












 (pdf)
(pdf)




 SciELO
SciELO  SciELO
SciELO 
 Permalink
Permalink
Introducción
Las zonas hiperáridas, áridas, semiáridas y subhúmedas secas, consideradas ecológicamente vulnerables y frágiles (Koutroulis, 2019), ocupan aproximadamente 47% de la superficie terrestre y se encuentran en expansión (Guida Johnson, 2021). En estas áreas, los procesos que generan degradación y desertización de la tierra son consecuencia de la combinación de factores climáticos adversos sumados a la sobreexplotación de los recursos naturales (Azzouzi et al., 2017). Esta situación impacta sobre dos mil millones de personas que viven en 100 países que poseen zonas áridas y semiáridas a nivel mundial (Gómez López, 2020), quienes ven afectadas su seguridad alimentaria, su economía, y en general, su sostenibilidad (Gomiero, 2016; Hossain et al., 2020).
Debido a que los pastizales constituyen la vegetación principal en las tierras áridas y que poseen un valor ecológico y socioeconómico importante (Zhao et al., 2024), una estrategia de recuperación de las tierras degradadas es implementar prácticas de manejo del suelo mediante la siembra de especies forrajeras anuales o perennes nativas (Quiroga y Correa, 2011; Tácuna et al., 2015; Gonçalves et al., 2023). En este sentido, la siembra de semillas de gramíneas típicas de las zonas áridas en Argentina, como Leptochloa crinita (Lag.) Parodi, Pappophorum caespitosum Fries y Digitaria californica (Benth.) Henrard, adaptadas a la escasez de agua y a las altas temperaturas (Villagra et al., 2011), sería una alternativa viable para la recuperación de suelos degradados, debido al bajo costo y alta efectividad que presentan (Scarfó et al., 2024). Sin embargo, la germinación de los cariopses de estas especies y el posterior establecimiento de plántulas pueden verse afectados si las condiciones ambientales en el momento de la emergencia no son adecuadas. De acuerdo a lo expresado, resulta vital para el éxito de prácticas de recuperación de suelos degradados, conocer los requisitos de germinación y emergencia de tales especies (Merino-Martín et al., 2017; Leger et al., 2019).
Entre los principales factores que afectan la germinación de las semillas se encuentran la disponibilidad de agua, la temperatura y la luz (cantidad y calidad o longitud de onda) (Ramírez Segura et al., 2022). Cuando las condiciones de humedad no son limitantes, la germinación está regulada por la temperatura (Bolaños et al., 2019), debido a que se relaciona con la activación de las enzimas que regulan la velocidad de las reacciones bioquímicas que ocurren en la semilla tras su hidratación (Caroca et al., 2016). Por otro lado, la presencia o ausencia de luz puede ser determinante en la germinación, dependiendo de si las semillas son fotoblásticas positivas o negativas (Villena et al., 2024). Además, las especies pertenecientes a la familia Poaceae presentan otro factor que afecta la germinación: la presencia de brácteas accesorias (glumas, palea, lema) que cubren a los cariopses (Quero Carrillo et al., 2017). Esas brácteas imponen latencia en las semillas actuando como barrera física, para la emergencia de la radícula (Duclos et al., 2013), o como inhibidores para la germinación (Oreja et al., 2017).
La información sobre los requerimientos de las especies de zonas áridas para la germinación y el establecimiento exitoso de las plantas constituye el punto de partida para su domesticación y uso en programas de recuperación de suelos degradados, restauración ecológica o con fines productivos como forraje para el ganado doméstico. Debido a lo expuesto, los objetivos planteados en este trabajo fueron: 1) Determinar el efecto de la luz, de la presencia de espiguilla y la temperatura sobre la germinación de L. crinita, P. caespitosum y D. californica y 2) Evaluar el efecto de la interacción de los factores luz, espiguilla y temperatura en el proceso de germinación de cada especie bajo estudio.
Materiales y Métodos
Material vegetal
Las semillas de Leptochloa crinita fueron obtenidas de la producción que realiza la Estación Experimental Agropecuaria Rama Caída, del Instituto Nacional de Tecnología Agropecuaria (INTA), departamento San Rafael, Provincia de Mendoza, Argentina. Los cariopses usados en este trabajo corresponden a la cosecha realizada en 2015. Pappophorum caespitosum fue colectada en otoño de 2015 en la zona de La Quebrada de las Burras a 1848 m s.n.m., departamento Ullúm, provincia de San Juan, Argentina (31°1'59.6''S, 68°56'33.2''O). Digitaria californica se recolectó en el otoño de 2015 en la localidad Bermejo a 630 m s.n.m., departamento Caucete, provincia de San Juan, Argentina (31°41'57.5''S, 67°49'5.8''O).
Pappophorum caespitosum y Digitaria californica fueron colectadas de al menos 30 individuos seleccionados al azar, de acuerdo con Di Sacco et al. (2020). En laboratorio, se eliminaron los restos vegetales dejando solo las espiguillas, las cuales fueron colocadas en un recipiente de vidrio con cierre hermético y conservadas con una humedad de 12%, con sílica gel en su interior, y guardado en heladera (Briket exhibidora, Rosario, Argentina) a 3 °C (banco de germoplasma ARG 1416) hasta su uso, en marzo de 2018.
La viabilidad de las semillas conservadas de cada especie se evaluó cada año, desde la recolección, hasta la actualidad mediante controles de monitoreo de la germinación, los cuales mostraron que se mantuvo sin variaciones. Las espiguillas de las tres especies se escarificaron entre dos gomas de caucho corrugado y luego se limpiaron en un soplador de semillas (Servicios Mecatrónicos, Mod. 2023, Argentina) con columna para semillas pequeñas para obtener los cariopses desnudos.
Germinación
Los propágulos (espiguillas y cariopses) fueron sumergidos en hipoclorito de sodio al 3% durante 5 minutos, y luego lavados tres veces con agua destilada. Los tratamientos aplicados fueron tres: 1) iluminación (luz y oscuridad); 2) tipo de propágulo (cariopse y espiguilla), y 3) temperatura (15, 20, 25, 30 y 35 °C). El estudio contempló también la interacción de los factores para cada especie de estudio. Se sembraron 25 cariopses o espiguillas de cada especie en cajas Petri estériles con papel filtro neutro (S&S tipo 0859, 90 mm Ø) humedecido con 5 ml de agua destilada, con seis repeticiones por tratamiento. Se aplicó un fotoperiodo de 12 h de luz y 12 h de oscuridad
Las condiciones de luz se brindaron con tubos fluorescentes de 36 w dispuestos verticalmente dentro de la cámara de germinación (Servicios Mecatrónicos Mod. 2015, Argentina). La intensidad de la luz fue cuantificada con una barra de medición de radiación fotosintéticamente activa (Apogee Quantum Flux, Mod MQ-306, Logan, UT, EUA), la cual mostró un valor de 185 µmol/m2/s. Para brindar oscuridad en los tratamientos que requirieron esta condición, las cajas de Petri fueron envueltas con papel de aluminio (Ferrari y Parera, 2015) y colocadas dentro de una bolsa de papel Kraft. En este caso, la germinación se evaluó en un cuarto oscuro iluminado solamente con un tubo de luz azul de 18 W. La germinación se evaluó cada 24 h, durante 5 días. Después del tercer día de la siembra no se registró nueva germinación. Los cariopses germinados fueron retirados de las cajas. Se consideró que una espiguilla o cariopse había germinado cuando la radícula fue visible (2 mm).
Para cada tratamiento se estimó el porcentaje de germinación (G) y el Tiempo Medio de Germinación (TMG), el cual expresa en días, el tiempo necesario para que germinen 50% de los propágulos. La estimación del TMG se realizó de acuerdo con Bewley y Black (1994), aplicando la siguiente ecuación:
Dónde: D es el número de días transcurridos desde el inicio de la germinación, n es el número de semillas germinadas el día D, nt es el número total de semillas germinadas.
Análisis estadístico
El experimento se realizó para cada especie con un arreglo factorial de tres factores, utilizando un diseño completamente al azar con seis repeticiones por tratamiento. El análisis estadístico se realizó con el software Infostat Profesional 2020 (Di Rienzo et al., 2020). Las respuestas evaluadas fueron porcentaje de germinación y tiempo medio de germinación (TMG). Para los casos de análisis de valores de porcentaje de germinación, los resultados fueron transformados con la función raíz cuadrada del arcoseno (Orantes García et al., 2019). Los resultados se analizaron mediante un ANOVA de tres vías, previa validación de los supuestos de normalidad y homogeneidad de la varianza mediante las pruebas de Shapiro-Wilk y Levene, respectivamente. En los casos en que el análisis fue significativo, se utilizó la prueba de LSD Fisher (α=0.05) como prueba a posteriori de separación de medias.
Resultados
Para las tres especies bajo estudio, el efecto de cada tratamiento evaluado de manera individual fue significativo (p<0.0001) para las variables evaluadas (G y TMG), a excepción de la temperatura en el porcentaje de germinación de Pappophorum caespitosum. En la interacción de los tratamientos evaluados, se observó una respuesta diferencial en cada especie en las variables G y TMG (Cuadro 1).
Cuadro 1: Valores de significancia (p <0.0001) obtenidos en las variables porcentaje de germinación (G) y tiempo medio de germinación (TMG) para cada tratamiento aplicado (iluminación, tipo de propágulo y temperatura) y sus interacciones, en Leptochloa crinita (Lag.) Parodi (error 5202.55, g.l. 100), Pappophorum caespitosum Fries (error 3603.18, g.l. 99) y Digitaria californica (Benth.) Henrard (error 4795.08, g.l. 100), pastos de zonas áridas de Argentina.
| Fuente de variación | Leptochloa crinita (Lag.) Parodi | Pappophorum caespitosum Fries | Digitaria californica (Benth.) Henrard | |||||||
| g.l. | F | G | TMG | F | G | TMG | F | G | TMG | |
| Iluminación (I) | 1 | 52.57 | <0.0001 | <0.0001 | 145.57 | <0.0001 | <0.0001 | 22.47 | <0.0001 | <0.0041 |
| Propágulo (P) | 1 | 564.41 | <0.0001 | <0.0001 | 700.28 | <0.0001 | <0.0001 | 64.70 | <0.0001 | <0.0001 |
| Temperatura (T) | 4 | 12.72 | <0.0001 | <0.0001 | 2.48 | 0.3422 | <0.0001 | 100.65 | <0.0001 | <0.0001 |
| I × P | 1 | 2.21 | 0.1407 | 0.7516 | 41.96 | <0.0001 | <0.0001 | 0.99 | 0.3210 | 0.4492 |
| I × T | 4 | 2.10 | 0.0862 | <0.0001 | 2.50 | 0.3135 | <0.0001 | 19.44 | <0.0001 | <0.0001 |
| P × T | 4 | 6.29 | 0.0001 | <0.0001 | 2.12 | 0.6379 | <0.0001 | 28.79 | <0.0001 | 0.7212 |
| I × P × T | 4 | 1.69 | 0.1580 | <0.0001 | 0.80 | 0.4094 | 0.1259 | 13.93 | <0.0001 | <0.0001 |
En Leptochloa crinita, la interacción entre el tipo de propágulo y la temperatura fue significativa (p<0.0001) en ambas variables (Cuadro 1). El valor promedio (±error estándar) máximo de germinación (72.56%±2.59) fue a 25 °C en cariopses. No se observaron diferencias germinativas en la germinación de cariopses a 25 °C con respecto a las temperaturas 30, 20 y 15 °C (Fig. 1). Al evaluar la germinación a partir de espiguillas, el máximo valor promedio se registró también a los 25 °C. No se encontraron diferencias significativas con los resultados obtenidos de germinación a las 20 y 30 °C (Fig. 1).
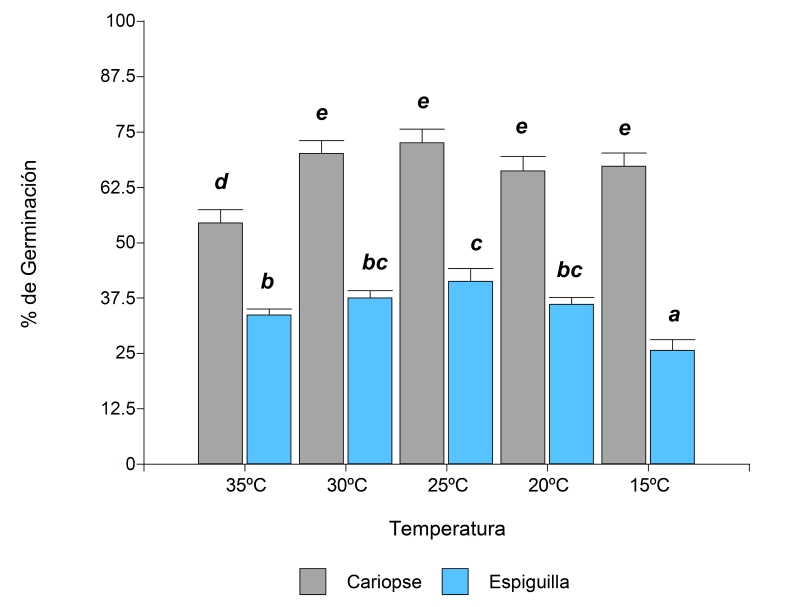
Figura 1: Porcentaje de germinación de la especie Leptochloa crinita (Lag.) Parodi en la interacción de los factores iluminación (luz, oscuridad) y temperatura (15, 20, 25, 30 y 35 °C). Las letras diferentes indican diferencias significativas entre los valores promedio de cada tratamiento de acuerdo con la prueba a posteriori de LSD Fisher (α˂0.05).
Los resultados obtenidos del TMG en L. crinita mostraron las interacciones significativas de los tratamientos iluminación y temperatura, tipo de propágulo y temperatura y en la triple interacción entre iluminación, tipo de propágulo y temperatura (Cuadro 1). En esta especie, el valor promedio más bajo de TMG a partir de cariopses se registró en el tratamiento de oscuridad a 35 °C. Se observaron diferencias significativas con el resto de las temperaturas evaluadas (Cuadro 2). En presencia de luz, el TMG promedio más bajo de cariopses fue en el tratamiento en el que se aplicó una temperatura de 35 °C. En el caso de las espiguillas, el tratamiento de oscuridad a 20°C fue el que mostró el menor valor promedio y su error estándar (0.51±0.11) para esta variable (Cuadro 2). No se encontraron diferencias significativas entre este tratamiento y espiguilla a 35 °C y cariopse en oscuridad también a 35 °C.
Cuadro 2: Valores promedio y error estándar (±E.E.) del TMG para Leptochloa crinita (Lag.) Parodi y Digitaria californica (Benth.) Henrard en la triple interacción de los tratamientos iluminación (luz, oscuridad), tipo de propágulo (cariopse, espiguilla) y temperatura (15, 20, 25, 30 y 35 °C). Las letras diferentes en una misma fila indican diferencias significativas entre las medias de los tratamientos de acuerdo con la prueba a posteriori de LSD Fisher (α˂0.05).
| Tratamiento | Leptochloa crinita (Lag.) Parodi | Digitaria californica (Bentd.) Henrard | ||||||
| Cariose | Espiguilla | Cariose | Espiguilla | |||||
| Luz | Oscuridad | Luz | Oscuridad | Luz | Oscuridad | Luz | Oscuridad | |
| 15°C | 3.93±0.16d | 2.63±0.23d | 1.35±0.14bc | 1.37±0.15c | 1.60±0.31b | 1.13±0.07bc | 1.11±0.08a | 2.72±0.038c |
| 20°C | 2.23±0.09c | 2.01±0.11c | 1.73±0.07d | 0.51±0.06a | 2.66±0.35a | 2.55±0.05d | 3.22±0.29c | 2.89±0.15c |
| 25°C | 1.32±0.05b | 1.19±0.10b | 1.67±0.18cd | 1.07±0.13bc | 1.35±0.07b | 1.22±0.08c | 2.05±0.15b | 1.07±0.12b |
| 30°C | 0.95±0.02a | 1.01±0.08b | 1.31±0.09ab | 0.92±0.06b | 1.06±0.08ab | 0.86±0.13b | 1.43±0.05a | 1.03±0.06b |
| 35°C | 0.85±0.08a | 0.63±0.07a | 0.82±0.03a | 0.85±0.11b | 0.49±0.06a | 0.56±0.14a | 1.51±0.07a | 0.17±0.04a |
En la germinación de P. caespitosum se observó una interacción significativa (p<0.0001) entre los tratamientos iluminación y tipo de propágulo (Cuadro 1). El valor promedio más elevado de G y su error estándar (73.93%±1.10) se registró en el tratamiento con luz a partir de cariopses. Se encontraron diferencias significativas entre los cuatro tratamientos (Fig. 2). En esta especie, el TMG resultó significativo para las dobles interacciones entre los tratamientos, mientras que la triple interacción no lo fue (Cuadro 1). El valor promedio menor de TMG y su error estándar (1.01±0.11) se observó en oscuridad a 25 °C, sin diferencias significativas con oscuridad a 30 °C (Cuadro 3). A partir de cariopses, el valor promedio más bajo de TMG y su error estándar (1.02±0.11) en esta especie se encontró a 30 °C, sin diferencias significativas con el valor encontrado a 35 °C (Cuadro 3). En la interacción del tipo de propágulo con el tipo de iluminación, el menor valor promedio de TMG y su error estándar (1.55±0.07) se registró en espiguilla con oscuridad, sin diferencias significativas con el tratamiento luz y espiguilla (Cuadro 3).
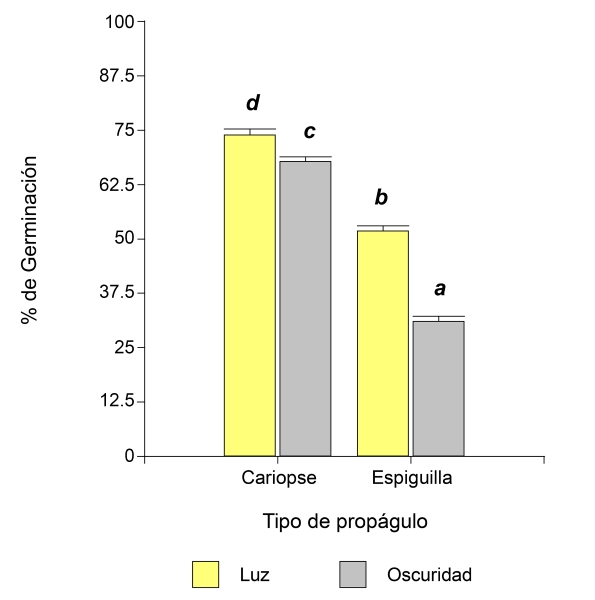
Figura 2: Porcentajes de germinación de la especie Pappophorum caespitosum Fries en la interacción de iluminación (luz, oscuridad) y tipo de propágulo (cariopse, espiguilla). Letras diferentes sobre las columnas indican diferencias significativas entre los valores medios de los tratamientos de acuerdo con la prueba a posteriori de LSD Fisher (α˂0.05).
Cuadro 3: Valores promedio y error estándar (±E.E.) del TMG de Pappophorum caespitosum Fries en las interacciones de los tratamientos de iluminación (luz, oscuridad) y temperatura (15, 20, 25, 30 y 35 °C), y tipo de propágulo (cariopse, espiguilla) y temperatura. Letras diferentes en la columna de un mismo tratamiento indican diferencias significativas entre los valores medios de acuerdo con la prueba a posteriori de LSD Fisher (α˂0.05).
| Tratamientos | 15°C | 20°C | 25°C | 30°C | 35°C |
| Luz | 4±0.17b | 2.55±0.27a | 2.15±0.14b | 1.61±0.22a | 1.74±0.24b |
| Oscuridad | 2.23±0.29a | 2.21±0.16a | 1.01±0.06a | 1.50±0.15a | 1.07±0.06a |
| Cariose | 3.79±0.22b | 1.82±0.07a | 1.48±0.14a | 1.02±0.04a | 1.09±0.03a |
| Espiguilla | 2.44±0.35a | 2.94±0.20b | 1.68±0.24a | 2.10±0.13b | 1.71±0.25b |
Los resultados mostraron que la única interacción de los factores evaluados que no fue significativa en la especie D. californica para el porcentaje de germinación, fue iluminación y tipo de propágulo (Cuadro 1). En la interacción tipo de propágulo y temperatura, el valor promedio de G más elevado y su error estándar (77.90±2.00) se encontró en cariopse a 25 °C, sin diferencias significativas con cariopse a 20 °C (Fig. 3). Con respecto a la triple interacción de los factores, se encontró que el porcentaje de germinación en cariopses fue mayor en oscuridad a 20 °C (78.93%±2.63), sin diferencias significativas con el tratamiento bajo las mismas condiciones, pero a 25 °C (Cuadro 4). La interacción entre la iluminación, el tipo de propágulo y la temperatura afectó al TMG (Cuadro 1). Para esta variable, el valor más bajo y su error estándar (0.17±0.17) se observó a partir de cariopses en oscuridad y a 35 °C (Cuadro 2). No se observaron diferencias significativas entre esta interacción y cariopse con luz a 35 °C y cariopse en oscuridad a 35 °C.
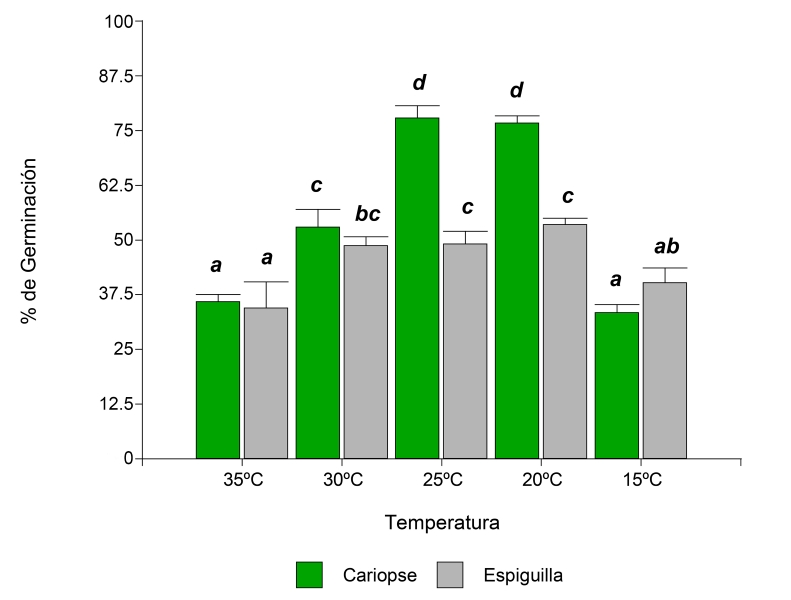
Figura 3: Porcentajes de germinación de la especie Digitaria californica (Benth.) Henrard en la interacción de los factores tipo de propágulo (cariopse, espiguilla) y temperatura (15, 20, 25, 30 y 35 °C). Letras diferentes sobre las columnas indican diferencias significativas entre los valores medios de los tratamientos de acuerdo con la prueba a posteriori de LSD Fisher (α<0.05).
Cuadro 4: Valores promedio y error estándar (±E.E.) del porcentaje de germinación de la especie Digitaria californica (Benth.) Henrard en las interacciones de los tratamientos iluminación (luz, oscuridad), tipo de propágulo (cariopse, espiguilla) y temperatura (15°C, 20°C, 25°C, 30°C y 35°C). Letras diferentes en una misma columna indican diferencias significativas entre las medias de los tratamientos de acuerdo con la prueba a posteriori de LSD Fisher (α˂0.05).
| Tratamientos | Cariopse | Espiguilla | ||
| Luz | Oscuridad | Luz | Oscuridad | |
| 15°C | 34.44±3.53a | 32.33±1.03a | 32.33±1.03 a | 48.13±4.76bc |
| 20°C | 74.56±1.37c | 78.93±2.63c | 52.12±2.46 b | 54.82±1.64c |
| 25°C | 77.39±4.31c | 78.41±3.72c | 51.54±2.32 b | 46.56±5.40bc |
| 30°C | 65.07±3.03b | 40.73±2.05b | 54.39±1.37 b | 43.08±1.85b |
| 35°C | 37.20±1.68a | 34.61±2.91ab | 53.66±2.20 b | 15.26±1.76a |
Discusión
Se considera que las semillas de tamaño pequeño, como son los cariopses de las especies de pastos estudiadas, tienen una respuesta positiva a la luz (Entio et al., 2016), lo cual podría estar asociado con la ruptura de la latencia de las semillas (Baskin y Baskin, 2014). En este trabajo, la condición de oscuridad produjo una disminución significativa del porcentaje de germinación en las tres especies estudiadas.
En Leptochloa crinita se encontró una diferencia significativa en los valores de germinación entre la condición de luz y oscuridad. Según Weller et al. (2019), la luz es un factor importante en la germinación de diferentes especies del género Leptochloa P. Beauv. Esta condición también quedó demostrada en el trabajo realizado por Malagrina et al. (2011), quienes evaluaron el efecto de la luz a una temperatura constante en la germinación de Trichloris crinita (Lag.) P.M. Peterson & N.W. Snow (sinonimia de Leptochloa crinita) proveniente de diferentes sitios de colección, y en los tres casos evaluados obtuvieron una disminución de 50% en la germinación cuando el experimento se realizó en ausencia de luz. Por otro lado, Chauhan y Johnson (2008) reportaron una germinación completamente inhibida en Leptochloa chinensis (L). Nees en oscuridad.
En la germinación de P. caespitosum, la germinación disminuyó considerablemente (22%) cuando el experimento se llevó a cabo en condiciones de oscuridad permanente. Si bien la bibliografía disponible sobre los factores que afectan la germinación sobre el género Pappophorum Schreber es escasa, en particular sobre la especie P. caespitosum es aún menor. De hecho, no se han encontrado trabajos previos que evalúen el efecto de la luz o la interacción de este factor con otros factores abióticos en la germinación de esta especie. Entio et al. (2016) evaluaron el efecto de la luz sobre la germinación en diferentes poblaciones de Pappophorum vaginatum Buckley, en las que encontraron resultados similares a los de este trabajo, ya que registraron que la luz incrementó la germinación acumulada en esa especie, en comparación con la condición de oscuridad continua. En P. phillippianum Parodi, Ferrari y Parera (2015) encontraron un efecto positivo de la luz en la germinación de esta especie, en la interacción con las mismas temperaturas evaluadas en este trabajo.
En D. californica también se registró una disminución en el porcentaje de germinación cuando los experimentos se realizaron en ausencia de luz. Ferrari y Parera (2015) encontraron resultados similares a los informados en este trabajo, al comparar la germinación de esta especie en luz y oscuridad. Por otro lado, en Digitaria sanguinalis (L.) Scop., Oreja et al. (2017) reportaron los mayores porcentajes de germinación de esta especie en condiciones de luz. En otras especies de la familia Poaceae, como Setaria pallide-fusca (Schumach.) Stapf & C.E.Hubb. y Pennisetum pedicellatum Trin. también se registró a temperatura constante y con fotoperiodo de 12 horas, mayor porcentaje de germinación cuando el proceso de germinación se llevó a cabo con incidencia de luz (Afolayan y Olugbami, 1993).
De acuerdo con Bewley et al. (2013), la luz es un estímulo determinante para la germinación, debido a la cantidad de fotorreceptores que poseen las semillas. Esta característica induce a las semillas fotoblásticas positivas a presentar un mayor porcentaje de germinación en presencia de luz (Bewley et al., 2013; Merchán et al., 2020). Lo observado en la germinación de las tres especies estudiadas, similar a lo expresado, permite inferir que los cariopses de estas especies tienen una respuesta fotoblástica positiva.
La interacción de los factores iluminación y temperatura también tuvo efectos significativos en el TMG de las tres especies bajo estudio. En un trabajo realizado por Kloster et al. (2016), donde evaluaron la germinación de diferentes poblaciones de Trichloris crinita, obtuvieron un TMG de ocho días, un valor muy elevado con respecto a los observados en este trabajo, los cuales, bajo cualquiera de las condiciones aplicadas, ninguna de las especies superó un TMG de 2.41 días (en P. caespitosum con luz). En P. phillippianum y D. californica,Ferrari y Parera (2015) informaron una disminución del TMG en presencia de luz con respecto a la condición de oscuridad a medida que aumentó la temperatura (de 15 a 35 °C). Los valores bajos registrados en esta variable podrían ser debido al tipo de propágulo usado para los experimentos de germinación. En este estudio, los valores promedio usados para estimar las variables se obtuvieron a partir de cariopses desnudos, mientras que las investigaciones mencionadas trabajaron solamente con espiguillas, las cuales retrasan la velocidad de germinación.
Con respecto al efecto de la temperatura sobre la germinación, en L. crinita y D. californica, los valores medios evaluados (20 y 25 °C) fueron los que mostraron mayores porcentajes de germinación. En P. caespitosum fue solo a los 30 °C. Estos resultados concuerdan con los obtenidos por Di Giambatista et al. (2010), quienes evaluaron el efecto de la temperatura en la germinación de Trichloris crinita y Digitaria eriantha Steud., y encontraron el mayor valor de germinación en T. crinita a 25 °C y en D. eriantha a 30 °C. En Digitaria nuda Schumach. Hugo et al. (2014) también encontraron mayores porcentajes de germinación a temperaturas constantes de 25 y 30 °C (99%), cuando aplicaron un pretratamiento de remojo en agua por 24 horas.
Sartor y Marone (2010) evaluaron la germinación de diferentes pastos perennes y anuales a partir de espiguillas. Luego de aplicar tratamientos de calor y frío para romper en las semillas su latencia, encontraron porcentajes de germinación a 30 °C, del 99.4% en T. crinita, 76.4% en P. caespitosum y de 62% en D. californica. En el presente trabajo no se aplicaron tratamientos germinativos ya que se encontraron porcentajes de germinación elevados 24 horas después de la siembra, principalmente en los casos donde los experimentos se realizaron con los cariopses desnudos, en las tres especies.
La presencia de brácteas accesorias es un factor determinante para la germinación de los pastos. Las lemas y paleas funcionarían impidiendo el paso de luz hasta los cariopses, como barreras físicas para la emergencia de las radículas, o bien como proveedoras de inhibidores de la germinación (Oreja et al., 2017). Se ha informado, además, que las brácteas accesorias son una barrera hidrofóbica donde la lignina, polisacáridos y pectinas fenólicas contribuyen a determinar el carácter impermeable de la cubierta seminal e impiden la difusión de agua para la imbibición (Quero Carrillo et al., 2017).
Si bien no se han encontrado antecedentes sobre la influencia de la espiguilla en la germinación de las especies estudiadas en este trabajo, existen estudios sobre el papel de los restos florales en la eficiencia germinativa en otros miembros de la familia Poaceae. Oreja et al. (2017) encontraron diferencias significativas en la germinación de Digitaria sanguinalis al comparar los resultados entre semillas desnudas y el testigo realizado con la espiguilla y obtuvieron mayor germinación cuando el experimento se realizó con las semillas desnudas. En pastos introducidos en México (Cenchrus ciliaris L., cultivar Americano y Rhodes Chloris gayana Kunth, cultivar Bell), Quero Carrillo et al. (2017) obtuvieron mayor germinación en ambas especies al eliminar las brácteas accesorias.
Ma et al. (2008) evaluaron el efecto de palea y lema en la germinación de Leymus chinensis (Trin.) Tzvelev, y encontraron que la presencia de lemas es un factor determinante en el proceso de germinación de esa especie, y que estaría jugando un papel importante en el mantenimiento de la latencia del cariopse. También, Herrera-C. et al. (2006) estudiaron la germinación de cuatro ecotipos de Chloris cucullata Bisch. y cuatro de C. subdolichostachya Lag., a partir de espiguillas y de cariopses desnudos, y registraron una mayor germinación en todos los ecotipos en los tratamientos en los que se usaron cariopses desnudos.
Asimismo, Jensen y Boe (1991) encontraron que la germinación de Panicum virgatum L. aumentó luego de aplicar escarificación mecánica para eliminar lemas y paleas. Rost (1975) informó que Setaria lutescens F.T. Hubb. no germina en presencia de las glumas. En este estudio, de los tres factores abióticos evaluados (luz, tipo de propágulo y temperatura), la presencia de la espiguilla fue el factor que más negativamente influyó en el proceso de germinación. En L. crinita y P. caespitosum, la presencia de los restos florales produjo un descenso de alrededor de 48% en la germinación y en D. californica, la caída del porcentaje de germinación fue aproximadamente de 25%.
En condiciones naturales, los factores abióticos influyen simultáneamente en el proceso de germinación de las especies, por ello es importante analizar la interacción entre ellos. En las tres especies evaluadas, los porcentajes de germinación más elevados se alcanzaron cuando el proceso de germinación se llevó a cabo en presencia de luz, con el cariopse desnudo y con temperaturas entre 20 y 30 °C. Estas combinaciones de factores, principalmente la presencia de la espiguilla en el momento de la germinación, son muy importantes para tener en cuenta en los programas de recuperación de suelos degradados, ya que en el caso de realizar una siembra directa de cariopses de las especies evaluadas, sería recomendable eliminar las brácteas accesorias y utilizar cariopses desnudos. Además, en cuanto a las características agronómicas, la presencia de glumas, palea y lema, dificulta el manejo de semillas para la siembra, y también influye negativamente en la pureza físico-botánica de un lote de semillas en su análisis de calidad (Quero Carrillo et al., 2017).
En las zonas áridas de Argentina, Leptochloa crinita, Pappophorum caespitosum y Digitaria californica son las principales especies que componen el estrato herbáceo, las cuales podrían ser seleccionadas para su siembra en programas de recuperación de áreas degradadas, ya que entre las tres cubren un amplio rango de temperaturas óptimas de germinación, además de mostrar una respuesta positiva en la germinación con respecto a la incidencia de la luz.
Los resultados también sugieren que el proceso de germinación puede mejorarse eliminando las estructuras florales. Estudios sobre tolerancia a estrés hídrico y salino, y sobre los requerimientos de plántulas para el establecimiento bajo condiciones de estrés abiótico, constituyen un complemento importante a los resultados obtenidos en este trabajo a la hora de planificar acciones de restauración de ecosistemas degradados en zonas áridas.
Contribución de autores
JS llevó a cabo la recolección del material vegetal en el campo. CM y CP realizaron el diseño de los experimentos. CM efectúo los experimentos de germinación y los análisis estadísticos. CM y JS elaboraron la redacción inicial del manuscrito. CM, CP y JS revisaron y ajustaron la versión final del manuscrito en igualdad de contribución.

