











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introduction
One of the main features of the tropical dry forests of Mexico is its pronounced seasonality, as well as high value of species richness, and several endemic and threatened species (Dirzo & Ceballos, 2010; Meave et al., 2012; Briones-Salas et al., 2016); therefore, this ecosystem had been considered a biodiversity hotspot (Olson & Dinerstein, 2002). In Oaxaca state, according to Ortiz-Pérez et al. (2004), these forests occupy 16 % of the territory. Still, in a similar way to the rest of the country, this ecosystem is threatened by high deforestation rate, forest fires, overhunting, wildlife trade, human population growth, and tourism development (Torres-Colín, 2004; Trejo, 2010).
In tropical dry habitats, water resource availability is a limiting factor that determines many aspects of species’ behavior and ecology and influences the aggregation of species and individuals in time and space during the dry season (Ayeni, 1975; Redfern, 2003), principally because there is a severe water shortage for several months of the year. Therefore, water scarcity has an impact on almost any aspect of the natural history of tropical vertebrates (Dias et al., 2014; Delgado-Martínez et al., 2022). Although many vertebrates obtain water directly from water bodies, meanwhile others have evolved the capacity to rehydrate from water contained in food, such as the white-tailed deer that consume wild plants that provide succulent flowers and fruits (Chapman, 1988; Villarreal & Marín 2005; Dias et al., 2014). The dry season imposes harsh environmental conditions due to water scarcity (O´Farrill et al., 2014). Moreover, due to orographic characteristics and topographic depressions of the Pacific Coastal Plain (one of the eight physiographic provinces of Oaxaca), there are areas with rainwater accumulation that are formed because their soil does not allow water drainage. Although there are no formal studies related to knowledge of the soil type in that region, we know that the soil type in some areas does not allow the drainage of rainwater, facilitating its accumulation in certain areas for long periods. These natural waterholes (NW) are locally known as “aguajes” and they are the main source of surface water in the region when water is scarce and high temperatures are frequent (Dávila et al., 2019).
Although the majority of the “aguajes” dry out during the dry season, the few NW that persist left have an important role for a variety of tropical vertebrates and might act as a keystone resource for their survival (Dávila et al., 2019). Recently in Mexico, López-Téllez et al. (2021) documented the visit of 20 vertebrates in NW during the dry season in the southeast of Puebla state, Mexico. Then, Delgado-Martínez et al. (2021) documented the use of water-filled three holes (dendrotelmata) by tropical vertebrates in the seasonal tropical forest of Calakmul Biosphere Reserve, in the southern Mexico.
Despite the Protected Areas are the main strategy to preserve biodiversity (Monterrubio-Solís, 2019), in Oaxaca state exists the Communal Natural Protected Areas (CNPAs), which operates as a mechanism for the conservation of biodiversity and local natural resources through the participation of local human communities (Rodríguez-Luna et al. 2011), little is known about the relationship between tropical vertebrates and NW in the tropical dry forest on the central coast of Oaxaca, so knowledge about the importance of these NW remains unknown in this region of the country. Therefore, this study aimed to evaluate the use of NW (aguajes) by vertebrates through their activity patterns in a tropical dry forest of the Communal Natural Protected Area El Gavilan (CNPA), central coast of Oaxaca, Mexico.
Materials and methods
Study area. The CNPA El Gavilán is in the municipality of Santa María Tonameca on the central coast of Oaxaca (Fig. 1), situated in the transition zone between the coastal plain and the Sierra Madre del Sur physiographic province. The area is characterized by an essentially plain topography with a mountain’s axis (NW-SE direction) perpendicular to the plain decline (Ortíz-Pérez et al., 2004) and pronounced slopes that make it difficult to transit vertically. Moreover, this area is under pressure due to various anthropogenic activities, such as high deforestation, overhunting, and human population growth around this site (García-Grajales et al., 2023).
The climate of this region is warm and sub-humid, with an annual mean temperature of 26.8 °C (Torres-Colín, 2004). The area is influenced by two distinctive seasons, wet (June-October) and dry (November-May), with a mean annual rainfall of 2 245 mm (García, 1973). Deciduous tropical forest, semideciduous tropical forest and shade coffee crops are the predominant land use and vegetation, but semideciduous tropical forest and shade coffee crops are in the form of patches with reduced surface (Torres-Colín, 2004).
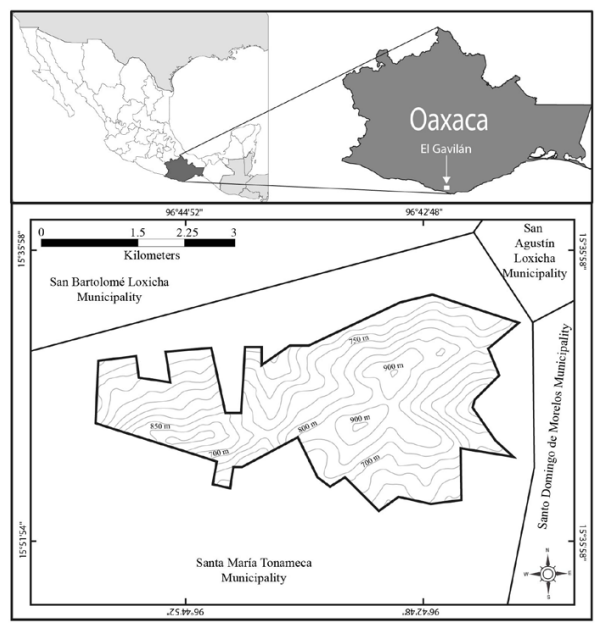
Figure 1 Geographical location of the Communal Natural Protected Area (CNPA) El Gavilán, Oaxaca, México.
Field sampling. The fieldwork was carried out from November 2016 to May 2017, during the dry season of this region. For this, eight NW were located in the CNPA El Gavilán that were at 715 m.a.s.l. on average (range: 330-1050 m; Table 1). All NW were in small depressions on steep mountainsides, with a minimum distance between them of 500 meters and were not accessed by cattle. The water source for all these waterholes was only rainwater from the respective season.
Table 1 List of natural waterholes selected into the Communal Natural Protected Area (CNPA) El Gavilán, Oaxaca, México. M.A.S.L.= Meters above sea level.
| Name of Waterhole | Code | M.A.S.L. |
| Venero Plátano Morado | NW 1 | 845 |
| Aguaje Amador | NW 2 | 965 |
| Rancho Amador | NW 3 | 1,050 |
| El Rieguito | NW 4 | 573 |
| Dos Higos | NW 5 | 330 |
| El Paso | NW 6 | 698 |
| Arroyo Grande | NW 7 | 480 |
| El Patio | NW 8 | 781 |
One camera trap (Stealth ® Cam unit Ops and Bushnell ® Trophy Cam HD models) with infrared and remote-trip digital camera units was placed at each NW. In addition, both camera models record the environmental temperature of the site where they have been deployed. The camera traps were placed at approximately 3 m from the edge of the waterhole, without bait, and at a height of 45 cm above the ground, tied on trees or poles at a suitable location to maximize encounters of species (Pin et al., 2018; Dávila et al., 2019). Moreover, we followed the procedures published by Dávila et al. (2019), each trap was programmed to remain active for 24 h and to take three pictures per trigger, with a one second interval between triggers. These three pictures were considered as one photo event (Monroy-Vilchis et al., 2011). Camera traps were also programmed to register the time of day at which each photo was taken, information which was later used to obtain temporal patterns of tropical vertebrates attending waterholes. To secure the information, data from the cameras was retrieved once a month and batteries were replaced every three months or sooner when it was necessary.
Photo-captures were separated for each waterhole and all vertebrate species were identified through specialized literature for reptiles (Köhler, 2011), birds (Van Perlo, 2006, Bojorges-Baños, 2011), and mammals (Reid, 1997), taking only those photographs that allowed to observe the vertebrate. Then, to minimize double counting and maximize the independence of photographic events at each waterhole, independent photographic records were considered when there were a) consecutive photographs of individuals of different species, b) consecutive photographs of individuals of the same species separated by more than six hours, because in a lower time there is a greater chance of counting the same individual, even more so in the case of birds (Dávila et al., 2019); or c) non-consecutive photographs of individuals of the same species. In the case of photographs of gregarious species where more than one individual was observed, the number of independent records considered was the same as the number observed in the image (Monroy-Vilchis et al., 2011).
In terms of endemism, all wild vertebrates were classified as endemic species (species with a geographical distribution limited to one country), quasi-endemic (species with a geographic distribution extends outside Mexico), and semi-endemic (species endemic to the country for a time of year) (González-García & Gómez de Silva, 2003) for a better understanding of their status.
Activity patterns in natural waterholes (“aguajes”). The 24-hour cycle was divided into one-hour segments and each record was obtained in the camera traps within these intervals.
Due to the duration of periods of the day is different for various authors, some of them take a twilight time interval with the duration of 120 minutes before and after sunrise and sunset, respectively (Bridges et al., 2004) or 60 minutes as well (Romero-Muñoz et al., 2010; Ross et al., 2013; Ikeda et al., 2016). In this study, four categories of activity were used: crepuscular morning (06:00-08:00), crepuscular evening (18:00-20:00), and the intervals between these were diurnal (08:00-18:00) and nocturnal (20:00-06:00) (Maffei et al., 2005, Monroy-Vilchis et al., 2011). If at least 50 % of the records of each species observed correspond to that time, then were classified in one of these categories. But, if the percentage were less than 50% of the records, then they were considered as cathemeral (Maffei et al., 2005; Monroy-Vilchis et al., 2011; Pérez-Irineo & Santos-Moreno, 2021). This last term (cathemeral) is applied to the pattern of an organism’s activity that occurs during both the light and dark portions of the 24-h cycle (Donati & Borgognini-Tarli, 2006). Thus, a cathemeral organism does not strictly adhere to a diurnal, nocturnal, or crepuscular phase, but rather shift its activity throughout the 24 h (Clarke et al., 1995; Curtis & Ramunsen, 2006).
In order to determine the frequency of use by species, the frequency of capture (FC; Karanth & Nichols, 1998) was calculated as: FC = Number of records from each sample X 100 / Number of camera-trap days. In addition, the time of the first detection of each species (latency time) was calculated as the number of days used between setting up the camera-traps and the first registration of each species captured (Monroy-Vilchis et al., 2011; Buenrostro-Silva et al., 2020).
A parametric one-way ANOVA test was used to examine differences among natural waterholes and the average time between visits of all species. Previously, the normality and homogeneity of variance of the data were checked with the Anderson-Darling test. Then, data were analyzed with Oriana software version 4 (Kovach, 2011) for circular statistics using the Rayleigh test of uniformity and Rao’s spacing test, to examine data that are distributed in a circular manner, such as time (Lehner, 1996).
Results
A total of 431 independent records were obtained from eight natural waterholes, with an accumulative sampling effort of 25,628 days-trap. Twenty-two species, belonging to three classes, 12 orders, 18 families, and 22 genera were recorded (Table 2,3). Birds were the group with the highest number (n=13) of species records, followed by mammals (n=8), and reptiles (n=1). Passeriformes and Columbiformes were the best-represented orders of birds in association with the NW, while Carnivora and Artiodactyla were the best-represented orders of mammals in the NW.
Table 2 Species list recorded in natural waterholes in Communal Natural Protected Area (CNPA) El Gavilán, Oaxaca, México.
| Class | Order | Family | Species |
| Reptilia | Sauria | Iguanidae | Iguana iguana |
| Galliformes | Cracidae | Ortalis poliocephala | |
| Patagioenas flavirostris | |||
| Columbiformes | Columbidae | Geotrygon montana | |
| Leptotila verreauxi | |||
| Cuculiformes | Cuculidae | Piaya cayana | |
| Strigiformes | Strigidae | Athene cunicularia | |
| Aves | Piciformes | Picidae | Aulacorhynchus prasinus |
| Falconiformes | Falconidae | Micrastur semitorquatus | |
| Calocitta formosa | |||
| Cervidae | Cyanocorax yncas | ||
| Passeriformes | Icteridae | Icterus graduacauda | |
| Quiscalus mexicanus | |||
| Cardinalidae | Cyanocompsa parellina | ||
| Artiodactyla | Cervidae | Odocoileus virginianus | |
| Tayassuidae | Dicotyles angulatus | ||
| Felidae | Leopardus pardalis | ||
| Carnivora | Procyonidae | Nasua narica | |
| Mammalia | Mustelidae | Mustela frenata | |
| Cingulata | Dasypodidae | Dasypus novemcintus | |
| Didelphomorphia | Didelphidae | Didelphis virginiana | |
| Rodentia | Sciuridae | Sciurus aureogaster |
Table 3 Records and frecuency of capture (FC) of species in natural waterholes in Communal Natural Protected Area (CNPA) El Gavilán, Oaxaca, México. En= Endangered, SP= Under special protection; En= Endemic species, NE= Non endemic species; NW= Natural waterhole (“aguaje”).
| Species | Total, of records | FC | NOM 059 | Endemic level | NW 1 | NW 2 | NW 3 | NW 4 | NW 5 | NW 6 | NW 7 | NW 8 |
| Iguana iguana | 2 | 0.01 | En | ✔ | ✔ | ✔ | ||||||
| Ortalis poliocephala | 31 | 0.12 | EN | ✔ | ✔ | ✔ | ✔ | |||||
| Patagioenas flavirostris | 23 | 0.09 | NE | ✔ | ✔ | ✔ | ||||||
| Geotrygon montana | 1 | 0.004 | NE | ✔ | ||||||||
| Leptotila verreauxi | 31 | 0.12 | NE | ✔ | ✔ | ✔ | ✔ | |||||
| Piaya cayana | 1 | 0.004 | NE | ✔ | ||||||||
| Athene cunicularia | 4 | 0.02 | NE | ✔ | ✔ | |||||||
| Aulacorhynchus prasinus | 4 | 0.02 | SP | NE | ✔ | |||||||
| Micrastur semitorquatus | 8 | 0.03 | SP | NE | ✔ | ✔ | ||||||
| Calocitta formosa | 6 | 0.02 | NE | ✔ | ✔ | ✔ | ||||||
| Cyanocorax yncas | 5 | 0.02 | NE | ✔ | ||||||||
| Icterus graduacauda | 1 | 0.004 | CE | ✔ | ||||||||
| Quiscalus mexicanus | 4 | 0.02 | NE | ✔ | ||||||||
| Cyanocompsa parellina | 2 | 0.01 | NE | ✔ | ||||||||
| Odocoileus virginianus | 20 | 0.08 | NE | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | |||
| Dicotyles angulatus | 29 | 0.11 | NE | ✔ | ✔ | ✔ | ✔ | ✔ | ||||
| Leopardus pardalis | 8 | 0.03 | SP | NE | ✔ | ✔ | ||||||
| Nasua narica | 148 | 0.58 | NE | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | ✔ | |
| Mustela frenata | 1 | 0.004 | NE | ✔ | ||||||||
| Dasypus novemcintus | 8 | 0.03 | NE | ✔ | ✔ | ✔ | ||||||
| Didelphis virginiana | 85 | 0.33 | NE | ✔ | ✔ | ✔ | ✔ | |||||
| Sciurus aureogaster | 9 | 0.04 | NE | ✔ | ✔ |
During the study period, April and May were the months with the greatest number of visits registered to the NW (Fig. 2). Of the eight NW, Venero Plátano Morado showed the biggest species record (n= 15 species); in contrast, El Patio recorded the lowest number of species (n= 3 species). Nasua narica was recorded in all NW, followed by Odocoileus virginianus that was recorded in six NW, meanwhile Dicotyles angulatus and Didelphis virginiana were recorded in five NW; nonetheless, five vertebrate species (Aulacorhyncus prasinus, Cyanocompsa parellina, Icterus graduacauda, Piaya cayana, Cyanocorax yncas), and one carnivore mammal (Mustela frenata) were exclusively recorded in the Venero Plátano Morado NW.
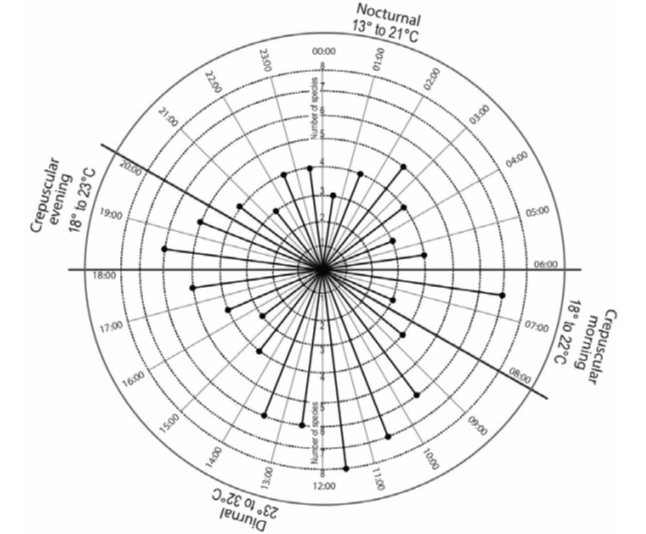
Figure 2 Records of visits per months in the natural waterholes (“aguajes”) of (CNPA) El Gavilán, Oaxaca, Mexico.
Of the total number of records, 88.93% were obtained during the day, 7.52% during crepuscular schedules (including morning and evening), and 3.55% during the night (Fig. 3). Most number of records were obtained between 11:00-12:00 h, with an average temperature of 28°C, while for the crepuscular schedules (morning and evening) the largest species records occurred between 06:00-07:00 hours (average 20°C) and 18:00-19:00 hours (average 23°C), respectively.
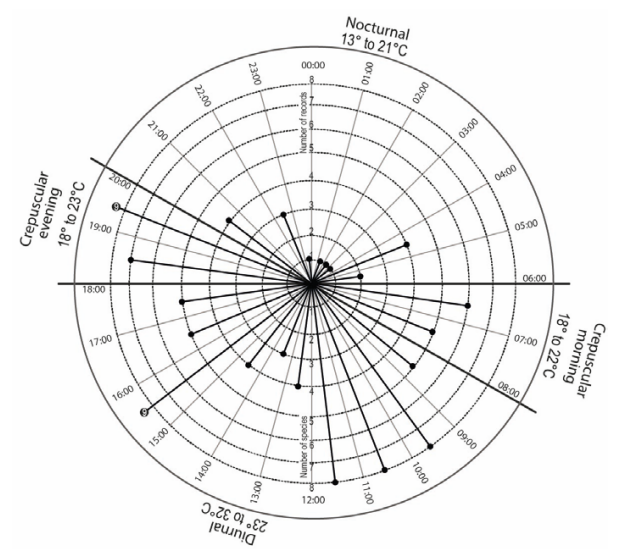
Figure 3 Number of independent records in natural waterholes (“aguajes”) of (CNPA) El Gavilán, during periods of 24 h, divide into 1 h segments.
Nasua narica was the species most frequently captured (FC= 0.58) in NW, followed by Orthalis poliocephala and Patagioenas flavirostris (FC= 0.12, respectively), while Geotrygon montana, Piaya cayana, Icterus graduacauda, and Mustela frenata were the least-used species of NW (Table 2). Moreover, Nasua narica showed two peaks of visits, the first one between 15:00 and 16:00 hours and the second between 19:00 and 20:00 hours (Fig. 4), while D. virginiana was the second species that also presented an important pattern of visits to the waterholes; however, this pattern was exclusively nocturnal with a peak between 19:00 and 22:00 hours (Fig. 5).
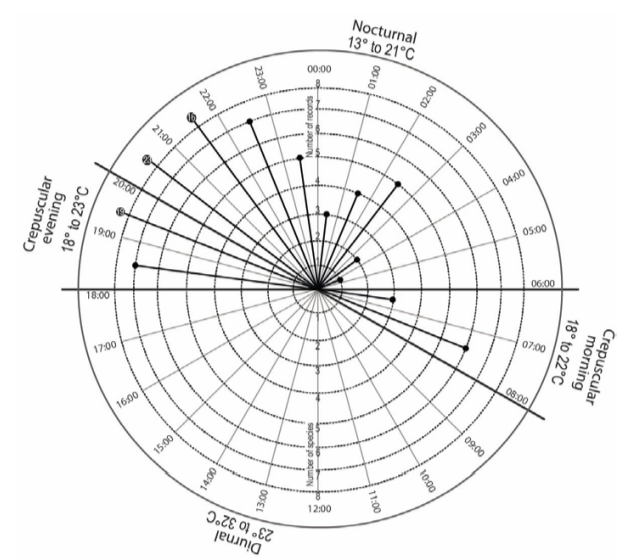
Figure 4 Activity patterns of Nasua narica associated to natural waterholes (“aguajes”) in CNPA El Gavilán, Oaxaca, Mexico.
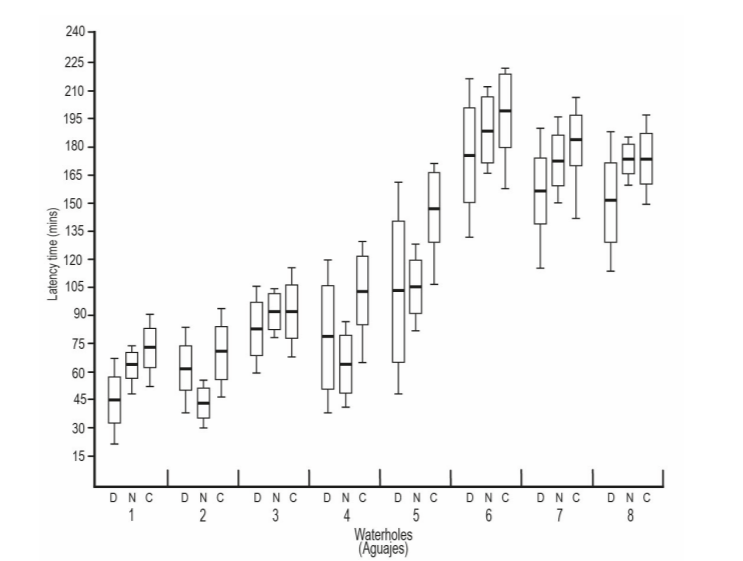
Figure 5 Activity patterns of Didelphis virginiana associated to natural waterholes (“aguajes”) in CNPA El Gavilán, Oaxaca, Mexico.
Regarding the latency time between visitors in NW, Venero Plátano Morado showed the shortest time, while Arroyo Grande and El Patio NW showed times of latency above 150 minutes. The frequency of visitors between all NW varied significantly (F= 6.25, df. = 5.25, P= 0.01; Fig. 6).
Discussion
Natural waterholes (“aguajes”) are important components of habitat, mainly in sites where there is a shortage of surface water and thus become a limiting factor (Dávila et al., 2019). Water availability is a limited resource for several months during the dry season; therefore, it has been hypothesized that the probability of negative interactions between species might increase especially when there are limited resources (Rosenstock et al., 1999; Krausman et al., 2006; Perera-Romero et al., 2021). In previous studies, some negative interactions among species such as kicking, pushing, or chasing when using waterholes have been recorded, but these behaviors have not been recorded in our study (Rosenstock et al., 2004; López-Téllez et al., 2021); however, we must point out that our records consisted exclusively of fixed images (photographs) to make the use of batteries efficient, so we do not have the exact certainty that these behaviors have not occurred. In this case, to corroborate these behaviors, the best is to program the cameras in video format as much as possible by shooting.
As for the species richness associated with NW, this relationship is probably due to the water quality contained in each NW; however, although no such assessment was made in this work, in the northern of the Mexico it has been shown for some species mainly that the pH values in water, chlorine concentrations and the minerals of it (iron, magnesium and aluminum concentrations) may influence the pattern of water consumption by wildlife, so that the waterhole most visited by a larger number of species are those that present the optimal water conditions for drinking and represent lower risk even of predation. Therefore, in a new assessment phase will require the implementation of water quality of the NW, to determine whether these have any influence on the pattern of visits and species associated with these water bodies characteristics during the dry season.
On the other hand, the greatest frequency of visits to water was recorded in April and May, precisely in the months when temperature increased more intensely, thus creating a marked restriction on access to surface water because of water evaporation. The most notorious effect of this situation is water scarcity on these months and the animals losing weight and several of them died (Ramírez Lozano, 2004).
The presence of water determines many aspects of species’ behavior and ecology, also influences the aggregation of species and individuals in time and spaces (Ayeni, 1975; Jiménez-Sánchez et al., 2024; Reyna-Hurtado & Arias-Domínguez, 2024); but this is more pronounced in diurnal birds (Fisher et al., 1972; Dávila et al., 2019). Our findings are like to those obtained by Lynn et al. (2006), and by Dávila et al. (2019), whose recorded diurnal bird’s attendance at NW. Moreover, Columbidae were the most frequent species recorded in these studies. But, in this work besides pigeons and doves (Columbidae), we also recorded to O. poliocephala (Cracidae), both are granivorous and frugivorous species. Probably, NW be relevant for granivorous species than for other birds, due to dry seeds contain a small amount of water and the NW represent a potential key resource for vertebrate species both as of drinking water and as a probable tool for regulating body temperature by bathing (Fisher et al., 1972; Delgado-Martínez et al., 2018).
The main feature of the dry season in tropical dry forests is the increase in temperature and the decrease in the rainfall, the latter determines the phenological patterns and water availability (Castro et al., 2018; Zou et al., 2021). Based on this, most “aguajes” are maintained due to its size and depth, which also affect their thermal and chemical composition. This could explain why three of eight “aguajes” present more vertebrates associated with them.
The schedule when more species records were obtained was daylight, with an average temperature on 28°C, being the birds the group that most used these NW during the day. Although water need varies between groups or species, birds are likely to have high water loss rates, mainly through expired air (Williams, 1996). Therefore, they require a water supply in times with the highest incidence of sunlight that they can obtain through diet, metabolic processes or through direct intake (García-Castellano et al., 2016), as it is in this case.
The NW in CNPA El Gavilán are the easiest surface water sources for water obtaining, unlike water they can obtain from food, metabolic water produced by oxidation of organic nutrients and preformed water associated with body tissues that are catabolized during periods when the energy balance is negative (as in long fasts) (Ramírez-Lozano, 2004). So, our results are like to those reported by Hervert & Krausman (1986), and Hazam & Krausman (1988), who conclude that visitors to NW increase in daytime periods and midday hours. In addition, although the assessment in this work was not extended to the rainy period due to logistical situations, the phenological contrast associated with the lack of rains in the drought period in this region, it highlights the importance of these NW. It has been shown that during the drought period, NW are of extreme importance as sources of water for tropical fauna (Aranda, 1994; Vaughan & Weiss, 1999; Hernández-Huerta et al., 2000; Naranjo, 2001; Biggs et al., 2017).
Although the pattern of frequency of visits by different species to NW has not been widely studied in tropical dry habitats, in some species like the white-tail deer (Odocoileus virginianus), Escobar-Flores & Sandoval (2017) found that deer tend to visit the waterholes in two main hours, from 05:00 to 08:00 am and from 18:00 to 19:00 hours. This activity pattern coincides with what is recorded in this study for the same species. Similarly, our results match to those reported in the Calakmul region, where water sources are critical to meeting the physiological needs of wildlife (Aranda, 1994; Hernández-Huerta et al., 2000; Naranjo, 2001; Jiménez-Sánchez et al., 2024; Reyna-Hurtado & Arias-Domínguez, 2024).
Conclusion
Our study emphasizes basic aspects of the natural history of species through the qualitative observation of photo-captures, which offer us the opportunity to observe the wild vertebrates in activities that are not easy to record. Natural waterholes assessed here contributes to the survival of several species and they serve as sites for hydration, thermoregulation and resting. Given the importance of these NW within the CNPA El Gavilan and under the context of global climate change, our findings are of relevance due to the growing concern regarding the expected pronounced dry seasons in the coast of Oaxaca with an increase of temperatures and a decrease of precipitation in the tropical regions (O’Farrill et al., 2014). Therefore, the protection of these sites is prevailing where the NW (“aguajes”) represent an important water source for wildlife.

