












 (pdf)
(pdf)




 SciELO
SciELO  SciELO
SciELO 
 Permalink
Permalink
Introducción
Los pastizales naturales de las regiones áridas y semiáridas del norte de México han estado sujetos a pastoreo desde la época de la colonia por ganado bovino y ovino y más recientemente, por caprinos, además de la fauna silvestre que lo habita, lo que ha inducido la pérdida gradual de especies nativas por efecto del pastoreo sin control (Mellink y Riojas 2020). El alarmante deterioro de los pastizales posterior a la revolución mexicana se ha atribuido a la sobreutilización del recurso (Jurado-Guerra et al. 2021) y se ha determinado que el sobrepastoreo o pastoreo intensivo ocasiona la disminución de las poblaciones de especies nativas (Kikoti et al. 2015). El principal efecto del pastoreo sobre el crecimiento de las plantas es la reducción de la capacidad fotosintética asociada con el consumo principalmente de hojas, lo que reduce el flujo de elementos hacia el resto de la planta. Una planta pastoreada en más de una ocasión y antes de que esta se recupere su condición original, ocasiona un debilitamiento y eventualmente, provoca la muerte de las plantas. Este fenómeno es más notorio sobre las plantas que habitualmente son preferidas por los animales (Torrano y Valderrábano 2004). La evaluación de la respuesta de una planta sujeta a pastoreo, en términos de capacidad de rebrote y producción de forraje, es importante para la elección de alternativas de manejo, tales como hacer una mejor asignación en la carga animal, rotación y duración del pastoreo (Chávez et al. 2000, Jurado-Guerra et al. 2021), inclusive, podría potencialmente ayudar a seleccionar plantas para ser usadas en programas de revegetación. La recuperación de las plantas se puede evaluar de diferentes maneras, sin embargo, conocer las tasas de elongación y la acumulación de forraje es indispensable para conocer la respuesta de las diferentes especies. La tasa de elongación permite saber a qué velocidad se recuperan las plantas y más, si lo relacionamos con la intensidad de uso, de esta manera se puede observar la diferente respuesta que tiene una especie con respecto a otra sometida a mismo grado de utilización o corte.
Es necesario saber la respuesta de diferentes especies a distintas intensidades de uso para poder tener elementos que ayuden en la selección de las mejores especies a considerar en programas de revegetación de agostaderos y conocer cuales especies tienen mayor probabilidad de supervivencia y brindan adecuada cantidad de forraje de calidad (Rohner y Ward 1997). El término resistencia al pastoreo, dentro del manejo de pastizales, se refiere a la habilidad de las plantas para sobrevivir y crecer en condiciones de pastoreo extensivo, por lo que la capacidad para recuperar su condición original después del pastoreo es característica deseable de las plantas forrajeras. Las estrategias de resistencia a la defoliación incluyen el escape y la tolerancia al pastoreo, estrategias que se basan en mecanismos propios de la planta que le confiere resistencia para sobrevivir (Marriot y Carrere 1998). El escape al pastoreo consiste en mecanismos estructurales, bioquímicos y mecánicos que reducen el acceso o disponibilidad de los tejidos vegetales para los animales, mientras que la tolerancia al pastoreo consiste en mecanismos que promueven el crecimiento después de la defoliación, como la posición de meristemos de crecimiento a nivel del suelo y procesos de reacción, como la fotosíntesis compensatoria, distribución de carbono y reservas de carbohidratos, mecanismos que varían entre especies (Fortes et al. 2004).
La mayoría de los estudios sobre la respuesta al pastoreo se han realizado con gramíneas, existiendo limitada información en plantas arbustivas. A pesar de la importancia forrajera que tienen los arbustos que conforman la flora de los pastizales semiáridos (Guerrero-Cervantes et al. 2009, Guerrero et al. 2010), es poca la información disponible sobre su capacidad de producción de forraje, su respuesta a la defoliación y a la intensidad de uso. Por tanto, el presente estudio se realizó con el objetivo de evaluar la capacidad de recuperación de siete especies de arbustos forrajeros de zonas áridas después de haber sido sometidos a tres intensidades de corte durante la estación de crecimiento.
Materiales y métodos
Localización del área de estudio
El estudio se desarrolló en el Campo Experimental San Luis del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, ubicado en el municipio de Soledad de Graciano Sánchez, San Luis Potosí (22° 13’ 46’’ LN, -100° 51’ 01’ LO y 1 837 msnm), el cual forma parte del Desierto Chihuahuense. El experimento se estableció en el año 2001 y la evaluación se realizó durante el periodo de lluvias del año 2005.
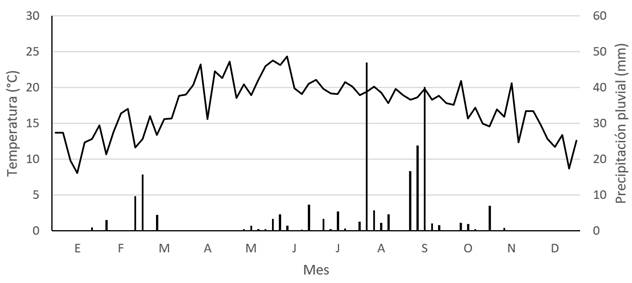
El clima se considera desértico frio de acuerdo con la clasificación de Köppen, cuya temperatura media anual es de 16.8 °C con mínima promedio de 10.0 °C, máxima promedio de 23.6 °C y precipitación media anual de 332.2 mm. Los meses más húmedos son junio a septiembre, con alrededor de 236 mm, y los más secos diciembre a mayo, con alrededor de 51.7 mm (Figura 1, Medina et al. 2005).
Establecimiento del experimento
Se utilizaron plantas de seis especies de arbustos forrajeros nativos del Desierto Chihuahuense: chamizo (Atriplex canescens (Pursh, Nutt.), ramoncillo (Dalea bicolor, Willd), mariola (Parthenium incanum, Kunth), vara dulce (Eysenhardtia polystachya, Ort. Sarg), escobilla (Buddleja scordioides, Kunth), guajillo (Acacia berlandieri, Benth) y una especie introducida, el cenizo (Atriplex nummularia, Lindl). Inicialmente se realizó la colecta de semillas de los arbustos señalados en el Altiplano Potosino y la semilla de la especie introducida se obtuvo de plantas existentes dentro del Campo Experimental de INIFAP. Una vez que se tuvieron las semillas de todas las especies, se procedió a aplicarles un método de escarificación, el cual consistió en sumergir las semillas en agua durante 48 horas, previas a la siembra en almácigo. Una vez emergidas las plantas y cuando alcanzaron una altura promedio de 12 cm fueron trasplantadas en bolsas de polietileno negro. Posteriormente, cuando las plantas tuvieron entre 25 y 30 cm de altura fueron trasplantadas manualmente en campo con una distancia entre plantas de 1.0 m. El experimento se estableció en un área de 870 m2 con suelo tipo castañozem de textura franco-arcillo-arenosa, con 1.4% de materia orgánica y pH de 8.1.
Diseño del experimento
El área experimental consistió en siete hileras de 30 m de largo, los cuales se subdividieron en tres secciones de 10 m cada uno, conformando 21 secciones o parcelas. Las parcelas se distribuyeron aleatoriamente a las siete especies (tres parcelas por especie). En cada parcela se plantaron nueve plantas con una distancia de 1 m de separación. En cada parcela se asignaron aleatoriamente tres plantas a cada uno de las tres intensidades de corte: baja, media y alta. Al inicio del experimento, se registraron las alturas promedio de cada especie, siendo estas como se muestran en la tabla 1.
Tabla 1 Altura inicial de las especies de arbustos forrajeros consideradas para la aplicación de las intensidades de corte.
| Especie | Altura inicial (cm) |
|---|---|
| Chamizo (Atriplex canescens) | 1.34 |
| Ramoncillo (Dalea bicolor) | 1.22 |
| Mariola (Parthenium incanum) | 0.64 |
| Vara dulce (Eysenhardtia polystachya) | 1.54 |
| Escobilla (Buddleja scordioides) | 0.48 |
| Guajillo (Acacia berlandieri) | 1.18 |
| Cenizo (Atriplex nummularia) | 1.26 |
Las intensidades de corte consistieron en retirar con tijera, de arriba hacia abajo, el 25, 50 y 75% del follaje de las plantas. La intensidad baja fue el corte del 25% de la altura de la planta, quedando en pie el 75% (hojas y tallos); de manera similar se aplicó a la intensidad media 50% y alta 75%. Se aplicó un corte inicial a la mitad de la estación de crecimiento (2 de agosto) y otro al final (14 de noviembre), registrando en ambos casos la altura de la planta antes y después del corte. Con la altura inicial de las plantas se define la intensidad de corte, la altura final de las plantas después del primer corte quedo como referencia, ya que se realizó un segundo corte a la misma altura después de un periodo de tiempo determinado y la diferencia en altura de las plantas entre el primer y segundo corte fue la tasa de elongación de las plantas. Todas las hojas y tallos con diámetro menor a 3 mm (Vernier TRUPER®) obtenidos en el segundo corte, fueron separados, pesados en báscula de precisión (0.5 x 0.001 kg, TRANSCELL Technology®) y secados en horno (FELISA©) a 70 ⁰C por 48 h para estimar la producción de materia seca.
Variables estudiadas
Tasa de elongación (TE)
Estima el crecimiento de la planta y está representada por la diferencia de altura de la planta antes del segundo corte (A2C) y la altura después del primer corte (A1C), dividido entre el número de días entre ambas mediciones (cm/día). Se refiere al incremento de lámina verde en un intervalo de tiempo dado.
Tasa de acumulación de forraje (AB)
Estima la acumulación de forraje en el periodo de crecimiento en base seca (g MS día-1). Para esto, se consideraron las hojas y ramas con diámetro inferior de 3 mm cosechadas en el segundo corte, producto del crecimiento ocurrido a partir del primer corte. considerando que es el forraje habitualmente consumido por el ganado caprino.
Análisis estadísticos
Se evaluó el efecto de la especie de arbusto forrajero (siete niveles: chamizo (Atriplex canescens, Pursh, Nutt.), ramoncillo (Dalea bicolor, Willd), mariola (Parthenium incanum, Kunth), vara dulce (Eysenhardtia polystachya, Ort. Sarg), escobilla (Buddleja scordioides, Kunth), guajillo (Acacia berlandieri, Benth) y cenizo (Atriplex nummularia, Lindl) y, el efecto de la intensidad de corte (tres niveles: baja, media y alta), por medio de un modelo lineal de efectos fijos:
Dónde: Yijk = variable respuesta, μ = media general, E i = efecto de la especie de arbusto, C j = efecto de la intensidad de corte, (EC) ij = interacción entre la especie de arbusto y la intensidad de corte y e (ijk) =error experimental.
Cuando hubo diferencia significativa se aplicó la Prueba de Tukey para detectar diferencias entre medias (α = 0.05). Cuando se detectó efecto de uno solo de los factores (P ≤ 0.05) sin interacción significativa entre factores, se aplicó la prueba de T de Student (α = 0.05) para comparar las medias dentro del mismo factor. Adicionalmente se correlacionó la tasa de elongación con la acumulación de forraje. Los resultados se expresan en medias ± error estándar de la media (± SEM). Todos los análisis se realizaron utilizando el paquete estadístico JMP Start Statistics (SAS 2004).
Resultados
La tasa de elongación de los arbustos forrajeros se muestra en el Tabla 2. En general, la tasa promedio de crecimiento diario fue de 0.299 ± 0.20 cm día-1. Se detectó una diferencia en la tasa de elongación entre especies (P < 0.0001), pero no entre intensidades de corte (P > 0.05). La interacción entre ambos factores fue significativa (P < 0.003). En la intensidad de corte bajo (25%) Vara dulce y Chamizo fueron los que tuvieron mayor tasa de elongación (P < 0.05); en la intensidad de corte medio (50%) Cenizo, Chamizo, Guajillo y Vara dulce tuvieron mayor tasa de elongación (P < 0.05); en la intensidad de corte alto (75%) el Chamizo se mantuvo con la mayor tasa de elongación (P < 0.05, Tabla 2).
Tabla 2 Tasa de elongación promedio (cm día-1 ± EE) de siete especies de arbustos forrajeros del sur del desierto chihuahuense sujetos a tres intensidades de corte.
| Arbusto | Intensidad de corte | ||
|---|---|---|---|
| Bajo (25%) | Medio (50%) | Alto (75%) | |
| Chamizo | 0.351 ± 0.057 ab | 0.418 ± 0.038 ab | 0.493 ± 0.027 a |
| Ramoncillo | 0.150 ± 0.043 c | 0.238 ± 0.043 bc | 0.359 ± 0.070 b |
| Mariola | 0.075 ± 0.020 d | 0.145 ± 0.038 c | 0.218 ± 0.048 c |
| Vara dulce | 0.551 ± 0.115 a | 0.354 ± 0.079 ab | 0.216 ± 0.082 c |
| Escobilla | 0.156 ± 0.036 c | 0.173 ± 0.034 c | 0.236 ± 0.034 c |
| Guajillo | 0.286 ± 0.050 bc | 0.417 ± 0.082 ab | 0.348 ± 0.046 b |
| Cenizo | 0.180 ± 0.044 c | 0.425 ± 0.093 a | 0.273 ± 0.054 bc |
abcd, Valores con distinta letra en la misma columna indica diferencias significativas (Tukey α = 0.05) en la tasa de elongación (cm día-1).
La acumulación de forraje, estuvo fuertemente afectada tanto por la especie de arbusto como por la intensidad de corte (P < 0.0001), detectando interacción significativa entre los dos factores (P < 0.003). En general, los arbustos mostraron una acumulación diaria de forraje en promedio de 3.39 ± 5.0 g día-1, tendiendo a incrementarse con la mayor intensidad de corte. Chamizo fue la especie que más forraje acumuló con las tres intensidades de corte (Tabla 3) observando la mayor acumulación con intensidad de corte alto (75%) con 14.65 ± 2.34 g día-1, lo que significa una muy buena respuesta de recuperación después de una defoliación intensa. En las tres intensidades de corte, Ramoncillo fue la especie que menos forraje acumuló, lo que indica que tiene poca capacidad de rebrote.
Tabla 3 Acumulación promedio de forraje (g d-1 ± EE) de siete especies de arbustos forrajeros del sur del desierto Chihuahuense, sometidos a tres intensidades de corte.
| Arbusto | Intensidad de corte | ||
|---|---|---|---|
| Bajo (25%) | Medio (50%) | Alto (75%) | |
| Chamizo | 5.42 ± 1.25a | 12.22 ± 2.33a | 14.65 ± 2.34a |
| Ramoncillo | 0.37 ± 0.13c | 0.81 ± 0.24d | 0.69 ± 0.10e |
| Mariola | 0.59 ± 0.10bc | 0.83 ± 0.19d | 1.43 ± 0.34d |
| Vara dulce | 1.12 ± 0.42b | 1.38 ± 0.34c | 4.68 ± 1.77bc |
| Escobilla | 0.93 ± 0.11b | 4.33 ± 1.04b | 6.91 ± 1.11b |
| Guajillo | 0.67 ± 0.21b | 0.81 ± 0.27d | 1.79 ± 0.62d |
| Cenizo | 0.77 ± 0.61b | 4.13 ± 1.52b | 3.06 ± 1.68c |
abcd, Valores con distinta letra en columnas indican diferencia significativa (Tukey, 0.05) en la acumulación diaria de forraje (g día-1).
La correlación entre la tasa de elongación y la acumulación de forraje fue baja pero significativa (R = 0.28; P < 0.0001). Los valores de correlación variaron de 0.14 en Vara dulce a 0.62 en Mariola (Tabla 4).
Tabla 4 Correlación entre la tasa de elongación y la acumulación de forraje en siete especies de arbustos forrajeros del sur del desierto Chihuahuense, sometidos a tres intensidades de corte.
| Arbusto | R | P |
|---|---|---|
| General | 0.28 | 0.0001 |
| Chamizo | 0.30 | >0.05 |
| Ramoncillo | 0.48 | <0.02 |
| Mariola | 0.62 | <0.0005 |
| Vara dulce | 0.14 | >0.05 |
| Escobilla | 0.39 | <0.05 |
| Guajillo | 0.41 | <0.05 |
| Cenizo | 0.32 | >0.05 |
Discusión
Para la rehabilitación o revegetación de pastizales, establecimiento de sistemas silvopastoriles o establecimiento de bancos de proteína con arbustivas forrajeras, es importante que las plantas que se seleccionen muestren buena capacidad para recuperarse después del pastoreo en ambientes áridos y semiáridos. Los resultados muestran que algunas especies responden mejor a una intensidad de corte elevada mientras otros responden mejor con intensidades de corte moderadas. La evaluación de las tasas de elongación de tallos y de acumulación de forraje de plantas forrajeras es de gran importancia, tanto para fines de sostenibilidad como para el diseño de sistemas de pastoreo (Fortes et al. 2004). Los resultados determinaron que las tasas de elongación y acumulación de forraje fueron diferentes entre las especies estudiadas, lo cual se debe, en parte, a la distinta arquitectura observada que muestran las diferentes especies de arbustos, de donde depende la respuesta en función al número de tallos por planta, la longitud de tallos, disposición de las hojas y el tamaño promedio de la especie.
Se observa que cada especie tiene su propia dinámica de crecimiento, lo cual es de importancia porque define que cada especie requiere un manejo diferente, en función a su respuesta a la defoliación, debido a las diferentes capacidades de recuperación tras la defoliación de las especies evaluadas. La buena capacidad de recuperación de una especie de arbusto en particular, es indicativo de alta adaptación al medio ambiente y a su propia arquitectura, la cual se ha desarrollado a través del tiempo y la caracteriza dentro del ecosistema. El chamizo, en general, fue la especie que mostró la mejor respuesta a la defoliación, siendo además muy apetecida por el ganado caprino y tiene del 16 al 20% de proteína (Romero y Ramírez 2003).
En las condiciones de pastoreo extensivo con cabras, los arbustos forrajeros suelen ser defoliados de forma intensa, por lo que aquellas especies poco tolerantes al pastoreo tienden a desaparecer paulatinamente. Los resultados muestran que la mayoría de las especies evaluadas mostraron buena acumulación de forraje cuando se les aplicó una alta intensidad de corte, con excepción de ramoncillo, mariola y guajillo. La tasa de elongación tendió a ser mayor al incrementar la intensidad de corte, específicamente en chamizo. Estos resultados evidencian que cada especie se comporta de manera única, dependiendo de la arquitectura de la planta, de la forma de crecimiento y de estrategias de sobrevivencia que tiene cada una de ellas. La acumulación de material forrajero fue mayor en chamizo (Tabla 3), superando por más del doble a las otras especies. En general, la acumulación de forraje se incrementó con la mayor intensidad de corte, sugiriendo que esta especie es capaz de tolerar una elevada intensidad de corte y aun así, responder con una elevada producción en condiciones áridas y semiáridas (Saucedo 1998). Al reducirse el área foliar mediante el pastoreo, las plantas defoliadas pueden presentar mayores estímulos para incentivar una mayor fotosíntesis que en las no defoliadas (Fortes et al. 2004). Se ha observado que la defoliación del estrato superior incrementa la actividad fotosintética, debido a que permite una mayor penetración de luz solar (Quentin et al. 2011). La intensidad de defoliación afecta tanto las reservas de carbono (Wang et al. 2020) como de carbohidratos (Wiley et al. 2017), ambos esenciales para el rebrote de las plantas. En el caso de ramoncillo, fue la especie que menos forraje acumuló en las tres intensidades de corte, lo que indica que, tiene poca capacidad de rebrote, lo cual fue consignado por García-Cervantes et al. (2023) quienes señalan que, en esta especie, de cuatro estaciones muestreadas solo en otoño y verano presento follaje.
Un resultado interesante fue que las mayores tasas de elongación no estuvieron en todos los casos asociadas con mayor acumulación de forraje (Tabla 4). Sólo en cuatro de las siete especies la tasa de elongación se relacionó con la acumulación de forraje. Se ha observado que la asociación entre altura de la planta y acumulación de forraje difiere de una especie a otra, esto en función a la diferente arquitectura que muestran las especies, básicamente en lo referente a la relación hoja - tallo (Villalobos 2007).
Conclusiones
Se encontró que las tasas de elongación y de acumulación de forraje de diferentes especies arbustivas fueron estadísticamente diferentes, determinadas por las características propias de cada especie, las cuales presentan diferente arquitectura, forma de crecimiento y estructura. Con respecto a la tasa de elongación, sobresale el chamizo, con excelente respuesta a la mayor intensidad de corte. Con respecto la acumulación de forraje, también el chamizo mostró tener excelente capacidad de recuperación a la defoliación, en las tres intensidades de corte. En general, es recomendable conocer la respuesta de los arbustos forrajeros presentes en los agostaderos de las zonas áridas a diferentes intensidades de corte para una planeación eficiente de programas de revegetación de agostaderos degradados y establecimiento de bancos de forraje con especies arbustivas.

