos y semiáridos, y generalmente evitan cuerpos de agua permanentes, además mantienen su actividad hasta noviembre y principios de diciembre, coincidiendo con la estación seca (Santos-Barrera, 2014).
Para los reptiles, la tortuga K. integrum fue la especie dominante en lluvias, debido probablemente a que utiliza los cuerpos de agua temporales o permanentes para alimentarse y reproducirse (Macip Ríos et al., 2009), en contraste con la estación seca donde entra en un período de estiaje (Enríquez-Mercado, 2017). La dominancia de la lagartija Urosaurus bicarinatus en la estación seca, puede deberse a que esta especie presenta una producción de gametos sexuales en ambos sexos, definida por el incremento del fotoperiodo pero que no se ve afectada por la temperatura (Ramírez-Bautista & Vitt, 1998). Otras especies en las que reportamos un incremento en sus poblaciones con la sequía, fueron S. horridus y A. deppii. La temperatura óptima de estas lagartijas oscila entre los 33 y 40 °C (Castro-Franco & Aranda, 1984; Vitt et al., 1993), siendo su termorregulación más eficiente en la estación seca que en la húmeda (Bustos-Zagal et al., 2013).
Nuestro estudio demuestra que las diferentes respuestas de la herpetofauna a la estacionalidad de la selva tropical seca en la Sierra Mixteca poblana, pueden ser explicadas por rasgos de historia natural de las especies, que incluyen características morfológicas, fisiológicas y reproductivas. Asimismo, resaltan la importancia de evaluar los diferentes atributos taxonómicos de las comunidades, para identificar patrones de cambio entre estaciones climáticas. Debe considerarse que la conversión de los fragmentos de selva a usos de suelo agrícola o ganadero, es la principal amenaza para la biodiversidad de las selvas tropicales caducifolias de México (Mesa-Sierra et al., 2022). Por tanto, es necesario diseñar e implementar medidas de conservación y manejo adecuadas para preservar los remanentes de STC en el centro de México, debido a que son refugio para las comunidades de anfibios y reptiles con alto porcentaje de endemismo.

