












 (pdf)
(pdf)




 SciELO
SciELO  SciELO
SciELO 
 Permalink
Permalink
Introducción
La familia Cactaceae comprende un conjunto de arbustos y árboles, los cuales poseen una estructura morfológica derivada y compartida: la aréola (Anderson, 2001; Vázquez-Sánchez et al., 2012). Por otro lado, Nyffeler (2002), utilizando secuencias del cloroplasto (región trnK/matK e IGS trnL- trnF), ha corroborado su monofilia y además ha confirmado su presencia en el orden Caryophyllales (Cuénoud et al., 2002). El número de especies reconocidas por autores como Nyffeler y Eggli (2010) es de ca. 1850, un número muy próximo al mencionado por Korotkova et al. (2021) quienes reportan aproximadamente 1851 especies.
Uno de los géneros más representativo de Cactaceae es Mammillaria Haw., que agrupa plantas que se caracterizan por tener látex que puede ser acuoso, semilechoso a lechoso, tallos provistos de tubérculos con los meristemos espinífero y florífero separados, flores diurnas que emergen en torno al ápice y frutos por lo general carnosos (Bravo-Hollis y Sánchez-Mejorada, 1991; Lüthy, 1995; Arias yAquino, 2019). En Mammillaria se han reconocido aproximadamente 143 especies (Korotkova et al., 2021). Recientemente, las exploraciones botánicas han permitido descubrir nuevas especies (García-Morales et al., 2020; González-Zamora et al., 2023). Por otro lado, estudios sistemáticos enfocados en la delimitación de especies han permitido esclarecer límites entre taxones anteriormente reconocidos como una sola entidad; por lo tanto, el número de especies continúa incrementándose (González-Zamora et al., 2022; Cervantes et al., 2023; Ortiz-Brunel et al., 2023).
La clasificación infragenérica de Mammillaria comprende cinco subgéneros, tres secciones y 16 series (Hunt et al., 2006). Mammilaria ser. Leucocephalae (Lem.) Schum. incluye plantas con tallos simples a ramificados por brotes laterales o por división del ápice, presencia de cristales extracelulares, axilas de los tubérculos provistas de cerdas y tricomas, así como espinas diferenciadas en radiales y centrales, que generalmente son blancas con el ápice oscurecido. Solo una especie (M. muehlenpfordtii C.F. Först.) presenta espinas amarillas (Lüthy, 1995; Hunt et al., 2006). Estudios filogenéticos basados en el análisis de secuencias de cpDNA (intrón rpl16 e IGS psbA- trnH) sugieren que Mammillaria ser. Leucocephalae no es un grupo monofilético (Butterworth y Wallace, 2004). Posteriormente, Cervantes et al. (2021), utilizando también secuencias de cpDNA (intrón rpl16 e IGS psbA- trnH), recuperaron a los miembros de Mammillaria ser. Leucocephalae como un grupo monofilético. Cabe mencionar que esta disparidad en cuanto a resultados se debe a diferencias en el muestreo, la codificación de indels en el segundo trabajo y a los métodos de reconstrucción filogenética empleados. Sin embargo, en ambas hipótesis no es posible reconocer a su grupo hermano debido a la falta de resolución de los árboles filogenéticos. Debido a esto, no hay un consenso en cuanto al número de especies reconocidas. Lüthy (1995) incluyó a 10 especies; por su parte, Hunt et al. (2006) aceptaron 10 especies y cinco subespecies, en tanto que Korotkova et al. (2021) enlistaron ocho especies y seis subespecies. En el Bajío y regiones adyacentes se han registrado aproximadamente 70 especies y nueve subespecies de Mammillaria, de las cuales 14 son endémicas de la región; de estas, siete especies y una subespecie pertenecen a M. ser. Leucocephalae (Arias yAquino, 2019).
Durante la revisión de ejemplares de herbario de la familia Cactaceae para el proyecto “Flora del Bajío y de regiones adyacentes”, se encontraron ejemplares cuya presencia de cerdas entre las axilas de los tubérculos y el número de espinas radiales eran semejantes a Mammillaria hahniana Werderm. (Mammillaria ser. Leucocephalae). Uno de los ejemplares tenía adjuntadas un par de flores de las que, debido al estado de preservación, no fue posible determinar el color de los tépalos. Exploraciones posteriores en el límite este del Bajío en el municipio San Luis de la Paz, Guanajuato, así como observaciones sobre ejemplares colectados y mantenidos en cultivo en el Jardín Botánico de la Universidad Nacional Autónoma de México (UNAM), permitieron corroborar que el color de los tépalos no coincidía con las descripciones para Mammillaria hahniana (Bravo-Hollis y Sánchez-Mejorada, 1991; Reppenhagen, 1992; Arias yAquino, 2019). Por lo tanto, el objetivo del presente trabajo es caracterizar y describir una nueva especie para el Bajío.
Material y Métodos
Muestreo de taxones
El trabajo de campo se realizó en dos localidades del municipio San Luis de la Paz, Guanajuato, en México, en junio 2022. Adicionalmente, dada su cercanía morfológica y distribución simpátrica, fueron incluidos tres taxones de Mammillaria ser. Leucocephalae como grupo comparativo: M. hahniana, M. morganiana Tiegel y M. parkinsonii C. Ehrenb. Este conjunto de taxones se caracteriza por tener tubérculos hasta con cuatro espinas centrales, axilas con cerdas y flores con tépalos de color crema a rosado pálido, excepto Mammillaria hahniana cuyos tépalos son color magenta.
Se consultaron las colecciones de los herbarios FCME, IEB, MEXU, QMEX y TEX (Thiers, 2023+). A partir de ello se obtuvieron tres localidades para cada taxón; estas se ubican en los estados de Querétaro y Guanajuato (Apéndice). En campo se tomaron fotografías de diez ejemplares junto con una escala milimétrica en diferentes planos del tallo de la planta, se recolectaron flores, frutos y semillas. Los ejemplares de respaldo se depositaron en IBUG y MEXU (Thiers, 2023+).
Análisis estadístico
La selección de caracteres se hizo con base en la revisión de literatura especializada (Ehrenberg, 1840; Werdermann, 1929; Bravo-Hollis y Sánchez-Mejorada, 1991; Reppenhagen, 1992; Pilbeam, 1999; Arias yAquino, 2019). Inicialmente se evaluaron ocho caracteres cuantitativos (Cuadro 1) a partir de los cuales se calculó la media y desviación estándar.
Cuadro 1: Caracteres usados en el análisis morfológico y acrónimos correspondientes. El carácter IND fue calculado a partir de la división entre ALT y DIAM; el asterisco indica que los caracteres fueron descartados del análisis canónico discriminante.
| Carácter | Acrónimo |
|---|---|
| Altura del tallo (cm) | ALT* |
| Diámetro del tallo (cm) | DIAM* |
| División entre ALT/DIAM | IND |
| Largo de la espina central inferior (cm) | LIEC |
| Número de espinas centrales | NEC |
| Número de espinas radiales | NER |
| Largo de las espinas radiales (cm) | LIR |
| Largo de la aréola (cm) | ALEN |
| Ancho de la aréola (cm) | AWID |
En campo se tomaron fotografías de diez ejemplares; por cada ejemplar fotografiado se midieron o contaron diez estructuras y los valores obtenidos se organizaron en una base de datos. Para evitar redundancia entre los caracteres, se calculó el coeficiente de correlación de Pearson utilizando el paquete dplyr (Wickham et al., 2020). Se consideró que hay correlación entre caracteres cuando el valor absoluto del coeficiente sea igual o mayor a 0.7 (Dormann et al., 2013). Se realizó un análisis discriminante canónico (ACD) con el paquete CANDISC (Friendly y Fox, 2021). Los análisis estadísticos se realizaron en RStudio v. 1.1.447 (RStudio Team, 2015), ejecutado en R v. 3.5.0 (R Development Core Team, 2008).
Caracterización ecológica
A partir de las localidades obtenidas de los ejemplares de herbario se construyó una base de datos de georreferenciación. Aquellas localidades duplicadas e imprecisas fueron eliminadas de la base de datos, así como las que se encuentran a menos de 5 km entre sí con el fin de evitar una sobrerrepresentación y sobrestimación de resultados (Aquino et al., 2021; Cervantes et al., 2023). Para calcular la distancia entre localidades se utilizó Geographic Distance Matrix Generator v. 1.2.3 (Ersts, 2018). La base final incluyó siete registros de Mammillaria sp., once de M. hahniana, tres de M. morganiana y ocho de M. parkinsonii.
La caracterización ecológica incluyó dos variables: el tipo de suelo y geoforma (Fischer et al., 2008; Pineda et al., 2014). Cada capa tiene una resolución espacial de aproximadamente 1 km2. Las variables se extrajeron utilizando QGIS v. 2.1.8 (QGIS Development Team, 2009); de igual forma, se empleó el mismo software para la elaboración de un mapa. Se construyó una matriz de contingencia a partir de la cual se calcularon los residuos de Pearson y se aplicó la prueba de bondad de ajuste chi-cuadrada (Gómez-Gómez et al., 2003), se utilizó con el paquete dplyr (Wickham et al., 2020) y se graficó con corrplot (Wei y Simko, 2017), los cuales se calcularon en RStudio v. 1.1.447 (RStudio Team, 2015), ejecutado en R v. 3.5.0 (R Development Core Team, 2008).
Caracterización anatómica
Con el fin de corroborar la correcta inclusión de la nueva especie en Mammillaria ser. Leucocephalae se realizó la búsqueda de cristales extracelulares (cuticulares sensuLüthy, 1995). Adicionalmente, se incluyó una muestra de M. gigantea Hildm. ex K. Schum. (M. ser. Mammillaria), una especie geográficamente cercana al nuevo taxón y con cierta afinidad morfológica, con el fin de contrastar la presencia-ausencia de cristales (Lüthy, 1995).
El tejido correspondiente a Mammillaria sp. se obtuvo de un ejemplar colectado en campo (D. Aquino et al. 510, MEXU), mientras que la muestra de tejido de M. gigantea se obtuvo de un individuo depositado en la colección del Jardín Botánico del IBUNAM (S. Arias y D. Aquino 2256, MEXU). Se cortaron de dos a tres tubérculos de la parte media de tallos maduros, dejándolos reposar en alcohol etílico comercial al 96% por 12 horas. Posteriormente, se realizaron cortes a mano alzada de aproximadamente 1 mm de grosor en sección transversal y se montaron en preparaciones mostrando segmentos de la cutícula y epidermis. Las preparaciones fueron observadas en un fotomicroscopio Axioskop (Carl Zeiss, Jena, Alemania), usando las técnicas de iluminación campo claro y polarización. Se buscaron cristales extracelulares y una vez localizadas dichas estructuras se tomaron fotomicrografías con una cámara de video ExwaveHAD (SONY, Tokio, Japón) y posteriormente se editaron con RisingView v. x64 (Softs Ton, 2019).
Conservación
Se estimaron el área de ocupación (AOO) y la extensión de ocurrencia (EOO) de la nueva especie propuesta mediante el uso de la herramienta de evaluación de conservación geoespacial GeoCAT (Bachman et al., 2011).
Resultados
Caracterización morfológica
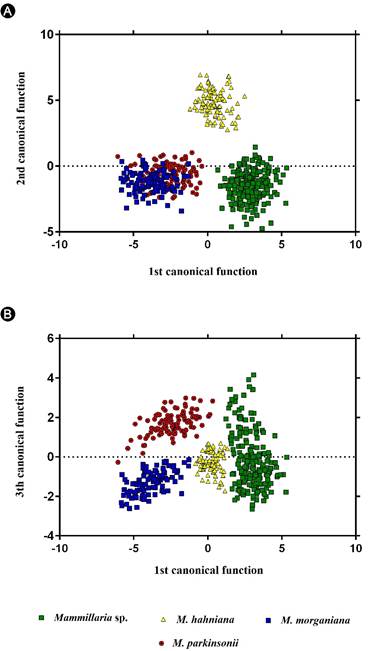
El Cuadro 2 corresponde a los valores de media y tendencia central de los ocho caracteres cuantitativos. El valor del coeficiente de correlación de Pearson entre ALT y DIAM indicó que hay correlación entre las variables: 0.735634 (Cuadro 3), por lo que se calculó la relación ALT/DIAM de tallo (IND) para retener la información obtenida de las dimensiones del tallo. Para este nuevo carácter se calculó la media y desviación estándar (Cuadro 2) y fue incluido en el análisis canónico discriminante (ACD), en conjunto con los seis caracteres restantes cuyo índice de correlación fue bajo (Cuadro 3). En el ACD, la primera función explicó 52.28% de la variación y el carácter con mayor peso fue NER. La segunda función canónica explicó 41.37% de la variación y ALEN resultó como el carácter de mayor peso (Cuadro 4). Mammillaria sp. se agrupó en el extremo derecho de la primera función canónica, en donde el rango de NER es de (18)19-24(26) espinas, mientras que en el extremo izquierdo fueron discriminados M. morganiana y M. parkinsonii, cuyos valores de NER son mayores a (24)25 espinas (Fig. 1A). Sobre el extremo superior de la segunda función canónica fueron segregados los individuos determinados a priori como M. hahniana cuyo rango de ALEN es de (0.09)0.11-0.14(0.16) cm (Fig. 1A). En el extremo inferior se segregan Mammillaria sp., M. morganiana y M. parkinsonii, cuyos valores de ALEN son >0.17 cm. La tercera función canónica explicó 6.35% de la variación y mostró que el carácter de mayor peso fue LIEC (Fig. 1B; Cuadro 2). Sobre el extremo superior de la tercera función canónica se discriminaron los individuos determinados como M. parkinsonii, cuyos valores de LIEC son de (1.62)1.74-2(2.1) cm (Fig. 1B). En tanto que los individuos determinados como Mammillaria sp. se distribuyeron a lo largo del eje de la tercera función canónica, pues presentan un amplio rango de variación: (0.5)0.65-1.5(2) cm. El rango de LIEC es de (0.38)0.43-0.55(0.65) cm para Mammillaria hahniana y de (1.21)1.32-1.40(1.55) cm para M. morganiana. Por lo tanto, se sitúan en el extremo de los valores negativos de la tercera función canónica.
Cuadro 2: Media (M) y desviación estándar (ds) calculada para cuatro especies de Mammillaria ser. Leucocephalae. Altura del tallo=ALT, Diámetro del tallo=DIAM, División entre ALT/DIAM=IND, Largo de la espina central inferior=LIEC, Número de espinas centrales=NEC, Número de espinas radiales=NER, Largo de espinas radiales=LIR, Largo de la aréola=ALEN, Ancho de la aréola=AWID. El asterisco indica que fueron descartados del análisis canónico discriminante. Especies: 1=Mammillaria sp., 2=Mammillaria hahniana Werderm., 3=Mammillaria morganiana Tiegel, 4=Mammillaria parkinsonii Ehrenb. *=variables no incluidas en el análisis canónico discriminante por correlación positiva entre ellas.
| Especie | ALT* | DIAM* | IND | NEC | LIEC | NER | LERA | ALEN | AWID | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | M | ds | M | ds | M | ds | M | ds | M | ds | M | ds | M | ds | M | ds | M | ds | |
| 1 | 20 | 8.58 | 1.497 | 11.56 | 2.368 | 1.37 | 0.290 | 5.58 | 1.038 | 1.108 | 0.434 | 21.69 | 2.143 | 0.72 | 0.089 | 0.217 | 0.022 | 0.136 | 0.107 |
| 2 | 10 | 8.06 | 1.371 | 9.35 | 2.141 | 1.159 | 0.166 | 2.65 | 0.925 | 0.551 | 0.139 | 26.86 | 1.869 | 0.581 | 0.088 | 0.124 | 0.016 | 0.053 | 0.011 |
| 3 | 10 | 6.77 | 1.146 | 7.91 | 0.970 | 1.181 | 0.124 | 3.39 | 0.941 | 1.332 | 0.066 | 37.41 | 3.607 | 0.856 | 0.088 | 0.206 | 0.016 | 0.111 | 0.014 |
| 4 | 10 | 5.91 | 0.848 | 6.43 | 1.108 | 1.096 | 0.171 | 3.19 | 0.981 | 1.881 | 0.124 | 31.82 | 3.761 | 0.666 | 0.058 | 0.189 | 0.021 | 0.107 | 0.095 |
Cuadro 3: Coeficientes de Correlación de Pearson (PCC) calculado para ocho variables morfológicas. Altura del tallo=ALT, Diámetro del tallo=DIAM, Largo de la espina central inferior=LIEC, Número de espinas centrales=NEC, Número de espinas radiales=NER, Largo de espinas radiales=LIR, Largo de la aréola=ALEN, Ancho de la aréola=AWID. *=variables no incluidas en el análisis canónico discriminante por correlación positiva entre ellas.
| ALT* | DIAM* | NEC | LIEC | NER | LERA | ALEN | AWID | |
|---|---|---|---|---|---|---|---|---|
| ALT | 1 | |||||||
| DIAM | 0.735634 | 1 | ||||||
| NEC | 0.338828 | 0.465574 | 1 | |||||
| LIEC | -0.4872 | -0.44767 | -0.02963 | 1 | ||||
| NER | -0.49535 | -0.57026 | -0.48628 | 0.322782 | 1 | |||
| LERA | 0.003097 | 0.132629 | 0.181458 | 0.117306 | 0.308323 | 1 | ||
| ALEN | 0.020096 | 0.202334 | 0.500581 | 0.388125 | -0.0861 | 0.475929 | 1 | |
| AWID | 0.037347 | 0.109246 | 0.269563 | 0.148309 | -0.1124 | 0.186181 | 0.314749 | 1 |
Cuadro 4: Coeficientes de estructura canónica para siete caracteres cuantitativos. Altura del tallo=ALT, Diámetro del tallo=DIAM, División entre ALT/DIAM=IND, Largo de la espina central inferior=LIEC, Número de espinas centrales=NEC, Número de espinas radiales=NER, Largo de espinas radiales=LIR, Largo de la aréola=ALEN, Ancho de la aréola=AWID. Los caracteres con mayor peso por cada función se remarcan en negritas.
| Can1 | Can2 | Can3 | |
|---|---|---|---|
| AWID | 0.003333 | -0.07311 | -0.00683 |
| IND | 0.215688 | 0.019231 | -0.05199 |
| ALEN | 0.121661 | -0.7109 | -0.17284 |
| LIR | -0.29973 | -0.42538 | -0.40253 |
| LIEC | -0.36327 | -0.53472 | 0.719958 |
| NEC | 0.44663 | -0.37854 | -0.08967 |
| NER | -0.83859 | 0.01369 | -0.25159 |
| % | 52.28 | 41.37 | 6.35 |
Caracterización ecológica
La asociación entre el tipo de suelo y Mammillaria sp. fue significativo para el tipo de geoforma (chi-cuadrada p=0.008966, Fig. 2A). De acuerdo con el análisis, Mammillaria sp. se establece con mayor frecuencia sobre relieves volcánicos (r de Pearson=3.316); a su vez, hay asociación negativa por sistemas de piedemonte (r de Pearson=-1.474). Los taxones con los cuales se contrastó Mammillaria sp. también mostraron asociación por un tipo particular de geoforma. Mammillaria hahniana tiene una asociación moderada en sistemas kársticos (r de Pearson=1.204). Un patrón semejante ocurre en Mammillaria morganiana donde la asociación por sistemas fluviales es moderada (r de Pearson=1.108). Mammillaria parkinsonii, por el contrario, es frecuente en sistemas de piedemonte (r de Pearson=2.232).
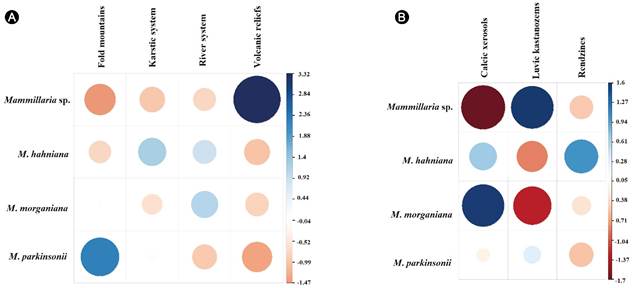
Figura 2: Correlogramas entre A. Geoforma; B. Tipo de suelo, calculados para cuatro especies de Mammillaria Haw. ser. Leucocephalae (Lem.) Schum. Las correlaciones positivas se muestran en colores azules y las negativas en colores rojos. Los residuos de Pearson se muestran en el lado derecho de cada gráfico.
La prueba de bondad de ajuste fue significativa para el tipo de suelo (chi-cuadrada p=0.008966, Fig. 2B). Mammillaria sp. se establece en suelos tipo kastanozems lúvicos (R de Pearson=1.597); a su vez, existe asociación negativa por xerosoles cálcicos (r de Pearson=-1.702). Mammillaria morganiana por su parte se establece con mayor frecuencia en xerosoles cálcicos (r de Pearson=1.578) y la asociación negativa se da con los suelos kastanozems lúvicos (r de Pearson=-1.287). En el caso de Mammillaria hahniana se detectó frecuencia positiva por rendzinas (r de Pearson=1.008) y frecuencia negativa por kastanozems lúvicos (r de Pearson=-0.84). Finalmente, Mammillaria parkinsonii no muestra preferencias por un tipo particular de suelo. Con base en evidencias morfológicas y ambientales, proponemos que Mammillaria sp. sea reconocido como un nuevo taxón.
Caracterización anatómica
La Figura 3A muestra una sección transversal del tubérculo de Mammillaria sp. El tejido está organizado en una cutícula gruesa al momento de cortar. Enseguida se observa la epidermis y finalmente la hipodermis cuyas paredes celulares muy engrosadas son fuertemente refringentes. Entre la cutícula y la pared periclinal de la epidermis se observaron cristales, cuya composición no es posible determinar por el momento. En contraste, en la Figura 3B, en sección transversal, el arreglo celular de Mammillaria gigantea indica la presencia de cutícula que al momento del corte no hubo resistencia. Se observa la epidermis seguido de la hipodermis cuyas paredes celulares no son engrosadas; por lo tanto, no son refringentes. Se corroboró la ausencia de cristales entre la cutícula y la epidermis en esta especie. Este resultado permite proponer a Mammillaria sp. como miembro de M. ser. Leucocephalae.
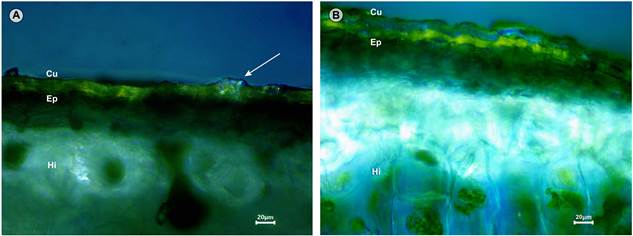
Figura 3: Sección transversal de tubérculos vistos con luz polarizada para A. Mammillaria ariasii U. Guzmán & D. Aquino, sp. nov. (D. Aquino et al. 510 (MEXU)); B. Mammillaria gigantea Hildm. ex K. Schum. (S. Arias y D. Aquino 2256 (MEXU)). Cu: cutícula, Ep: epidermis, Hi: hipodermis. Se marca con una flecha la presencia de cristales extracelulares. Fotografías de E. Sandoval-Zapotitla y D. Aquino.
Taxonomía
Mammillaria ariasii U. Guzmán & D. Aquino, sp. nov. Fig. 4.

Figura 4: Mammillaria ariasii U. Guzmán & D. Aquino, sp. nov. (D. Aquino et al. 510 (MEXU)). A-C. vista general del tallo; D. disposición y variación de las espinas centrales y radiales; E. flor; F. frutos; G. semillas. Fotografías de D. Aquino.
TIPO: MÉXICO. Guanajuato, municipio San Luis de la Paz, Sierra Gorda, 1333 m, bosque de Quercus, 27.VI.2022, D. Aquino et al. 510 (holotipo: MEXU!, isotipo: IBUG!).
Mammillaria ariasii is distinguished from M. hahniana, M. morganiana and M. parkinsonii by having (18-)19-24(-26) radial spines, while the three taxa referred to have more than 30 radial spines. Mammillaria ariasii has simple stems and rarely branches; when it does it is through lateral branching. In contrast, M. morganiana and M. parkinsonii, when branching, do so by division of the apex. Finally, Mammillaria ariasii differs from M. hahniana by the yellow color of the inner tepals, contrasting with the magenta tepals observed in M. hahniana. Additionally, the number of central spines differs, Mammillaria ariasii has (4-)5-7(-8) central spines, while M. hahniana, M. morganiana and M. parkinsonii have only 2 to 4 central spines; M. morganiana occasionally has 5 central spines.
Planta con tallo simple, ocasionalmente ramificada por brotes laterales; tallo hasta 9.1(-11.5) cm de alto, 13.9(-15.4) cm de diámetro, globosas a ligeramente deprimido-globosas, látex de consistencia lechosa, cristales extracelulares presentes, tubérculos 1.3-1.5(-1.65) cm × 0.7-1(-1.2) cm, ca. 4-angulados, axilas provistas con cerdas y tricomas de longitud y densidad variable, blancas, aréolas (0.17-)0.19-0.23(-0.26) cm × (0.08-)0.12-0.15(-0.16) cm, ovaladas, espinas radiales (18-)19-24(-26), (0.47-)0.63-0.77(-0.90) cm de largo, aciculares, blancas, espinas centrales (4-)5-7(-8), la inferior (0.5-)0.65-1.5(-2) cm de largo, rectas a ligeramente curvas, no uncinadas, de color rojo en tonos oscuros a castaño claro con el ápice oscurecido; flor (1.55-)1.64-1.75(-1.85) cm de largo; tubo receptacular reducido; tépalos externos hasta 0.4 cm de largo, lanceolados, agudos, castaño-rojizo con los bordes amarillos, tépalos internos hasta 0.7(-0.8) cm de largo, lanceolados, acuminados, amarillos, filamentos amarillos, anteras amarillas, estilo verde, estigma con 3 a 5 lóbulos, verdes; fruto (1.4-)1.5-2 cm × 0.5-0.8(-0.9) cm, rosados, claviformes, conservando los restos secos del perianto; semillas ca. 1.5 mm × ca. 1 mm, castaño oscuro.
Distribución y hábitat: Mammillaria ariasii es conocida de la Sierra Gorda de Guanajuato, México, en el extremo norte de la región del Bajío (Fig. 5). Se establece en bosque de Quercus y bosque tropical caducifolio, sobre sitios con pendientes. Ocupa un rango de altitud de 920 a 2000 metros. Cohabita con Quercus spp., Isolatocereus dumortieri (Scheidw.) Backeb., Stenocactus dichroacanthus A. Berger ex Backeb. & F.M. Knuth, Stenocereus huastecorum Alvarado-Sizzo, Arreola-Nava & Terrazas, Agave xylonacantha Salm-Dyck. y Jatropha dioica Sessé. El cálculo de los residuos de Pearson indica que Mammillaria ariasii tiene una fuerte preferencia por suelos tipo kastanozems lúvicos, los cuales se caracterizan por presentar hasta 8% de arcilla y sin estructura de roca (FAO, 2014). De acuerdo con nuestros resultados, Mammillaria ariasii se distribuye frecuentemente sobre relieves volcánicos, al mismo tiempo que evade los sistemas de piedemonte.
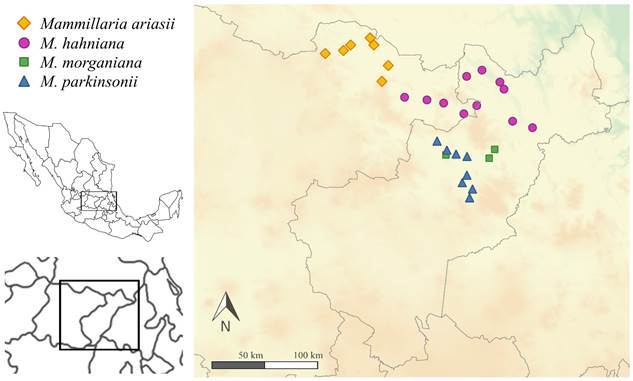
Figura 5: Distribución geográfica de Mammillaria ariasii U. Guzmán & D. Aquino y miembros de M. ser. Leucocephalae (Lem.) Schum. incluidos en el análisis comparativo.
Fenología: la floración inicia a mediados de octubre y finaliza a principios de diciembre; la apertura de la flor es completamente diurna. La fructificación ocurre al siguiente año entre principios de junio y mediados de septiembre. Se desconoce si los frutos son consumidos por los pobladores.
Etimología: el epíteto específico “ariasii” está dedicado al profesor Salvador Arias, estudioso de los cactus mexicanos, amigo y mentor de los autores de este trabajo.
Estado de conservación: se conocen por lo menos siete localidades con una población estimada de 300 plantas adultas y 100 plantas juveniles (criterio C1), ocupando una extensión de ocurrencia (EOO) de 345 km2 (criterio B1) y un área de ocupación (AOO) de 28 km2 (criterio B2). Por lo tanto, Mammillaria ariasii debe ser considerada como especie En Peligro (EN) de acuerdo con la clasificación de la Lista Roja de la Unión Internacional para la Conservación de la Naturaleza (IUCN, 2022). Según los criterios enlistados en la NOM-059-SEMARNAT-2010 (SEMARNAT, 2010), Mammillaria ariasii puede ser considerada dentro de la categoría Pr (sujeta a protección especial). Si bien se observó que el cambio de uso de suelo para la ganadería provoca erosión y, en consecuencia, arrastre de plantas, estos podrían ser factores a futuro que incidan negativamente sobre la viabilidad de las localidades. Sin embargo, no existen evidencias tempranas de recolecta ilegal. Se debe considerar que Mammillaria ariasii es un taxón con requerimientos ambientales específicos, por lo que se necesitan estudios enfocados en dinámicas poblacionales.
Especímenes adicionales examinados: MÉXICO. Guanajuato, municipio San Luis de la Paz, Sierra Gorda II, 22.VI.2022, D. Aquino et al. 514 (MEXU); ca. 37 km al NE de San Luis de la Paz, sobre el camino a San Antón de los Martínez, sobre el camino a El Vergel, 06.X.1995, R. T. Bárcenas y C. Gómez-Hinostrosa 664 (MEXU); ca. 55 km al NE de San Luis de la Paz, sobre el camino de Mineral del Realito, la desviación a Vergel y Mineral del Realito está a 1 km al NE de Mesa de Jesús, 06.X.1995, R. T. Bárcenas y C. Gómez-Hinostrosa 673 (MEXU); ca. 59 km al NE de San Luis de la Paz sobre el camino a Mineral del Realito, la desviación a El Vergel, Cerro de Bernalejo y Mineral del Realito está a 1 km de Mesa de Jesús, 07.X.1995, R. T. Bárcenas y C. Gómez-Hinostrosa 687 (MEXU); La Teresa, ca. 66 km al NE de San Luis de la Paz, sobre el camino a Mineral del Realito, la desviación a La Teresa está a 1 km al NE de Mesa de Jesús, 07.X.1995, R. T. Bárcenas y C. Gómez-Hinostrosa 692 (MEXU); cerros al N de La Teresa, ca 66 km al NE de San Luis de la Paz, sobre el camino a Mineral del Realito, la desviación a La Teresa está a 1 km al NE de Mesa de Jesús, 07.X.1995, R. T. Bárcenas y C. Gómez-Hinostrosa 702 (IEB, MEXU). Municipio Victoria, ca. 81 km desde Cañada de Moreno sobre la terracería a Mineral del Refugio, la desviación a Mineral del Refugio está a 28 km de Cañada de Moreno sobre la terracería a Xichú, 12.III.1995, R. T. Bárcenas y C. Gómez-Hinostrosa 490 (MEXU); ca. 500 m de la desviación hacia El Platanal, sobre el camino a Álamos de Martínez, la desviación a Álamos de Martínez está a 28 km de Cañada de Moreno sobre el camino a Xichú, 18.XI.1995, R. T. Bárcenas et al. 821 (MEXU); ca. 1.5 km al NO de Álamos de Martínez, sobre el camino a Mineral del Realito, 19.XI.1995, R. T. Bárcenas 883 (MEXU); ±7 km al NE de Joya Fría, 20.XI.2009, S. Zamudio 14671 (IEB).
Discusión
Con el empleo del análisis discriminante canónico es posible detectar cuáles son las mediciones lineales que más contribuyen a la distinción entre grupos e informa sobre la estructura de la varianza entre y dentro de grupos (De Luna, 2020). Los análisis morfométricos en géneros morfológicamente variables como Echinocereus Engelm. o Mammillaria no solo han permitido delimitar especies, sino también documentar la variación morfológica dentro de las especies, la propuesta de taxones a nivel infraespecífico, la dispersión en el morfoespacio de los especímenes muestreados, así como las descripciones con rangos de variación en los caracteres cuantitativos así lo demuestran (Sánchez et al., 2020; González-Zamora et al., 2022).
El uso de los análisis morfométricos para la descripción de especies nuevas es relevante, pues los caracteres diagnósticos cobran mayor relevancia cuando son estadísticamente significativos (Texeira et al., 2013). Los pocos estudios que han abordado la delimitación de especies de Mammillaria a través del análisis de secuencias de ADN han corroborado gran parte de las especies propuestas tradicionalmente. En uno de ellos, el análisis de cuatro secuencias de cpDNA corrobora la delimitación de cinco de nueve especies de Mammillaria ser. Polyedrae K. Schum., que son especies previamente propuestas mediante un análisis morfométrico (Aquino, 2014).
Por otro parte, la delimitación de especies mediante el análisis genómico, morfométrico y ecológico del complejo Mammillaria haageana Pffeif. permitió el reconocimiento de seis especies distintas (Cervantes et al., 2023). Este resultado contrasta con la propuesta de considerar a M. haageana como una especie con una variación amplia y continua (Hunt et al., 2006; Korotkova et al., 2021). Si bien el presente trabajo no presenta un análisis de secuencias de ADN, se muestra evidencia morfológica y ecológica para reconocer a una nueva especie y representa una hipótesis robusta que puede ser evaluada mediante caracteres moleculares.
De esta forma, una combinación de caracteres morfológicos permitió diferenciar a Mammillaria ariasii con respecto a otros miembros de M. ser. Leucocephalae (Fig. 6, Cuadro 5). Mammillaria ariasii es el taxón con el menor rango de espinas radiales: (18-)19-24(-26), mientras que en M. hahniana, M. morganiana y M. parkinsonii el número de espinas radiales es, en promedio, mayor a 25 espinas, alcanzando hasta 36 espinas en Mammillaria hahniana, o igual o más de 40 espinas radiales en M. morganiana y M. parkinsonii. Adicionalmente, la longitud de la aréola (ALEN) permitió discriminar a Mammillaria ariasii y M. hahniana, ya que esta última presentó la aréola más corta entre los taxones comparados, (0.09-)0.11-0.14(-0.16) cm. Por otra parte, la longitud de la espina central inferior (LIEC) presentó un amplio rango de variación en Mammillaria ariasii ((0.5-)0.65-1.5(-2) cm) y solo permitió discriminar entre Mammillaria parkinsonii ((1.62-)1.74-2(-2.1) cm) y M. morganiana ((1.21-)1.32-1.40(-1.55) cm). Los tres caracteres con mayor poder discriminante recuperados por el análisis canónico han sido reconocidos en otros análisis morfométricos realizados en taxones de la tribu Cacteae (Aquino et al., 2019; González-Zamora et al., 2022; Ortíz-Brunel et al., 2023). El número de espinas centrales (NEC) no fue recuperado como un carácter con carga discriminante por el análisis, a pesar de que Mammillaria ariasii tiene un rango de (4-)5-7(-8), lo cual contrasta con el rango aquí registrado para M. hahniana, M. morganiana y M. parkinsonii que es de 2-4, llegando hasta cinco en esta última. En otros estudios, el número de espinas centrales ha sido recuperado como un carácter útil para reconocer especies (Sánchez et al., 2020; González-Zamora et al., 2022). Adicionalmente, otros caracteres cualitativos permiten corroborar los límites de los taxones comparados en este estudio (Cuadro 3).

Figura 6: Comparación morfológica entre cuatro especies de Mammillaria ser. Leucocephalae (Lem.) Schum. A-C. Mammillaria ariasii U. Guzmán & D. Aquino (D. Aquino et al. 510 (MEXU)); D-F. Mammillaria hahniana Werderm. (D. Aquino et al. 522 (MEXU)); G-I. Mammillaria morganiana Tiegel (D. Aquino et al. 523 (MEXU)); J-L. Mammillaria parkinsonii Ehrenb. (D. Aquino et al. 525 (MEXU)). Fotografías de D. Aquino.
Cuadro 5: Comparación morfológica entre Mammillaria ariasii U. Guzmán & D. Aquino con respecto a tres especies pertenecientes a M. ser. Leucocephalae correspondiente al grupo funcional.
| Mammillaria ariasii U.Guzmán & D. Aquino | Mammillaria hahniana Werderm. | Mammillaria morganiana Tiegel | Mammillaria parkinsonii C. Ehrenb. | |
|---|---|---|---|---|
| Altura del tallo (cm) | (5.7-)7.4-9.1(-11.5) | (5.1-)7.1-9.5 | 5.7-7.6(-9.1) | (4.5-)5.5-6.4(-7.7) |
| Diámetro del tallo (cm) | (6.8-)10.3-13.9(-15.4) | (5.4-)8.2-10.2(-12.1) | (6.4-)7.2-8.7(-9.2) | (4.4-)6.1-7.2(-8.4) |
| Ramificación | simple | simple | dicotómica | dicotómica |
| Número de espinas centrales | (4-)5-7(-8) | 2-4 | 2-4(-5) | 2-4 |
| Largo de la espina central inferior (cm) | (0.5-)0.65-1.5(-2.0) | (0.38-)0.43-0.55(-0.65) | (1.21-)1.32-1.40(-1.55) | (1.62-)1.74-2.0(-2.1) |
| Número de espinas radiales | (18-)19-24(-26) | (24-)25-28(-30) | (31-)35-40(-45) | (23-)30-35(-40) |
| Largo de las espinas radiales (cm) | (0.47-)0.63-0.77(-0.90) | (0.42-)0.55-0.64(-0.79) | (0.71-)0.79-0.96(-1.03) | (0.52-)0.62-0.74(-0.81) |
| Largo de la aréola (cm) | (0.17-)0.19-0.23(-0.26) | (0.09-)0.11-0.14(-0.16) | (0.17-)0.19-0.22(-0.25) | (0.15-)0.17-0.23(-0.25) |
| Ancho de la aréola (cm) | (0.08-)0.12-0.15(-0.16) | (0.035-)0.04-0.06(-1.0) | (0.08-)0.1-0.12(-0.14) | (0.06-)0.071-0.12(-0.7) |
| Color de los tépalos internos | amarillo | magenta | crema a rosado pálido | crema a rosado pálido |
| Tipo de suelo | kastanozems lúvico | redzinas | xerosol cálcico | sin preferencia |
| Geoforma | relieve volcánico | sistema kárstico | sistema fluvial | piedemonte |
| Tipo de vegetación | bosque de Quercus y bosque tropical caducifolio | bosque tropical caducifolio | matorral xerófilo | matorral xerófilo |
| Distribución | Guanajuato | Guanajuato y Querétaro | Querétaro | Querétaro |
Las descripciones originales de Mammillaria parkinsonii y M. morganiana consideran a la ramificación del tallo como un carácter diagnóstico, pues en ambos taxones el ápice se divide en dos formando nuevas ramas (Ehrenberg, 1840; Reppenhagen, 1992; Arias yAquino, 2019), mientras que en Mammillaria ariasii y M. hahniana no hay ramificación o cuando ocurre se lleva a cabo por brotes basales laterales (Cuadro 3). Por otra parte, la descripción original de Mammillaria hahniana no incluyó la descripción de las flores (Werdermann, 1929); sin embargo, descripciones posteriores (Bravo-Hollis y Sánchez-Mejorada, 1991; Reppenhagen, 1992) y nuestras observaciones indican que los tépalos internos son de color magenta. Pilbeam (1999) cita un ejemplar cultivado en Cante A.C. como una forma de Mammillaria hahniana de espinas oscuras, flores amarillentas y cuyo sitio de procedencia se desconoce.
No es posible llegar a una conclusión sobre la identidad de este morfotipo, pues al observar la figura 146 de la obra de Pilbeam (1999) el individuo posee hasta cuatro espinas centrales. De acuerdo con nuestro análisis, Mammillaria ariasii posee (4-)5-7(-8), por lo que el rango de variación no concuerda con lo aquí reportado. Por lo tanto, no es posible determinar la identidad de los individuos depositados en Cante A.C. en tanto no se disponga de información de procedencia. Mammillaria morganiana y M. parkinsonii muestran la base de los tépalos de color rosado, mientras que el tercio superior es crema. En contraparte, los tépalos internos de Mammillaria ariasii son en su totalidad de color amarillo. En este mismo sentido, el color de los filamentos y la antera en Mammillaria ariasii es amarillo, mientras que en el grupo comparativo las flores mostraron filamentos y anteras rosadas (Fig. 6).
Otros miembros de Mammillaria ser. Leucocephalae distribuidas en el Bajío poseen rangos semejantes en el número de espinas centrales: M. sempervivi DC. 3-4 (De Candolle, 1828), M. formosa Galeotti ex Scheidw. subsp. pseudocrucigera (R.T. Craig) D.R. Hunt (2-)4(-6) (Craig, 1945) y M. perbella Hildm. ex K. Schum. 1-2 (Schumann, 1898), mientras que M. geminispina ha sido descrita con 2-4 espinas (Bravo-Hollis y Sánchez-Mejorada, 1991; Reppenhagen, 1992; Arias yAquino, 2019). Este grupo de especies se distingue de Mammillaria ariasii por presentar tépalos internos rosados, rojo carmín a magenta en su totalidad, excepto M. sempervivi cuyos tépalos son blancos a crema en la parte inferior.
Finalmente, la descripción de Mammillaria muehlenpfordtii indica que posee ca. cuatro espinas centrales (Förster, 1847), pero en descripciones recientes se reportan de cuatro a seis espinas (Reppenhagen, 1992; Arias yAquino, 2019). Esta especie se caracteriza además por ramificar por división del ápice, por tener 30 a 45 espinas radiales y por tener flores con tépalos internos rosados.
Nuestro análisis mostró que la distribución geográfica de Mammillaria ariasii se ve influenciada fuertemente por el tipo de suelo. Es evidente la fuerte evasión de Mammillaria ariasii por xerosol cálcico. Por el contrario M. hahniana y sobre todo M. morganiana evaden los suelos kastanozems lúvicos y prefieren xerosol cálcico y rendzinas. Únicamente M. parkinsonii no exhibe preferencias por un tipo particular de suelo. Fenómenos similares se han observado en los miembros que integran a Epithelantha F.A.C. Weber ex Britton & Rose y Pilosocereus Byles & G.D. Rowley, ya que este mecanismo permitió la divergencia entre especies, a su vez de fungir como barreras entre las mismas (Aquino et al., 2021; Franco-Estrada et al., 2022). Mammillaria parkinsonii, por el contrario, es frecuente en sistemas de piedemonte y evade los relieves volcánicos, mientras que M. hahniana y M. morganiana presentan frecuencias moderadas por sistemas kársticos y fluviales, respectivamente. Considerando que el flujo de materia y agua genera microambientes, la distribución de las comunidades vegetales sobre una geoforma no es al azar y con ello se promueve el aislamiento de linajes (Schaetzl y Anderson, 2005; Aquino et al., 2021).
Finalmente, Mammillaria ariasii es superficialmente parecida a M. gigantea, ambos taxones poseen látex lechoso característica de los miembros de M. sect. Mammillaria; además, los tallos son simples y los tépalos internos son de color amarillo. Mammillaria gigantea se caracteriza por tener tallos de hasta 14 cm de alto, por la ausencia de cerdas entre las axilas de los tubérculos y por presentar ca. 12 espinas radiales (Schumann, 1898; Arias yAquino, 2019). En contraparte, la altura de los tallos de Mammillaria ariasii es de hasta 9.1(-11.5) cm, presenta cerdas entre las axilas de los tubérculos y posee (18-)19-24(-26) espinas radiales. Las axilas de los tubérculos de Mammillaria ariasii están provistas de cerdas y tricomas. Además, nuestros resultados corroboran la presencia de cristales extracelulares (Fig. 3A), los cuales están presentes en los miembros de Mammillaria ser. Leucocephalae (Lüthy, 1995).
Por otro lado, Mammillaria gigantea es incluida en M. ser. Mammillaria, en donde los miembros de esta sección se caracterizan por la presencia de tricomas entre los tubérculos (cerdas presentes solo en M. compressa DC.) y ausencia de cristales extracelulares (Fig. 1B). Lüthy (1995) denominó a los biominerales presentes en Mammillaria ser. Leucocephalae como cristales cuticulares. Sin embargo, De la Rosa-Tilapa (2020) nombró a estas estructuras cristales extracelulares, puesto que se depositan entre las paredes periclinales de la epidermis y la cutícula (Fig. 1A). Desde el punto de vista de la sistemática de Cactaceae, la presencia de biominerales ha sido importante para soportar la monofilia de Turbinicarpus (Backeb.) Buxb. & Backeb., Rapicactus Buxb. & Oehme y Kadenicarpus Doweld, en donde la presencia de un tipo de cristal en la hipodermis es específica para cada género (De la Rosa-Tilapa et al., 2018; Vázquez-Sánchez et al., 2019). De la misma forma, el reconocimiento de Kimnachia S. Arias & N. Korotkova es soportado por la presencia de drusas, con respecto a Pseudorhipsalis Britton & Rose, género que se caracteriza por la presencia de cristales cúbicos (Martínez-Quezada et al., 2020).
A continuación, se proporciona una clave de identificación para las especies de Mammillaria ser. Leucocephalae distribuidos en el Bajío, incluyendo a Mammillaria ariasii.
Clave de identificación para las especies de Mammillaria ser. Leucocephalae distribuidas en el Bajío (México)
1a. Tallos al principio simples, posteriormente ramificados por división del ápice ................. 2
1b. Tallos simples o ramificados por brotes laterales ………………………...………………. 5
2a. Espinas centrales amarillas ………………..………………. M. muehlenpfordtii C.F. Först.
2b. Espinas centrales blancas a grises, en ocasiones con el ápice oscurecido …………..……. 3
3a. Espinas radiales (12-)14-18(-22), flores con los tépalos internos rojo carmín ………….……………………………………......................…… M. perbella Hildm. ex K. Schum.
3b. Espinas radiales >30, flores con los tépalos internos de color crema a rosado pálido …..... 4
4a. Número de espinas radiales (31-)35-40(-45), espina central inferior hasta 1.40(-1.55) cm de largo …………………………………………………….......…………...…. M. morganiana Tiegel
4b. Número de espinas radiales (23-)30-35(-40), espina central inferior hasta 2(-2.1) cm de largo ……………………………………………………............………… M. parkinsonii Ehrenb.
5a. Tallos depreso-globosos, espinas radiales presentes en plantas juveniles, ausentes en plantas adultas …………………………………………….………………………………..…… 6
5b. Tallos globosos, espinas radiales presentes tanto en plantas juveniles como adultas …… 7
6a. Aréolas circulares a ovaladas, flores hasta 1 cm de ancho, tépalos internos blancos …………………………………………………………………………….....…. M. sempervivi DC.
6b. Aréolas ovadas, flores hasta 1.8 cm de ancho, tépalos internos rosados …………................ M. formosa Galeotti ex Scheidw. subsp. pseudocrucigera (R.T. Craig) D.R. Hunt
7a. Espinas centrales (4-)5-7(-8), flores con los tépalos internos amarillos …………………………………………………………….…... M. ariasii U. Guzmán & D. Aquino
7b. Espinas centrales 2-4, flores con los tépalos internos magenta ……………...……………. 8
8a. Plantas simples, por lo general profusamente ramificadas, tubérculos 0.6-1 cm de alto ………………………………………………………..........................……… M. geminispina Haw.
8b. Plantas simples, ocasionalmente ramificadas, tubérculos 0.5-0.6 cm de alto ……………………………………………………………………………. M. hahniana Werderm.

