Articles
A pheromone lure for catching fall armyworm males (Lepidoptera: Noctuidae) in Mexico
Una feromona para la captura de machos del gusano cogollero (Lepidoptera: Noctuidae) en México

ABSTRACT
Moth sex pheromones can be used for monitoring or for control (e.g., mass trapping or mating disruption) of agricultural pests. However, pheromone application in field conditions requires that all crucial pheromonal components are included and optimized in a pheromone lure. In this study, we optimized a sex pheromone lure for monitoring Mexican populations from the fall armyworm (FAW), Spodoptera frugiperda Smith. In corn fields, we evaluated the activity of single compounds, three binary blends, and one tertiary blend of (Z)-7-dodecenyl acetate (Z7-12:OAc), (Z)-9-tetradecenyl acetate (Z9-14:OAc), and (Z)-11-hexadecenyl acetate (Z11-16:OAc). The results showed that the traps baited with a binary blend of Z7-12:Ac + Z9-14:Ac captured significantly more S. frugiperda males than single compounds, other binary blends or the tertiary blend. The absence of Z11-16:OAc in the blends did not significantly affect the capture of males, indicating that this compound is non-essential for male attraction in the field. We demonstrated that the dose of the binary blend of Z7-12:OAc and Z9-14:OAc influenced the catch rate of FAW males. The pheromone lure optimized in this study can be useful for capturing S. frugiperda in Mexico.
Key words::
Spodoptera frugiperda, insects, semiochemicals, trapping, geographical variation
INTRODUCTION
The sexual communication system of a many of moth species is mediated by pheromones and may vary geographically among populations (Toth et al., 1992; Gemeno et al., 2000; Groot et al., 2009). This variation includes qualitative or quantitative changes of sexual pheromone components and/or male responses to them (Phelan, 1992; Baker, 2002; Smadja & Butlin, 2009). From a practical point of view, the geographical variation in sexual communication of moths may be important for the management of agricultural pests, by either monitoring or control (i.e., mass trapping, attraction-annihilation, or mating disruption) using synthetic pheromones (Cardé & Minks, 1995; El-Sayed et al., 2009; Witzgall et al., 2010). For instance, a monitoring program could be less successful if it uses pheromone lures formulated from populations distributed in regions different from the target population.
-
Toth et al., 1992Attraction of male turnip moths Agrotis segetum (Lepidoptera: Noctuidae) to sex pheromone components and their mixtures at 11 sites in Europe, Asia, and AfricaJournal of Chemical Ecology, 1992
-
Gemeno et al., 2000Pheromone blend variation and cross-attraction among populations of the black cutworm moth (Lepidoptera: Noctuidae)Annals of the Entomological Society of America, 2000
-
Groot et al., 2009Geographic and temporal variation in moth chemical communicationEvolution: International Journal of Organic Evolution, 2009
-
Phelan, 1992Evolution of sex pheromones and the role of asymmetric trackingInsect Chemical Ecology: An Evolutionary Approach, 1992
-
Baker, 2002Mechanism for saltational shifts in pheromone communication systemsProceedings of the National Academy of Sciences USA, 2002
-
Smadja & Butlin, 2009On the scent of speciation: the chemosensory system and its role in premating isolationHeredity, 2009
-
Cardé & Minks, 1995Control of moth pests by mating disruption: successes and constraintsAnnual Review of Entomology, 1995
-
El-Sayed et al., 2009Potential of “lure and kill” in long-term pest management and eradication of invasive speciesJournal of Economic Entomology, 2009
-
Witzgall et al., 2010Sex pheromones and their impact on pest managementJournal of Chemical Ecology, 2010
The fall armyworm (FAW), Spodoptera frugiperda (J. E. Smith), is the most important insect pest of corn crops in Mexico (Blanco et al., 2014), but it is also widely distributed from north of the USA to north of Argentina (Prowell et al., 2004; Hardke et al., 2011; Pashley, 1986). However, recently, FAW has become a harmful, invasive pest of corn in sub-Saharan Africa (Goergen et al., 2016; Stokstad, 2017) and Asia (Wu et al., 2019; Padhee & Prasanna, 2019).
-
Blanco et al., 2014Maize pests in Mexico and challenges for the adoption of integrated pest management programsJournal of Integrated Pest Management, 2014
-
Prowell et al., 2004Multilocus genetic analysis of host use, introgression, and speciation in host strains of fall armyworm (Lepidoptera: Noctuidae)Annals of the Entomological Society of America, 2004
-
Hardke et al., 2011Damage and survivorship of fall armyworm (Lepidoptera: Noctuidae) on transgenic field corn expressing Bacillus thuringiensis Cry proteinsCrop Protection, 2011
-
Pashley, 1986Host-associated genetic differentiation in fall armyworm (Lepidoptera: Noctuidae): a sibling species complex?Annals of the Entomological Society of America, 1986
-
Goergen et al., 2016First report of outbreaks of the fall armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central AfricaPloS One, 2016
-
Stokstad, 2017New crop pest takes Africa at lightning speedScience, 2017
-
Wu et al., 2019Estimation of the potential infestation area of newly-invaded fall armyworm Spodoptera frugiperda in the yangtze river valley of ChinaInsects, 2019
-
Padhee & Prasanna, 2019The emerging threat of fall armyworm in IndiaIndian Farming, 2019
Spodoptera frugiperda females expose the last abdominal segments for releasing their sex pheromone components into the environment during calling behavior (Cruz-Esteban et al., 2017). The effluvia and gland pheromone extracts from calling FAW females contain several acetates, including (Z)-9-tetradecenyl acetate (Z9-14:OAc) as a major component, (Z)-7-dodecenyl acetate (Z7-12:OAc), (Z)-11-hexadecenyl acetate (Z11-16:OAc), (Z)-9-dodecenyl acetate (Z9-12:OAc), and (E)-7-dodecenyl acetate (E7-12:OAc) (Mitchell et al., 1985; Tumlinson et al., 1986; Descoins et al., 1988; Groot et al., 2008; Lima & McNeil, 2009; Unbehend et al., 2013; Batista-Pereira et al., 2006). The latter compound has only been reported being produced by Brazilian FAW populations (Batista-Pereira et al., 2006).
-
Cruz-Esteban et al., 2017Calling behavior, copulation time, and reproductive compatibility of corn-strain fall armyworm (Lepidoptera: Noctuidae) from populations in MexicoEnvironmental Entomology, 2017
-
Mitchell et al., 1985Field evaluation of commercial pheromone formulations and traps using a more effective sex pheromone blend for the fall armyworm (Lepidoptera: Noctuidae)Journal of Economic Entomology, 1985
-
Tumlinson et al., 1986Sex pheromone of fall armyworm, Spodoptera frugiperda (JE Smith)Journal of Chemical Ecology, 1986
-
Descoins et al., 1988Monitoring of crop pests by sexual trapping of males in Guadeloupe and GuyanaAgriculture, Ecosystems & Environment, 1988
-
Groot et al., 2008Host strain specific sex pheromone variation in Spodoptera frugiperdaFrontiers in Zoology, 2008
-
Lima & McNeil, 2009Female sex pheromones in the host races and hybrids of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae)Chemoecology, 2009
-
Unbehend et al., 2013Pheromonal divergence between two strains of Spodoptera frugiperdaJournal of Chemical Ecology, 2013
-
Batista-Pereira et al., 2006Isolation, identification, synthesis, and field evaluation of the sex pheromone of the Brazilian population of Spodoptera frugiperdaJournal of Chemical Ecology, 2006
-
Batista-Pereira et al., 2006Isolation, identification, synthesis, and field evaluation of the sex pheromone of the Brazilian population of Spodoptera frugiperdaJournal of Chemical Ecology, 2006
Commercial FAW sex pheromone lures formulated from USA populations are used for monitoring this agricultural pest in several countries. However, the capture of FAW males by traps baited with these commercial lures are erratic when these were evaluated in Mexico, Central America and South America (Andrade et al., 2000; Malo et al., 2001; Batista-Pereira et al., 2006), suggesting that pheromonal communication of this moth species may vary geographically. Generally, the commercial lures of FAW are formulated with three pheromone components: Z9-14:OAc, Z11-16:OAc, and Z7-12:OAc (Malo et al., 2001; Bratovich et al., 2019). In Mexico, FAW females collected from corn consistently emitted Z9-14: OAc (65-90%), Z11-16: OAc (4-34%), and Z7-12: OAc (traces-5%) (Cruz-Esteban et al., 2018).
-
Andrade et al., 2000Optimization of a pheromone lure for Spodoptera frugiperda (Smith) in Central AmericaJournal of the Brazilian Chemical Society, 2000
-
Malo et al., 2001Evaluation of commercial pheromone lures and traps for monitoring male fall armyworm (Lepidoptera: Noctuidae) in the coastal region of Chiapas, MexicoFlorida Entomologist, 2001
-
Batista-Pereira et al., 2006Isolation, identification, synthesis, and field evaluation of the sex pheromone of the Brazilian population of Spodoptera frugiperdaJournal of Chemical Ecology, 2006
-
Malo et al., 2001Evaluation of commercial pheromone lures and traps for monitoring male fall armyworm (Lepidoptera: Noctuidae) in the coastal region of Chiapas, MexicoFlorida Entomologist, 2001
-
Bratovich et al., 2019Evaluation of sex pheromone formulations to attract Spodoptera frugiperda (Lepidoptera: Noctuidae) adult males in ArgentinaRevista de la Sociedad Entomológica Argentina, 2019
-
Cruz-Esteban et al., 2018Geographic variation in pheromone component ratio and antennal responses, but not in attraction, to sex pheromones among fall armyworm populations infesting corn in MexicoJournal of Pest Science, 2018
In this work, we investigated whether all three components emitted by FAW females from Mexican populations are crucial for attracting conspecific males in field conditions. The objective of this work was the optimization of a sex pheromone lure that would be useful for monitoring S. frugiperda in corn crops in Mexico.
MATERIALS AND METHODS
Chemicals. Synthetic Z7-12:OAc, Z9-14:OAc and Z11-16:OAc were purchased from commercial sources (Sigma-Aldrich, Toluca, Mexico). The purity of synthetic compounds (<97%) was determined with a Shimadzu GC-2010 plus gas chromatograph (Tokyo, Japan) equipped with a flame ionization detector. We prepared standard solutions of the compounds in (HPLC)-grade hexane (Sigma-Aldrich, Toluca, Mexico).
Field tests. In the first experiment, we evaluated the attractiveness of single compounds or the binary and tertiary blends of synthetic pheromone compounds formulated according to the relative proportion of each compound as released by FAW females from Tapachula, Chiapas (Cruz-Esteban et al., 2018). The compound (Z7-12:OAc) that was found at trace levels was assigned to 1% of the blend. The experiment was performed in the Ejido Joaquín Miguel Gutiérrez (14° 44' N, 92° 19' W), municipality of Tapachula, Chiapas, Mexico, in March 2017. This experiment was deployed in a fully randomized block design with four replicates per treatment. In each block, nine yellow plastic jug traps (Malo et al., 2018) were used for one of each treatment (see below). The blocks were arranged in parallel lines separated ca. 50 m apart in a corn crop (approximately 8 ha). Rubber septa lures containing 400 µg of each synthetic compound (Z9-14:OAc, Z7-12:OAc or Z11-16:OAc) or the binary and tertiary blends were placed in the traps. A virgin female of four-five day old, reared in the laboratory, was used as a positive control, and a non-baited trap was used as a negative control. The traps with the lures were hung at a height of 1.5 m above the ground on wooden stakes placed 50 m apart from each another.
-
Cruz-Esteban et al., 2018Geographic variation in pheromone component ratio and antennal responses, but not in attraction, to sex pheromones among fall armyworm populations infesting corn in MexicoJournal of Pest Science, 2018
-
Malo et al., 2018A home-made trap baited with sex pheromone for monitoring Spodoptera frugiperda males (Lepidoptera: Noctuidae) in corn crops in MexicoJournal of Economic Entomology, 2018
In the second experiment, we tested the effect of the proportion of the most attractive blend obtained in the first experiment. This experiment was performed in the Ejido Joaquín Miguel Gutiérrez in June 2017. The experiment was deployed in a fully randomized block design. Yellow plastic jug traps were placed and distributed as in the first experiment. Lures consisted of rubber septa loaded with 400 µg of Z7-12:OAc + Z9-14:OAc at 99:1, 1:99, 50:50, 89:11, or 11:89 proportion.
In the third experiment, we evaluated the effect of three proportions (1:99, 1.5:98.5, and 2:98) of the binary blend Z7-12:OAc + Z9-14:OAc for catching FAW males. Three trials of this experiment were performed, one in the Ejido Joaquín Miguel Gutiérrez, another in San Nicolás Lagartero (14° 49' N, 92° 15' W), Tapachula, Chiapas, and Ciudad Guzmán (19° 42' N, 103° 28' W), Jalisco. The distance between the Ejido Joaquín Miguel Gutiérrez and San Nicolás Lagartero was greater than 10 km. The first two trials were performed during August-September 2017. The distribution of the yellow plastic jug traps was like the first experiment.
In the fourth experiment, we tested the effect of the dose of the binary blend of Z7-12:OAc + Z9-14:OAc on the capture of FAW males. The experiment was performed in the Ejido Joaquín Miguel Gutiérrez in October-November 2017. The experiment was deployed in a fully randomized block design. The distribution of the yellow plastic jug traps was like the first experiment. Rubber septa were loaded with 200, 400, 600, or 1600 µg of Z7-12:OAc + Z9-14:OAc (99:1).
In all experiments, the traps were checked every 3 days. On each observation date, traps were emptied, and the number of males caught was recorded. Treatments were rotated within each block, and all experiment lures were changed every 15 days and virgin females were exchanged daily for another of the same age.
Data analysis. Data were analyzed with statistical software R version 3.6.1 (R Development Core Team, 2019). The number of moths captured/trap/night was analyzed by repeated measures analysis of variance (ANOVA). Prior to analysis, we checked that data meet the ANOVA´s assumptions, otherwise they were transformed according to the Box-Cox method. Treatment means were compared with a Tukey test (α = 0.05).
-
R Development Core Team,
2019R: A language and environment for statistical computing, 2019
RESULTS
In the first experiment, we found that the treatments affected the captures of FAW males (F = 17.6; df = 8, 135; P < 0.001). Traps baited with the binary blend of Z9-14:OAc + Z7-12:OAc caught significantly more FAW males than traps baited with other treatments (Fig. 1). Traps baited with Z7-12:OAc caught a similar number of males as traps baited with females. There were no differences in the number of FAW males captured by traps baited with Z9-14:OAc alone, Z11-16:OAc alone, Z7-12:OAc + Z11-16:OAc, Z9-14:OAc + Z11-16:OAc, Z7-12:OAc + Z9-14:OAc + Z11-16:OAc, and non-baited traps (Fig. 1).
Thumbnail

Figure 1
Mean number (± S. E.) of males caught by traps baited with single compounds, binary, and tertiary blend in corn cultivations. The binary and tertiary blends were prepared according was found in the effluvia of females from Tapachula, Chiapas. The numbers related to each compound are the percentages used to prepare the blend, and the different letters above column indicate significant differences among treatments (α = 0.05).
Mean number (± S. E.) of males caught by traps baited with single compounds, binary, and tertiary blend in corn cultivations. The binary and tertiary blends were prepared according was found in the effluvia of females from Tapachula, Chiapas. The numbers related to each compound are the percentages used to prepare the blend, and the different letters above column indicate significant differences among treatments (α = 0.05).
In the second experiment, our results showed that the proportion of the Z9-14: OAc+Z7-12:OAc blend affected the capture of FAW males (F = 16.6; df = 4, 95; P < 0.0001). Traps baited with the proportion 99:1 of this binary blend captured more males in comparison to the other proportions tested (Fig. 2).
Thumbnail
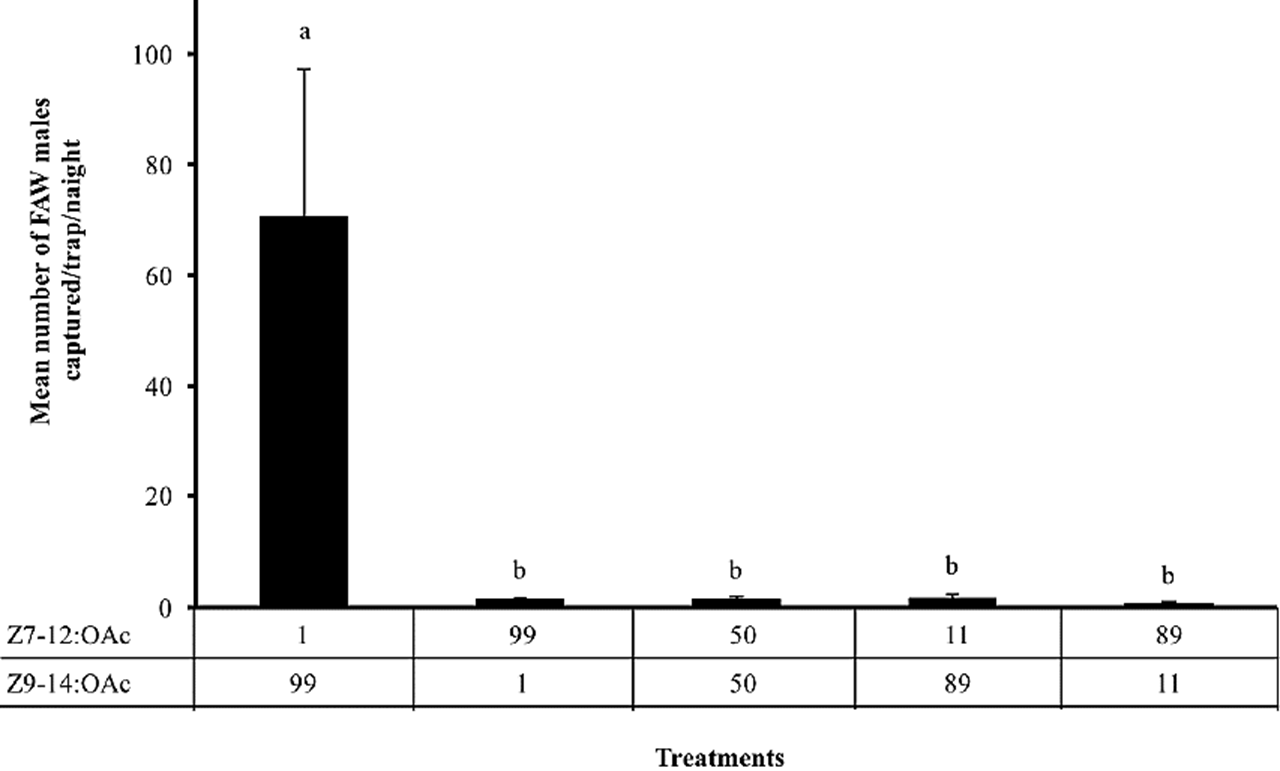
Figure 2
Mean number (± S. E.) of males caught by traps baited with blends of different proportion of the synthetic components (Z7-12:OAc + Z9-14:OAc) tested in corn cultivations. The numbers related to each compound are the percentages used to prepare the blend, and different letters above column indicate significant differences among treatments (α = 0.05).
Mean number (± S. E.) of males caught by traps baited with blends of different proportion of the synthetic components (Z7-12:OAc + Z9-14:OAc) tested in corn cultivations. The numbers related to each compound are the percentages used to prepare the blend, and different letters above column indicate significant differences among treatments (α = 0.05).
In the third experiment, we found that there was no effect of the three proportions of the binary blend Z9-14:OAc + Z7-12:OAc tested on the FAW male captures in the Ejido Joaquín Miguel Gutiérrez (F = 1.8; df = 2, 126; P = 0.71) and Ejido San Nicolás Lagartero (F = 1.52; df = 2, 117; P = 0.22), Tapachula, Chiapas (Fig. 3A, 3B), but there was a significant effect at Ciudad Guzman (F = 11.64; df = 2,105; P < 0.0001), Jalisco (Fig. 3C). In the last location (Jalisco), traps baited with the proportion 99:1 (Z9-14:OAc and Z7-12:OAc, respectively) captured more FAW males than traps baited with the proportions of 98.5:1.5 and 98:2 (Fig. 3C). In the two experiments that were carried out in Tapachula, Chiapas, the proportion of 99:1 (Z9-14:OAc and Z7-12:OAc, respectively) attracted numerically the largest number of males (Fig. 3A, 3B).
Thumbnail
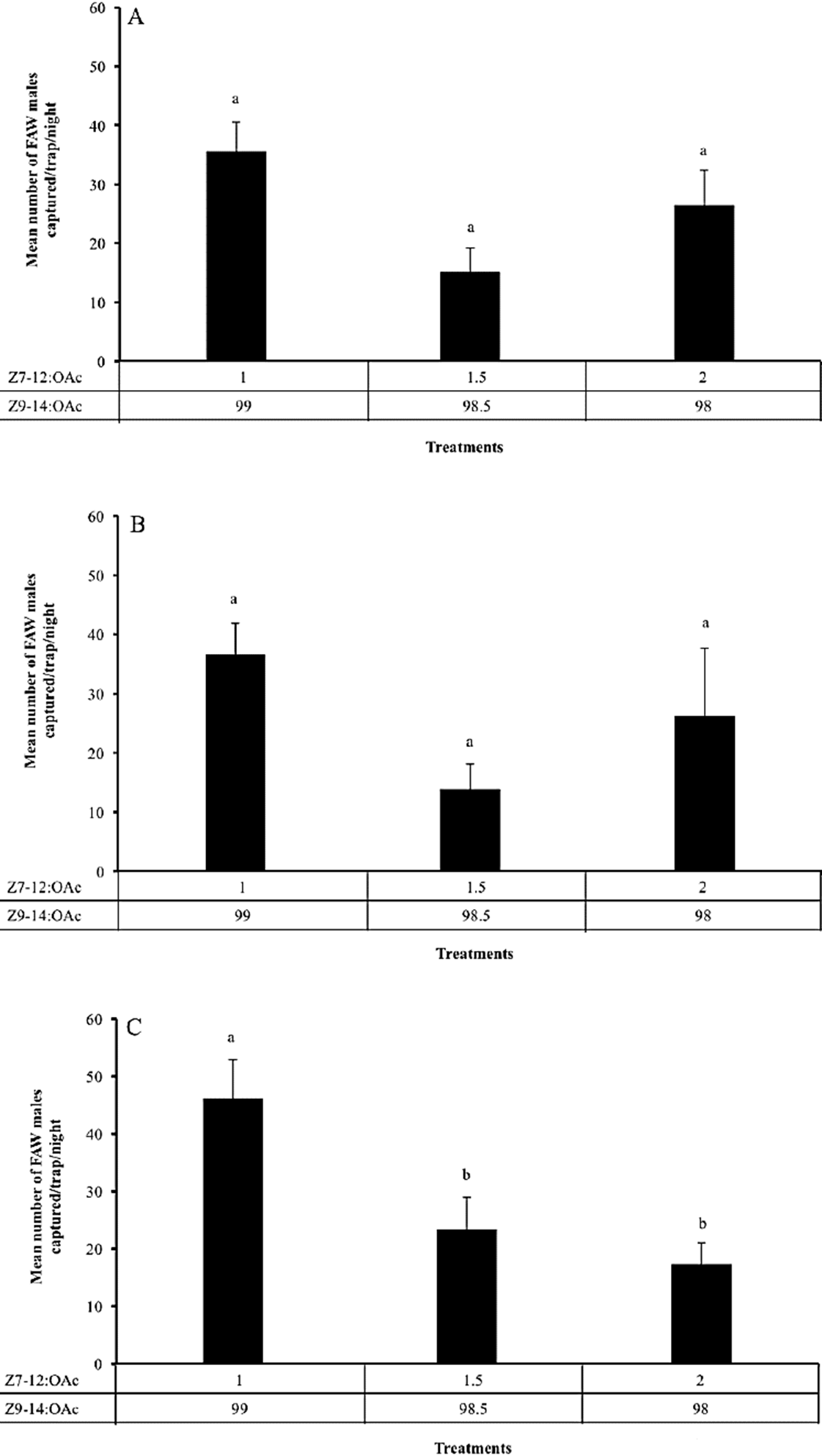
Figure 3
Mean number (± S. E.) of males caught by traps baited with the binary blend (Z7-12:OAc + Z9-14:OAc) in different proportion in corn cultivations. (A) Joaquín Miguel Gutiérrez, Tapachula Chiapas, Mexico. (B) San Nicolás Lagartero, Tapachula Chiapas, Mexico. (C) Ciudad Guzmán, Jalisco, Mexico. The numbers related to each compound are the percentages used to prepare the blend, and the letters above column indicate significant differences among treatments (α = 0.05).
Mean number (± S. E.) of males caught by traps baited with the binary blend (Z7-12:OAc + Z9-14:OAc) in different proportion in corn cultivations. (A) Joaquín Miguel Gutiérrez, Tapachula Chiapas, Mexico. (B) San Nicolás Lagartero, Tapachula Chiapas, Mexico. (C) Ciudad Guzmán, Jalisco, Mexico. The numbers related to each compound are the percentages used to prepare the blend, and the letters above column indicate significant differences among treatments (α = 0.05).
In the fourth experiment, our results showed that the dose of the blend constituted by the Z9-14: OAc + Z7-12:OAc blend affected the capture of FAW males (F = 14.47; df = 4, 5; P < 0.001). Traps baited with 600 µg of Z9-14: OAc + Z7-12:OAc captured more males than traps baited with 200 µg of this blend or non-baited traps (Fig. 4). The captures of traps baited with 600 µg of the blend were not different than those baited with 400 and 1600 µg of the blend. There were no significant differences in the captures of traps baited with 200 µg of the blend and those traps baited with 400 and 1600 µg of the blend (Fig. 4).
Thumbnail
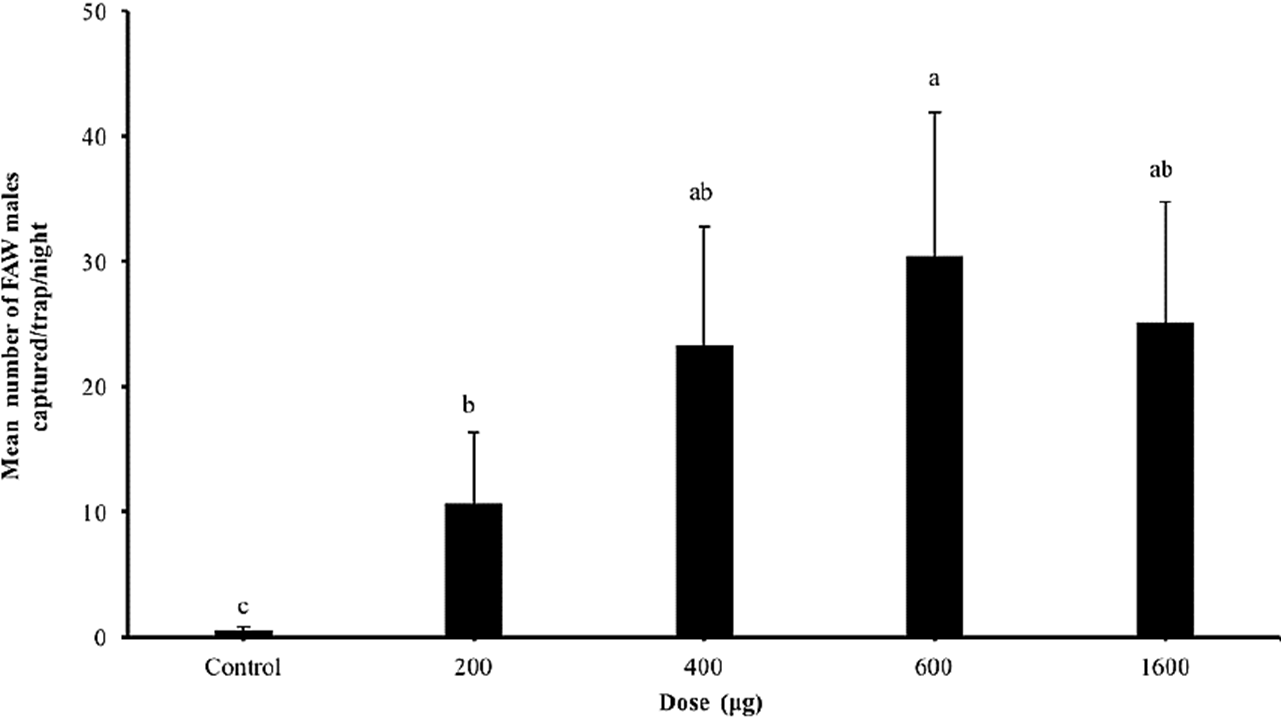
Figure 4
Mean number (( S. E.) of males caught by traps baited with different doses (µg) of the binary blend of Z9-14:OAc (99%) + Z7-12:OAc (1%) tested in the corn cultivations. The numbers related to each compound are the percentages used to prepare the blend, and different letters above column indicate significant differences among treatments (α = 0.05).
Mean number (( S. E.) of males caught by traps baited with different doses (µg) of the binary blend of Z9-14:OAc (99%) + Z7-12:OAc (1%) tested in the corn cultivations. The numbers related to each compound are the percentages used to prepare the blend, and different letters above column indicate significant differences among treatments (α = 0.05).
DISCUSSION
In this study, we optimized a synthetic sexual pheromone for monitoring S. frugiperda males in Mexico. Our results demonstrated that the binary blend of Z9-14:OAc + Z7-12:OAc captured more males. Previously, we found that Z11-16:OAc elicited lower antennal responses in comparison to those evoked by Z9-14:OAc and Z7-12:OAc (Cruz-Esteban et al., 2018). Thus, the male antennal responses matched with the attraction of FAW males to both compounds. Our results also confirm that both acetates are critical components of the sex pheromones of FAW, as reported previously for other studies performed in several regions of the North American continent (Tumlinson et al., 1986; Andrade et al., 2000; Fleischer et al., 2005; Unbehend et al., 2014), but not in Brazil. In the latter country, in addition to Z7-12:OAc and Z9-14:OAc, E12:OAc is a third active component in the sex pheromone of the Brazilian populations of S. frugiperda (Batista-Pereira et al., 2006).
-
Cruz-Esteban et al., 2018Geographic variation in pheromone component ratio and antennal responses, but not in attraction, to sex pheromones among fall armyworm populations infesting corn in MexicoJournal of Pest Science, 2018
-
Tumlinson et al., 1986Sex pheromone of fall armyworm, Spodoptera frugiperda (JE Smith)Journal of Chemical Ecology, 1986
-
Andrade et al., 2000Optimization of a pheromone lure for Spodoptera frugiperda (Smith) in Central AmericaJournal of the Brazilian Chemical Society, 2000
-
Fleischer et al., 2005Spodoptera frugiperda pheromone lures to avoid nontarget captures of Leucania phragmatidicolaJournal of Economic Entomology, 2005
-
Unbehend et al., 2014Geographic variation in sexual attraction of Spodoptera frugiperda corn-and rice-strain males to pheromone luresPloS One, 2014
-
Batista-Pereira et al., 2006Isolation, identification, synthesis, and field evaluation of the sex pheromone of the Brazilian population of Spodoptera frugiperdaJournal of Chemical Ecology, 2006
We found that a change in the proportions of Z7-12:OAc and Z14:OAc did not affect the capture of FAW males in Chiapas, but it did in Jalisco. This result could suggest that there is a geographical variation in the sexual communication in the Mexican populations of S. frugiperda. However, in a previous study, we found that while the proportion of pheromone components emitted by females and the male antennal responses to the compounds varied among regions, males were not differentially caught by traps baited with different proportions of Z7-12:OAc, Z9-14:OAc and Z11-16:OAc (Cruz-Esteban et al., 2018). Thus, other factors such as habitat composition or population density of FAW may affect the capture of FAW males in Chiapas and Jalisco. For example, traps were placed in different corn varieties from different phenological states. Environmental factors were also different in the experimental sites. Thus, background odors could have influenced the response of the males to their pheromone. For instance, other studies have documented that capture of FAW males of the corn race differ in traps baited with the same pheromone blend placed in sorghum, corn, rice, sugarcane, turfgrass or vegetables located in North and South Florida, USA (Unbehend et al., 2013; Meagher & Nagoshi, 2013). Previously, we reported that host volatiles elicited antennal responses in both sexes of S. frugiperda (Malo et al., 2004). Further studies will be performed to investigate the role of host volatiles in the male response to sex pheromones.
-
Cruz-Esteban et al., 2018Geographic variation in pheromone component ratio and antennal responses, but not in attraction, to sex pheromones among fall armyworm populations infesting corn in MexicoJournal of Pest Science, 2018
-
Unbehend et al., 2013Pheromonal divergence between two strains of Spodoptera frugiperdaJournal of Chemical Ecology, 2013
-
Meagher & Nagoshi, 2013Attraction of fall armyworm males (Lepidoptera: Noctuidae) to host strain femalesEnvironmental Entomology, 2013
-
Malo et al., 2004Antennal sensilla and electrophysiological response of male and female Spodoptera frugiperda (Lepidoptera: Noctuidae) to conspecific sex pheromone and plant odorsAnnals of the Entomological Society of America, 2004
At first glance, it seems that the addition of Z11-16:OAc to the binary blend of Z9-14:OAc + Z7-12:OAc interrupted the capture of FAW males. This result could also explain the erratic results of male capture by commercially available sex pheromones of S. frugiperda, which, generally, contain Z11-16:OAc (Malo et al., 2001; Bratovich et al., 2019). However, Z11-16:OAc did not affect the capture of FAW males when added to Z7-12:OAc and Z14:OAc in similar trials in Florida, USA, Brazil, and Argentina (Tumlinson et al., 1986; Batista-Pereira et al., 2006). A possibility to explain this contradiction is that we used a higher percentage of Z11-16:OAc in the tertiary blend compared to previous studies. However, Cruz-Esteban et al. (2018) found that a similar percentage of Z11-16:OAc in the tertiary blend did not affect the males caught in the same region that we performed the present study. Unfortunately, the binary blend of Z7-12:OAc and Z9-14:OAc was not included in the study of Cruz-Esteban et al. (2018). The compound Z11-16:OAc was found in the effluvia of calling females from 22 regions of Mexico (Cruz-Esteban et al., 2018). It will be necessary to perform further experiments to investigate if this compound has an ecological role in the sexual communication of S. frugiperda or if it is only an unavoidable byproduct of the pheromone biosynthetic pathway of this moth species (Groot et al., 2008).
-
Malo et al., 2001Evaluation of commercial pheromone lures and traps for monitoring male fall armyworm (Lepidoptera: Noctuidae) in the coastal region of Chiapas, MexicoFlorida Entomologist, 2001
-
Bratovich et al., 2019Evaluation of sex pheromone formulations to attract Spodoptera frugiperda (Lepidoptera: Noctuidae) adult males in ArgentinaRevista de la Sociedad Entomológica Argentina, 2019
-
Tumlinson et al., 1986Sex pheromone of fall armyworm, Spodoptera frugiperda (JE Smith)Journal of Chemical Ecology, 1986
-
Batista-Pereira et al., 2006Isolation, identification, synthesis, and field evaluation of the sex pheromone of the Brazilian population of Spodoptera frugiperdaJournal of Chemical Ecology, 2006
-
Cruz-Esteban et al. (2018)Geographic variation in pheromone component ratio and antennal responses, but not in attraction, to sex pheromones among fall armyworm populations infesting corn in MexicoJournal of Pest Science, 2018
-
Cruz-Esteban et al. (2018)Geographic variation in pheromone component ratio and antennal responses, but not in attraction, to sex pheromones among fall armyworm populations infesting corn in MexicoJournal of Pest Science, 2018
-
Cruz-Esteban et al., 2018Geographic variation in pheromone component ratio and antennal responses, but not in attraction, to sex pheromones among fall armyworm populations infesting corn in MexicoJournal of Pest Science, 2018
-
Groot et al., 2008Host strain specific sex pheromone variation in Spodoptera frugiperdaFrontiers in Zoology, 2008
Our results suggest that a dose of 400 or 600 (g can be used for monitoring S. frugiperda males. In contrast, other authors used 2000 (g of the sex pheromone blend (Tumlinson et al., 1986; Andrade et al., 2000). However, our results were similar to those reported by Unbehend et al. (2014), who found that a blend of 300 (g of Z9-14:OAc + 6 (g Z7-12:OAc was the most effective to capture males of S. frugiperda in corn fields in several countries in North and South America. In Mexico, only Z9-14:OAc and Z7-12:OAc are needed to capture S. frugiperda males in corn and sorghum fields, it differs to other regions where it needs the addition of Z11-16:OAc and Z9-12:OAc (Malo et al., 2001; Bratovich et al., 2019). Future studies will be necessary to test the commercial sex pheromones of S. frugiperda to find the most attractive for geographical region.
-
Tumlinson et al., 1986Sex pheromone of fall armyworm, Spodoptera frugiperda (JE Smith)Journal of Chemical Ecology, 1986
-
Andrade et al., 2000Optimization of a pheromone lure for Spodoptera frugiperda (Smith) in Central AmericaJournal of the Brazilian Chemical Society, 2000
-
Unbehend et al. (2014)Geographic variation in sexual attraction of Spodoptera frugiperda corn-and rice-strain males to pheromone luresPloS One, 2014
-
Malo et al., 2001Evaluation of commercial pheromone lures and traps for monitoring male fall armyworm (Lepidoptera: Noctuidae) in the coastal region of Chiapas, MexicoFlorida Entomologist, 2001
-
Bratovich et al., 2019Evaluation of sex pheromone formulations to attract Spodoptera frugiperda (Lepidoptera: Noctuidae) adult males in ArgentinaRevista de la Sociedad Entomológica Argentina, 2019
In summary, we found that traps baited with binary blends of pheromones Z9-14:OAc (99%) and Z7-12:OAc (1%) caught significantly more S. frugiperda males in corn crops compared to other binary blends, a tertiary blend of Z7-12:OAc, Z9-14:OAc, and Z11-16:OAC, and a natural pheromone (e.g., females). This result suggests that this blend may be useful for monitoring FAW males in Mexico. Future studies will be performed to test the binary blend of the sex pheromone using a mass trapping to control S. frugiperda in corn crops in Mexico.
Acknowledgements
We thank Armando Virgen Sanchez, Neby M. Méridia Torres, and Álvaro José Campuzano Granados for their support in field tests, and Ferosex S.A. de C.V. for the logistic support during the trial performed in Jalisco. SCE (CVU: 316166) acknowledges receipt of a doctoral scholarship from the Mexican National Council for Science and Technology (CONACyT).
LITERATURE CITED
- Andrade, R., Rodriguez, C., Oehlschlager, A. C. (2000) Optimization of a pheromone lure for Spodoptera frugiperda (Smith) in Central America. Journal of the Brazilian Chemical Society, 11 (6), 609-613. http://dx.doi.org/10.1590/S0103-50532000000600009 Links
- Baker, T. C. (2002) Mechanism for saltational shifts in pheromone communication systems. Proceedings of the National Academy of Sciences USA, 99 (21), 13368-13370. https://doi.org/10.1073/pnas.222539799 Links
- Batista-Pereira, L. G., Stein, K., de Paula, A. F., Moreira, J. A., Cruz, I., Figueiredo, M. L. C., Perri, J. Jr., Corrêa, A. G. (2006) Isolation, identification, synthesis, and field evaluation of the sex pheromone of the Brazilian population of Spodoptera frugiperda. Journal of Chemical Ecology, 32 (5), 1085. https://doi.org/10.1007/s10886-006-9048-5 Links
- Blanco, C. A., Pellegaud, J. G., Nava-Camberos, U., Lugo-Barrera, D., Vega-Aquino, P., Coello, J., Terán-Vargas, A. P., Vargas-Camplis, J. (2014) Maize pests in Mexico and challenges for the adoption of integrated pest management programs. Journal of Integrated Pest Management, 5 (4), E1-E9. https://doi.org/10.1603/IPM14006 Links
- Bratovich, C., Saluso, A., Murua, M. G., Guerenstein, P. G. (2019) Evaluation of sex pheromone formulations to attract Spodoptera frugiperda (Lepidoptera: Noctuidae) adult males in Argentina. Revista de la Sociedad Entomológica Argentina, 78 (3), 7-14. Links
- Cardé, R. T., Minks, A. K. (1995) Control of moth pests by mating disruption: successes and constraints. Annual Review of Entomology, 40 (1), 559-585. https://doi.org/10.1146/annurev.en.40.010195.003015 Links
- Cruz-Esteban, S., Rojas, J. C., Malo, E. A. (2017) Calling behavior, copulation time, and reproductive compatibility of corn-strain fall armyworm (Lepidoptera: Noctuidae) from populations in Mexico. Environmental Entomology, 46 (4), 901-906. https://doi.org/10.1093/ee/nvx120 Links
- Cruz-Esteban, S., Rojas, J. C., Sánchez-Guillén, D., Cruz-López, L., Malo, E. A. (2018) Geographic variation in pheromone component ratio and antennal responses, but not in attraction, to sex pheromones among fall armyworm populations infesting corn in Mexico. Journal of Pest Science, 91 (3), 973-983. https://doi.org/10.1007/s10340-018-0967-z Links
- Descoins, C., Silvain, J. F., Lalanne-Cassou, B., Cheron, H. (1988) Monitoring of crop pests by sexual trapping of males in Guadeloupe and Guyana. Agriculture, Ecosystems & Environment, 21 (1-2), 53-56. https://doi.org/10.1016/0167-8809(88)90139-9 Links
- El-Sayed, A. M., Suckling, D. M., Byers, J. A., Jang, E. B., Wearing, C. H. (2009) Potential of “lure and kill” in long-term pest management and eradication of invasive species. Journal of Economic Entomology, 102 (3), 815-835. https://doi.org/10.1603/029.102.0301 Links
- Fleischer, S. J., Harding, C. L., Blom, P. E., White, J., Grehan, J. (2005) Spodoptera frugiperda pheromone lures to avoid nontarget captures of Leucania phragmatidicola. Journal of Economic Entomology, 98 (1), 66-71. https://doi.org/10.1093/jee/98.1.66 Links
- Gemeno, C., Lutfallah, A. F., Haynes, K. F. (2000) Pheromone blend variation and cross-attraction among populations of the black cutworm moth (Lepidoptera: Noctuidae). Annals of the Entomological Society of America, 93 (6), 1322-1328. https://doi.org/10.1603/0013-8746(2000)093[1322:PBVACA]2.0.CO;2 Links
- Goergen, G., Kumar, P. L., Sankung, S. B., Togola, A., Tamò, M. (2016) First report of outbreaks of the fall armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PloS One, 11 (10), e0165632. https://doi.org/10.1371/journal.pone.0165632 Links
- Groot, A. T., Marr, M., Schöfl, G., Lorenz, S., Svatos, A., Heckel, D. G. (2008) Host strain specific sex pheromone variation in Spodoptera frugiperda. Frontiers in Zoology, 5 (1), 20. https://doi.org/10.1186/1742-9994-5-20 Links
- Groot, A. T., Inglis, O., Bowdridge, S., Santangelo, R. G., Blanco, C., López, Jr., Terán-Vargas, A., Fred, G., Schal, C. (2009) Geographic and temporal variation in moth chemical communication. Evolution: International Journal of Organic Evolution, 63 (8), 1987-2003. https://doi.org/10.1111/j.1558-5646.2009.00702.x Links
- Hardke, J. T., Leonard, B. R., Huang, F., Jackson, R. E. (2011) Damage and survivorship of fall armyworm (Lepidoptera: Noctuidae) on transgenic field corn expressing Bacillus thuringiensis Cry proteins. Crop Protection, 30 (2), 168-172. https://doi.org/10.1016/j.cropro.2010.10.005 Links
- Lima, E. R., McNeil, J. N. (2009) Female sex pheromones in the host races and hybrids of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Chemoecology, 19 (1), 29-36. https://doi.org/10.1007/s00049-009-0005-y Links
- Malo, E. A., Cruz-Lopez, L., Valle-Mora, J., Virgen, A., Sanchez, J. A., Rojas, J. C. (2001) Evaluation of commercial pheromone lures and traps for monitoring male fall armyworm (Lepidoptera: Noctuidae) in the coastal region of Chiapas, Mexico. Florida Entomologist, 84 (4), 659-664. Links
- Malo, E. A., Castrejón-Gómez, V. R., Cruz-López, L., Rojas, J. C. (2004) Antennal sensilla and electrophysiological response of male and female Spodoptera frugiperda (Lepidoptera: Noctuidae) to conspecific sex pheromone and plant odors. Annals of the Entomological Society of America , 97 (6), 1273-1284. https://doi.org/10.1603/0013-8746(2004)097[1273:ASAERO]2.0.CO;2 Links
- Malo, E. A., Cruz-Esteban, S., González, F. J., Rojas, J. C. (2018) A home-made trap baited with sex pheromone for monitoring Spodoptera frugiperda males (Lepidoptera: Noctuidae) in corn crops in Mexico. Journal of Economic Entomology , 111 (4), 1674-1681. https://doi.org/10.1093/jee/toy128 Links
- Meagher, R. L., Nagoshi, R. N. (2013) Attraction of fall armyworm males (Lepidoptera: Noctuidae) to host strain females. Environmental Entomology , 42 (4), 751-757. https://doi.org/10.1603/EN13007 Links
- Mitchell, E. R., Tumlinson, J. H., McNeil, J. N. (1985) Field evaluation of commercial pheromone formulations and traps using a more effective sex pheromone blend for the fall armyworm (Lepidoptera: Noctuidae). Journal of Economic Entomology , 78 (6), 1364-1369. https://doi.org/10.1093/jee/78.6.1364 Links
- Padhee, A. K., Prasanna, B. M. (2019) The emerging threat of fall armyworm in India. Indian Farming, 69 (1), 51-54. Links
- ´Pashley, D. P. (1986) Host-associated genetic differentiation in fall armyworm (Lepidoptera: Noctuidae): a sibling species complex? Annals of the Entomological Society of America , 79 (6), 898-904. https://doi.org/10.1093/aesa/79.6.898 Links
- Phelan, P. L. (1992) Evolution of sex pheromones and the role of asymmetric tracking. Insect Chemical Ecology: An Evolutionary Approach, 265-314. Kluwer, Netherlands. Links
- Prowell, D. P., McMichael, M., Silvain, J. F. (2004) Multilocus genetic analysis of host use, introgression, and speciation in host strains of fall armyworm (Lepidoptera: Noctuidae). Annals of the Entomological Society of America , 97 (5), 1034-1044. https://doi.org/10.1603/0013-8746(2004)097[1034:MGAOHU]2.0.CO;2 Links
- R Development Core Team (2019) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: http://www.R-project.org/ Links
- Smadja, C., Butlin, R. K. (2009) On the scent of speciation: the chemosensory system and its role in premating isolation. Heredity, 102 (1), 77-97. https://doi.org/10.1038/hdy.2008.55 Links
- Stokstad, E. (2017) New crop pest takes Africa at lightning speed. Science, 356 (6337), 473-474. https://doi.org/10.1126/science.356.6337.473 Links
- Toth, M., Löfstedt, C., Blair, B. W., Cabello, T., Farag, A. I., Hansson, B. S., Kovalec, B. G., Maini, S., Nesterov, E. A., Pajor, I., Sazonov, A. P., Shamshev, I. V., Subchev, M., Szöcs, G. (1992) Attraction of male turnip moths Agrotis segetum (Lepidoptera: Noctuidae) to sex pheromone components and their mixtures at 11 sites in Europe, Asia, and Africa. Journal of Chemical Ecology, 18 (8), 1337-1347. https://doi.org/10.1007/BF00994360 Links
- Tumlinson, J. H., Mitchell, E. R., Teal, P. E. A., Heath, R. R., Mengelkoch, L. J. (1986) Sex pheromone of fall armyworm, Spodoptera frugiperda (JE Smith). Journal of Chemical Ecology, 12 (9), 1909- 1926. https://doi.org/10.1007/BF01041855 Links
- Unbehend, M., Hänniger, S., Meagher, R. L., Heckel, D. G., Groot, A. T. (2013) Pheromonal divergence between two strains of Spodoptera frugiperda. Journal of Chemical Ecology, 39 (3), 364-376. https://doi.org/10.1007/s10886-013-0263-6 Links
- Unbehend, M., Hänniger, S., Vásquez, G. M., Juárez, M. L., Reisig, D., McNeil, J. N., Meager R. L., Jenkins, D. A., Heckel, D. G., Groot, A. T. (2014) Geographic variation in sexual attraction of Spodoptera frugiperda corn-and rice-strain males to pheromone lures. PloS One , 9(2), e89255. https://doi.org/10.1371/journal.pone.0089255 Links
- Witzgall, P., Kirsch, P., Cork, A. (2010) Sex pheromones and their impact on pest management. Journal of Chemical Ecology, 36 (1), 80-100. https://doi.org/10.1007/s10886-009-9737-y Links
- Wu, Q. L., He, L. M., Shen, X. J., Jiang, Y. Y., Liu, J., Hu, G., Wu, K. M. (2019) Estimation of the potential infestation area of newly-invaded fall armyworm Spodoptera frugiperda in the yangtze river valley of China. Insects, 10 (9), 298. https://doi.org/10.3390/insects10090298 Links