












 (pdf)
(pdf)




 SciELO
SciELO  SciELO
SciELO 
 Permalink
Permalink
Introducción
El cambio climático global (CCG) es uno de los mayores desafíos que enfrenta la humanidad en el siglo XXI. No solo la biodiversidad y la salud humana se ven directamente afectadas por este fenómeno, también la producción agrícola, ya que el cambio climático impacta significativamente el desarrollo de las plantas y su rendimiento. Además, el CCG facilita el establecimiento y la propagación de plagas (insectos, patógenos y malezas) (Garrett et al., 2021), alterando así las interacciones plaga-hospedero. Se ha observado un incremento en la aparición de ciertas enfermedades (Gvozdenac, Dedic, Mikic, Ovuka y Miladinovic, 2022) o modificaciones en sus patrones de distribución espacial y temporal (Ghini, Hamada y Bettiol, 2008). Como resultado, aumentan las pérdidas de cultivos y las plagas y enfermedades se propagan rápidamente, con consecuencias devastadoras para el sector agrícola.
Una de las especies que se ha visto afectada por esta situación es el banano (Musa paradisiaca), el cual es considerado el cuarto alimento más importante del planeta (Suárez, Ortega y Jaimes, 2019). Es una fruta de consumo frecuente, con más del 33% de las personas consumiendo de tres a cinco bananos por semana (Flórez y Agudelo, 2019). En Ecuador, es considerado uno de los principales productos agrícolas y es el principal exportador a nivel mundial, representando el 29% del mercado internacional (Ajila, Aguilar, Romero y Campoverde, 2023). El clima tropical de Ecuador es un ambiente ideal para la producción constante de esta fruta durante todo el año (Armijos, Aguilar, Romero y Campoverde, 2022).
En el contexto del cambio climático global, el impacto de ciertas enfermedades puede exacerbar las afectaciones graves sobre este cultivo (Álvarez, Llano, Loke y González, 2007). La marchitez letal o enfermedad del moko causada por Ralstonia solanacearum (Smith, 1896) es una de las enfermedades bacterianas más agresiva y mortal de la familia Musaceae y, por lo tanto, representa un riesgo potencial significativo para la producción de banano en Ecuador. Este patógeno, transmitido por el suelo, habita en todos los continentes y muchas islas en los trópicos (Elphinstone, 2005) y puede afectar a más de 200 especies de plantas de 50 familias (Denny, 2007), como las solanáceas, muchas malas hierbas, cultivos, arbustos y árboles (Genin y Denny, 2012). Las enormes pérdidas económicas a nivel mundial y la falta de medidas de gestión efectivas contra esta bacteria, cuyo control se limitaba a medidas preventivas, uso excesivo de agroquímicos y erradicación de cultivos enfermos, terminaron por reconocerla como fitopatógeno A2:58 (alto riesgo) por parte de la Organización Europea y Mediterránea de Protección de las plantas para regular y limitar su propagación a nivel mundial.
La agricultura tropical del Ecuador es muy favorable a la presencia y propagación de esta plaga, e indirectamente, los cambios en las condiciones climáticas pueden exacerbar esta situación. En Ecuador, R. solanacearum ha causado pérdidas económicas en la producción de banano (Vivas-Cedeño, González, López, Tacuri y Palacios, 2022). Por estas razones, deben desarrollarse nuevas estrategias basadas en los principios del manejo integrado de plagas para mejorar la eficiencia del control (Norris, Caswell y Kogan, 2003). Se necesita información detallada sobre las características moleculares de las bacterias, las condiciones climáticas favorables para su desarrollo y el estudio de su dinámica poblacional para apoyar la toma de decisiones (Fonseca-Lacerda, Coelho, Gomes, Sentelhas y Postali, 2019). Por lo tanto, comprender la posible variación espacial del rango de distribución adecuado de la plaga en escenarios climáticos futuros puede proporcionar una base científica para controlar su propagación (Jiang et al., 2022).
Gracias al desarrollo continuo de métodos de simulación climática combinados con datos de pronóstico del tiempo, el uso de modelos de distribución potencial de especies puede predecir la distribución de plagas importantes en condiciones climáticas futuras (Jeschke y Strayer, 2008). Varios estudios han predicho rangos adecuados para diferentes plagas bajo escenarios climáticos futuros (Ramasamy, Das y Ramesh, 2021; Ferrer-Sánchez, Mafaldo, Plasencia y Urdánigo, 2022). Sin embargo, posibles cambios en la distribución de R. solanacearum no se han estudiado en Ecuador, aunque su impacto en la producción agrícola ha afectado significativamente la seguridad alimentaria. En este sentido, el propósito de este estudio es evaluar el impacto potencial del cambio climático en la distribución geográfica potencial de R. solanacearum y su impacto en las plantaciones de banano en Ecuador continental. Los hallazgos de este estudio son cruciales para entender la trayectoria futura de R. solanacearum en las plantaciones bananeras ecuatorianas en medio de condiciones climáticas cambiantes. Como han destacado Álvarez-Restrepo, Rodríguez y Marín (2008), R. solanacearum representa una amenaza significativa para la producción mundial de banano debido a su virulencia y persistencia. Mediante la predicción de su potencial propagación geográfica bajo escenarios de cambio climático, esta investigación proporciona información útil para las estrategias de gestión preventiva de la enfermedad. Además, como se subraya en la literatura reciente, estas medidas proactivas son esenciales para salvaguardar la estabilidad económica de las comunidades dependientes del banano en la región (Ramos-Veintimilla et al., 2024). En última instancia, este estudio pretende dotar a las partes interesadas de los conocimientos necesarios para mitigar el impacto de R. solanacearum, garantizando así el cultivo sostenible del banano en Ecuador.
Materiales y Métodos
Área de Estudio
Ecuador está ubicado en la región noroeste de América del Sur, limitando al norte con Colombia, al este y sur con Perú, y al oeste con el Océano Pacífico (01° 28’ N y 05° 01’ S; 75° 11’ y 81° 01’O) (Farfán, 2018). Su territorio abarca una superficie de aproximadamente 283 561 km², lo que lo convierte en uno de los países más pequeños de Sudamérica (Figura 1).
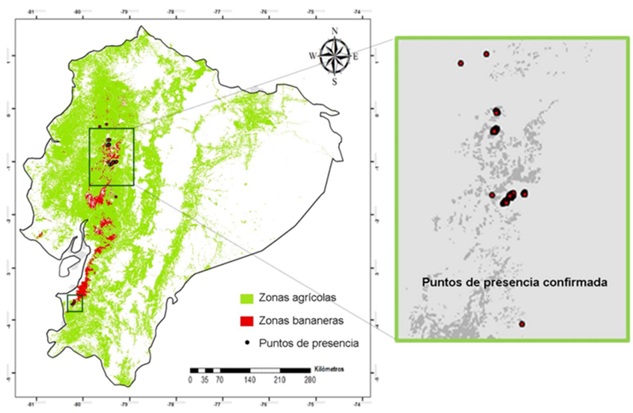
Compilación y Limpieza de Registros de Presencia
Se recopilaron registros de presencia georreferenciados de R. solanacearum de varias fuentes, principalmente, bases de datos nacionales e internacionales, muestreos de campo y fuentes bibliográficas. La principal base fue el Global Biodiversity Information Facility (GBIF, 2023), en la cual se ubicaron 672 registros. También se incluyeron registros tomados de la base Global Distribution of plant pathogenic Ralstonia strains (Lowe-Power et al., 2020) y EPPO Datasheet: Ralstonia solanacearum species complex (EPPO, 2024). Además, se solicitaron los datos de las bases del Instituto Nacional de Investigaciones Agropecuarias de Ecuador.
Los registros fueron filtrados y se excluyeron del análisis los que no tenían la información geográfica, presentaban errores como coordenadas incompletas, ubicaciones sin sentido o con metadatos contradictorios. Luego de la recopilación y limpieza de los datos de presencia se lograron 340 registros totales, de ellos 218 con alcance global, 122 en Sudamérica y de estos 62 en Ecuador continental. Los registros globales y a nivel del continente americano (excluye a Ecuador) se emplearon solamente para tener una referencia de la variabilidad de condiciones climáticas en las que puede aparecer el patógeno, mientras que en las modelaciones solo se emplearon los registros en el área de Ecuador continental.
Datos Ambientales
Para describir el nicho ecológico general de este patógeno se siguieron dos enfoques: uno estadístico multivariado, acorde al carácter multidimensional del nicho, y uno de máxima entropía para la proyección geográfica del nicho y la obtención de los mapas de adecuación de hábitat y de presencia potencial. El primer enfoque permite describir el nicho de la especie en Ecuador en el contexto del espacio ambiental general de la especie y del ambiente de la región, con énfasis en las características del clima. El segundo enfoque posibilita la proyección de la distribución geográfica potencial de la especie a los escenarios futuros de cambio climático. Para esta caracterización cuantitativa del nicho se emplearon dos tipos de variables ambientales: variables bioclimáticas y variables biogeofísicas obtenidas por sensores remotos.
Los datos bioclimáticos correspondientes a la actualidad se tomaron del WorldClim 2.0 (Hijmans, Cameron, Parra, Jones y Jarvis, 2005; Fick y Hijmans, 2017) a una resolución espacial de 30 segundos de arco (~1 km). Este modelo climático incluye 19 variables bioclimáticas que representan factores extremos o limitantes, ecológicamente importantes. Se siguieron los criterios de Escobar, Lira, Medina y Peterson (2014) y Ferrer-Sánchez et al. (2022) y se excluyeron las variables climáticas Bio8, Bio9, Bio18 y Bio19, al ser variables climáticas derivadas no relacionadas con la ecología de la especie.
Las variables biogeofísicas fueron obtenidas de información satelital. Se obtuvieron dos índices espectrales de vegetación como una medida de salud de los elementos vegetales y de la actividad fotosintética: el índice de vegetación de diferencia normalizada (NDVI) y el índice de vegetación mejorado (EVI), con indicadores de su variabilidad temporal (DS) en cada caso. Ambos se obtuvieron del producto compuesto del MODIS (MOD13Q1.006) promedio de 16 días, con 250 m de resolución espacial. También se obtuvo el Índice de área foliar (leaf area index). Como descriptores de propiedades superficiales del suelo se extrajo el Índice de humectación superficial (surface wetness) del producto SPL4SMGP-006 y la temperatura superficial del suelo (Land Surface Temperature) del producto satelital ECOSTRESS (ECO2LSTE-001). La altitud (m) se obtuvo del Modelo Digital de Elevación (Digital Elevation Model - DEM) del Shuttle Radar Topography Mission (SRTM v4.1) de la NASA (NASADEM_NC.001).
A toda esta información se accedió usando distintos servicios online. Se empleó, en primer lugar, la plataforma AppEEARS versión 3.28 (Application for Extracting and Exploring Analysis Ready Samples), (AppEEARS Team, 2022). Este es un sistema de cómputo en la nube que permite acceder a datos precisos, ya pre-procesados y listos para su análisis, del centro de datos LP DAAC (Land Processes Distributed Active Archive Center) del NASA Earth Observing System (AppEEARS Team, 2022). En este sitio se solicitaron los valores diarios de todos los puntos entre enero de 2020 y diciembre de 2022, que rindieron un total de 25 988 muestras. Los datos fueron filtrados para seleccionar solo los que cumplieran con los máximos indicadores de calidad en las bandas correspondientes (usando las bandas de control de calidad-QC) y se trabajó con los promedios generales por cada punto.
El Potencial de evapotranspiración (PET) y el índice de aridez (AI) se obtuvieron de los mapas generados por la Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO), a partir de los modelos de WordClim y ubicados en la base de datos CGIAR-CSI Global-Aridity and Global-PET (Zomer, Trabucco, Bossio y Verchot, 2008). Estas son variables derivadas del clima que permiten evaluar la pérdida potencial de agua del suelo por evaporación a partir de la temperatura y el gradiente de humedad relativa, y cuantifican el grado de suficiencia o insuficiencia de las precipitaciones para el sostenimiento de un ecosistema.
Finalmente, como medidas de heterogeneidad ambiental general a pequeña escala y en reflejo de las propiedades espaciales de las áreas se utilizaron varios índices derivados del EVI (Enhanced Vegetation Index): el índice de Shannon, la entropía, la homogeneidad espacial, el rango, máximo y la desviación estándar de los valores. Estos datos se obtuvieron para cada zona de presencia del GoogleEarth Engine (Tuanmu y Jetz, 2015).
La información ambiental (climática y biogeofísica) se extrajo de cada registro de presencia del patógeno y como marco de referencia, se extrajo la misma información en 10 000 puntos aleatorios distribuidos en toda el área de Ecuador continental. Estos puntos permitieron describir el espacio ambiental potencial del país, y dentro del mismo, ubicar el espacio ambiental de las zonas de cultivo del banano y las zonas específicas ya afectadas por moko. Todo el trabajo con los mapas, registros de presencia y puntos aleatorios se hizo con el uso del ArcGIS 10.1 (Esri, 2011).
Modelación de Nicho Ecológico
Para la modelación del nicho y la obtención de mapas de distribución potencial actual y futura se emplearon solo los registros de presencia dentro del área de Ecuador continental y las variables bioclimáticas. Para ello se eliminaron los registros que aparecían repetidos dentro de un mismo píxel o a una distancia menor de 1 km. La limpieza de los registros se realizó con los paquetes spThin (Aiello-Lammens, Boria, Radosavljevic, Vilela y Anderson, 2015) y Niche Toolbox (Osorio-Olvera et al., 2020) de R.
El modelo de nicho en la actualidad fue proyectado a dos escenarios futuros de cambio climático, en los años 2050 y 2070. Para ello se emplearon dos escenarios climáticos futuros correspondientes al modelo de circulación general GISS-E2-R (NASA Goddard Institute for Space Studies, USA). Se seleccionaron dos escenarios del cambio climático del Panel Intergubernamental sobre Cambio Climático (IPCC) (van-Vuuren et al., 2011) para dos periodos, uno con su centro en 2050 (2041-2060) y otro lejano centrado en 2070 (2061-2080). Los dos escenarios representaron distintas trayectorias de concentraciones de gases de efecto invernadero en la atmósfera, uno moderado (2-45) donde se asume que las emisiones ya alcanzaron su punto máximo y se predice un aumento de temperatura promedio de 1.4 °C (0.9-2.0 °C), y otro escenario extremo (5-85) donde las emisiones seguirán aumentando durante el siglo veintiuno que predice un aumento de temperatura promedio de 2.0 °C (1.4-2.6 °C) (Ward, Morh, Myres y Nel, 2012). Elegir estas proyecciones climáticas permite alinear el trabajo con el marco de las Naciones Unidas de los desafíos mundiales en la agricultura (Lipper et al., 2014). Se proyectó el modelo de nicho actual a los dos escenarios seleccionados en cada año para predecir la distribución potencial de la especie en ambos casos y evaluar sus diferencias con la obtenida en la actualidad e identificar las zonas más vulnerables.
El proceso de modelado se realizó con el algoritmo de máxima entropía en el programa MaxEnt versión 3.4.0 (Phillips, Anderson, Dudík, Schapire y Blair, 2017). Este es un método correlativo que sólo requiere de información sobre la presencia de las especies y una serie de variables ambientales para generar un modelo de las condiciones que favorecen la presencia de una especie en particular (Phillips et al., 2017). El modelo estadístico obtenido se proyectó al espacio geográfico para generar un mapa que representa la distribución de las condiciones favorables para la especie, o su distribución potencial (Phillips et al., 2017).
Se seleccionó como área accesible la región continental de Ecuador, que fue el área de interés de este estudio y se mantuvieron los parámetros de multiplicador de regularización y tipos de clases por defecto en el programa, que son conservativos y permiten al algoritmo llegar cerca de la convergencia (Phillips et al., 2017). Para la calibración del modelo, se realizó una validación cruzada mediante la asignación aleatoria (sin reemplazo) de datos de ubicación. El 70% de los datos de presencia se utilizaron para calibrar el modelo y el otro 30% para validarlo. Se consideraron como parámetros una tasa de omisión de 5%, un porcentaje de puntos aleatorios de 50% y 500 iteraciones de bootstrap (Peterson, Papeş y Soberón, 2008). Para evaluar el desempeño del modelo se usó el área bajo la curva ROC parcial (AUC) (Phillips, Anderson y Schapire, 2006; Osorio et al., 2020). La contribución de las variables a los modelos se evaluó mediante Jackknife.
Evaluación de las Zonas de Riesgo
Para evaluar las zonas de riesgo de expansión de la enfermedad, se utilizó la diferencia normalizada entre la idoneidad actual del patógeno y su idoneidad proyectada bajo escenarios de cambio climático como indicador de riesgo. Este enfoque permitió evaluar la probabilidad incrementada de presencia del patógeno en cada área afectada por este efecto. El mapa de idoneidad para la línea base y los climas futuros se convirtió en mapas binarios de presencia-ausencia (0 y 1) usando como umbral de corte el de máxima sensibilidad (verdadero positivo) + especificidad (verdadero negativo) (Liu, Berry, Dawson y Pearson, 2005; Liu, White y Newell, 2013).
Se usó el mapa de uso de suelo de Ecuador para extraer las zonas de agricultura dedicadas al cultivo del plátano, que representarán la zona potencial de distribución. A partir de los valores del indicador de riesgo se clasificaron en tres categorías: Alto, Medio y Bajo Riesgo, considerando además la distancia mínima a zonas de presencia confirmada de la enfermedad.
Para conocer las posibles expansiones, contracciones o estabilidad de la distribución potencial en el futuro del patógeno (Rangos de ampliación o de pérdida) se realizaron comparaciones del mapa binario de cada escenario con la herramienta Distribution Changes Between Binary SDMs que aparece en la caja de herramientas SDMtoolbox 2.4 en ArcGis (Esri, 2011). A partir de este análisis, se obtuvo la extensión del área (km2) para cada escenario y se visualizaron los rangos de cambios posibles en cada escenario futuro.
Análisis de Datos
Los datos ambientales fueron descritos a partir de los valores medios, su desviación estándar y el intervalo de confianza por el método de los percentiles. Para evaluar las posibles redundancias en la información de las variables ambientales se redujo la multidimensionalidad con un Análisis de Componentes Principales, que permitió analizar el nicho ecológico desde el enfoque estadístico, acorde al carácter multidimensional del mismo. El área o volumen de las nubes de puntos en los mapas de ordenamiento de los componentes de mayor aporte de varianza se emplearon como indicadores del hipervolumen ambiental o del nicho del patógeno. Un análisis de superposición permitió comparar el hipervolumen ambiental del patógeno con el disponible en las zonas bananeras de Ecuador, evaluando su grado de superposición y centralidad. Las representaciones gráficas de los resultados se diseñaron en el programa R (R Core Team, 2020) y en las plataformas Extended Boxplot (Denis y Ramírez, 2020); Extended Scatterplots (Denis y Ramírez, 2021) y Rawgraphs 2.0.
Resultados y Discusión
Nicho ambiental de Ralstonia solanacearum en Ecuador Continental
La alta variabilidad y heterogeneidad de variables ambientales empleadas dificultó la concentración de varianza explicada en pocos ejes, por lo que se llegó a nueve componentes efectivos que explican más del 90% de la varianza del sistema (Figura 2). La proyección de los dos primeros componentes formó una nube de puntos que refleja casi la mitad de la variabilidad de las 29 dimensiones utilizadas para describir el nicho ambiental de R. solanacearum, el cual muestra una extensión general semejante en sus registros globales (excluye América y Ecuador) y aquellos restringidos a América (Figura 3).
Figura 2: A) Espacio formado por la reducción de 29 dimensiones del nicho ambiental por medio de un Análisis de Componentes Principales (ACP), del patógeno Ralstonia solanacearum donde se compara la extensión del nicho en Ecuador continental, en relación con el nicho global (Excluye a América) y en América. B) Componentes efectivos (90% de varianza explicada) del ACP.
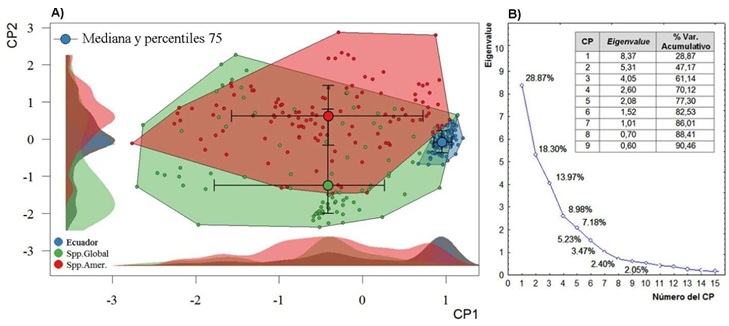
Figure 2: A) Space formed by the reduction of 29 dimensions of the environmental niche through a Principal Component Analysis (PCA), of the pathogen Ralstonia solanacearum where the extension of the niche in continental Ecuador is compared, in relation to the global niche (America omitted) and in America. B) Effective components (90% of explained variance) of the PCA.
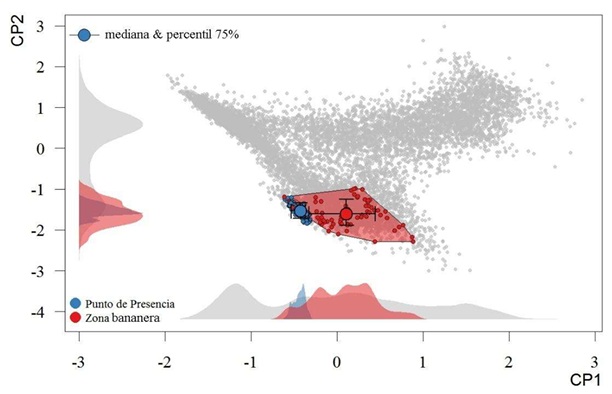
En el contexto de esta variación global, los registros de presencia en Ecuador continental se encuentran más restringidos y concentrados hacia uno de los extremos del nicho general. Esta baja centralidad sugiere que las condiciones en Ecuador no suelen ser las típicas para el patógeno, aunque aún se incluyen dentro del rango de variabilidad que puede soportar. La proyección de los espacios ambientales utilizados por la especie evidencia que, el patógeno también tiende a ocupar combinaciones de variables ambientales que tienden a diferenciarse de los más típicos dentro del país y hacia un extremo de las condiciones generales de las zonas de cultivos de banano (Figura 3). Esta particular disposición sugiere que las condiciones actualmente, si bien permiten la presencia del patógeno, no son las más apropiadas para tener una invasión a gran escala, al menos predeciblemente, y que son relativamente reducidas las zonas con las condiciones que suele tener en los lugares donde habita.
En el primer eje del ACP, las dos variables de mayor influencia fueron relacionadas con las temperaturas mínimas [Temperatura mínima del mes más frío (BIO6) y Temperatura media del trimestre más frío (BIO11)]. Les siguieron en importancia las máximas precipitaciones [Precipitación del trimestre más húmedo (BIO16) y Precipitación del mes más húmedo (BIO13)]. Esto ratifica que, en Ecuador, los principales factores climáticos que determinan o regulan la distribución de las zonas de adecuación potencial para este patógeno son las temperaturas mínimas y máximas precipitaciones. Estas dos variables también son determinantes para el cultivo del banano, que prefiere un clima cálido húmedo con temperatura media óptima de 27 °C (rango de 26 a 27 °C, según la variedad), y un alejamiento de este rango óptimo puede enlentecer el crecimiento y desarrollo del cultivo o deteriorar las plantas (Gómez-Calle, 20171). El cultivo también necesita un régimen de lluvias bien distribuidas y amplias, idealmente con precipitaciones de 1800 mm anuales. La temperatura y humedad del suelo son factores que también influyen sobre la severidad de la enfermedad del moko, así como sobre los hospedantes susceptibles (Florez y Agudelo, 2019).
Los patrones de las variables biogeofísicas no fueron tan consistentes, probablemente debido a la homogeneidad de propiedades físicas generales de los cultivos de banano. A nivel espacial, la entropía, la heterogeneidad y los valores máximos de EVI tendieron a ser iguales entre las zonas de presencia y las medias generales de los cultivos. Estas variables son buenas caracterizadoras de las propiedades de la vegetación, pero tienen más utilidad en la discriminación o comparación de formaciones vegetales tipológicamente más diferentes (Hao, Zhao, Fan, Zhang y Zhang, 2023). Las variables de variabilidad temporal (rango y desviación estándar) fueron diferentes, pero de manera estadísticamente marginal y poco tamaño de efecto.
El mapa de distribución de valores de adecuación del hábitat muestra condiciones similares a las requeridas por la especie de patógeno en el centro norte del país (Figura 4). La mayor zona de probabilidad de presencia se encuentra al norte de la provincia Los Ríos y al extremo sur de Esmeraldas, incluyendo parte de Cotopaxi, Manabí y Santo Domingo de los Tsáchilas. El modelo que generó este mapa tuvo un buen rendimiento, con valores altos de AUC de 0.999, tanto para los datos de entrenamiento como los de prueba.
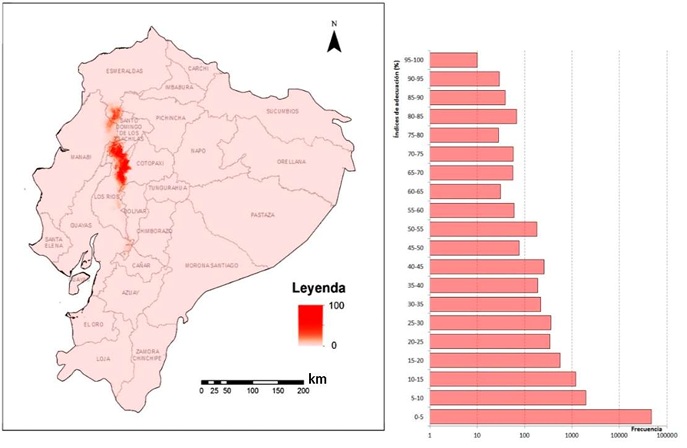
Las variables que más aportaron de forma individual al modelo fueron las precipitaciones del mes y del trimestre más húmedos y la estacionalidad de las precipitaciones (BIO13, BIO16 y BIO15). Le siguieron en aporte la estacionalidad de la temperatura (BIO4) y la precipitación anual (BIO12). El intervalo anual de temperatura fue la menos relevante para el modelo (BIO7). En general, el área de distribución potencial del patógeno muestra una superficie relativamente conservadora en comparación con el país y las zonas de cultivo del banano según el clima en la actualidad. Esto es un elemento esperanzador, ya que permite suponer que no es tan propicia la situación actual para una invasión a gran escala de la enfermedad, y que hay mayores posibilidades para un control exitoso. Los puntos con valores superiores al 90% de adecuación del hábitat son relativamente escasos, pero una proporción muy superior está en el rango de adecuación intermedia. Son precisamente esas áreas las que presuponen un riesgo futuro, ya que en dependencia de la dirección de cambio de las condiciones climáticas pueden derivar en zonas de probabilidad de presencia más fuerte.
Ampliaciones y Contracciones en la Distribución Geográfica Potencial de Ralstonia solanacearum ante Escenarios de Cambio Climático
Las diferencias entre los valores bioclimáticos actuales y futuros determinan diferencias en el espacio geográfico físico de la distribución potencial del patógeno. Las proyecciones de cambios climáticos en las zonas de cultivo del banano en Ecuador predicen, para el año 2050, una alteración general que se reflejan en un desplazamiento de las nubes de puntos multidimensionales en el espacio de componentes principales (Figura 5). Este desplazamiento es mucho más marcado en el caso del modelo climático 5-85 (Figura 5b) y se manifiesta de forma más fuerte en el eje del primer componente, que recoge cerca del 40% de la varianza total (los tres primeros componentes ya alcanzan más del 90%). En este primer eje la temperatura y sus derivadas son las variables que tienen un mayor peso. Los puntos de presencia conocida del patógeno en la actualidad tienden a estar ubicados hacia la parte inferior de la nube de puntos, y se asume que con el cambio climático mantiene aproximadamente la misma ubicación y dispersión, de acuerdo a la teoría del conservadurismo del nicho ecológico. La suma de estas variaciones climáticas es la causante del patrón de cambio observado en la Figura 5, y en los cambios modelados en las áreas de distribución potencial del patógeno, dado mayormente a que no se esperan cambios en las zonas de uso de suelo destinados a este cultivo.
Figura 5: Cambios en el espacio climático multidimensional en los escenarios futuros de cambio climático de las trayectorias socioeconómicas compartidas (SSPs) 2-45 y 5-85 para el año 2050 en las zonas de cultivo de banano en Ecuador continental, con relación a las condiciones actuales y específicamente a las que aparecen en los puntos de presencia confirmada del patógeno Ralstonia solanacearum.
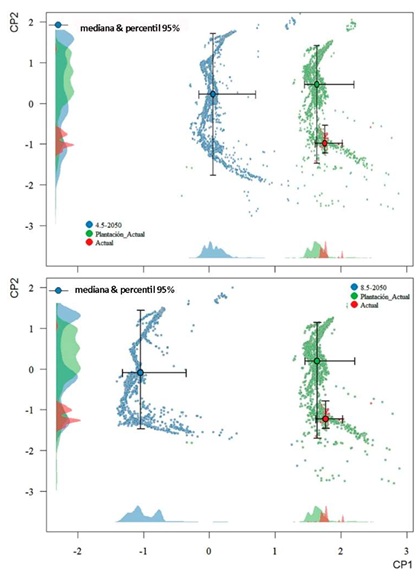
Figure 5: Changes in the multidimensional climate space in the future climate change scenarios of the Shared Socioeconomic Pathways (SSPs) 2-45 and 5-85 for the year 2050 in the banana plantations areas of continental Ecuador, in relation to current conditions and specifically those that appear in points of confirmed presence of the pathogen Ralstonia solanacearum.
Desglosando este patrón general multivariado en sus componentes se puede observar que existen diferencias marcadas en las variables climáticas actuales con las previstas en ambos escenarios, en ambos casos muy superior a lo planteado como nivel deseado por los organismos internacionales (Cuadro 1). La temperatura media anual, según el escenario 2-45 aumentaría en las áreas bananeras en 3.3 °C como promedio, y en el escenario 5-85 en 5.2 °C. Las precipitaciones medias anuales, sin embargo, se espera que se reduzcan en ambos escenarios, aunque el comportamiento relativo es diferente: el proyectado por el 2-45 tendría menor cambio en la temperatura, pero mayor reducción en las precipitaciones (en un 26.5%) que el escenario 5-85, en el cual se reducirían en un (24.8%).
Cuadro 1: Comparación de los valores climáticos medios en los puntos de presencias registradas actualmente del patógeno Ralstonia solanacearum en Ecuador continental y valores medios en los mismos lugares, pero en escenarios climáticos futuros (2050 y 2070) según dos trayectorias representativas de concentraciones de gases.
Table 1: Comparison of the average climatic values in the points of currently recorded presence of the pathogen Ralstonia solanacearum in continental Ecuador and average values in the same places, but in future climatic scenarios (2050 and 2070) according to two representative trajectories of gas concentrations.
| Variable | Escenario (N=2366) | ||||
| Actual | 2-45-2050 | 2-45-2080 | 5-85-2080 | 5-85-2050 | |
| Índice de Adecuación | 85.16±14.55 | 35.39±44.74 | 30.35±39.89 | 51,1±44.98 | 7.7±19.13 |
| (82.89-87.43) | (33.57-37.2) | (28.74-31.96) | (49.29-52.92) | (6.9-8.51) | |
| Bio_1 | 24.52±2.89 | 27.80±4.95 | 27.80±4.95 | 29.66±5.21 | 29.7±5.13 |
| (24.48-24.57) | (27.78-27.82) | (27.78-27.82) | (29.64-29.68) | (29.68-29.72) | |
| Bio_2 | 86.01±3 | 86.26±5.44 | 86.26±5.44 | 88.29±5.21 | 88.39±5.31 |
| (85.54-86.48) | (86.04-86.48) | (86.04-86.48) | (88.1-88.5) | (88.2-88.6) | |
| Bio_3 | 76.4±0.97 | 74.72±4.08 | 74.72±4.08 | 76±3.51 | 75.92±3.62 |
| (76.25-76.55) | (74.56-74.89) | (74.56-74.89) | (75.86-76.14) | (75.77-76.07) | |
| Bio_4 | 801.77±57.52 | 915.77±164.16 | 915.77±164.16 | 883.6±151.43 | 892.37±152.76 |
| (792.79-810.75) | (909.16-922.39) | (909.16-922.39) | (877.5-889.71) | (885.99-898.75) | |
| Bio_5 | 3032±4.03 | 33.72±7.21 | 33.72±7.21 | 35.53±6.19 | 35.58±6.08 |
| (30.26-30.38) | (33.69-33.74) | (33.69-33.74) | (35.51-35.56) | (35.55-35.60) | |
| Bio_6 | 19.13±2.51 | 22.24±4.41 | 22.24±4.41 | 23.99±4.27 | 24.01±4.29 |
| (19,86-19,165) | (22.22-22.25) | (22.22-22.25) | (23.98-24.01) | (23.99-24.03) | |
| Bio_7 | 11.19±3.59 | 11.48±5.22 | 11.48±5.22 | 11.54±4.42 | 11.57±4.31 |
| (11.13-11.24) | (11.46-11.5) | (11.46-11.5) | (11.53-11.56) | (11.55-11.59) | |
| Bio_10 | 25.53±2.87 | 28.91±5.49 | 28.91±5.49 | 30.76±5.05 | 30.80±4.95 |
| (25.48-25.57) | (28.88-28.93) | (28.88-28.93) | (30.74-30.78) | (30.78-30.82) | |
| Bio_11 | 23.57±2,73 | 26.66±4.18 | 26.66±4.18 | 28.55±4.61 | 28.56±4.68 |
| (23.53-23.61) | (26.64-26.62) | (26.64-26.68) | (28.53-28.56) | (28.54-28.58) | |
| Bio_12 | 2447.1±329.33 | 1798.08±907.77 | 1798.08±907.77 | 1924.76±928.19 | 1838.72±896.24 |
| (2395.7-2498.5) | (1761.5-1834.7) | (1761.5-1834.7) | (1887.3-1962.2) | (1801.3-1876.2) | |
| Bio_13 | 491.69±60.73 | 347.62±155.64 | 347.62±155.64 | 366.96±182.05 | 349.5±174.91 |
| (482.2-501.17) | (341.35-353.9) | (341.35-353.9) | (359.62-374.3) | (342.2-356.8) | |
| Bio_14 | 22.76±7.19 | 11.97±11.49 | 11.97±11.49 | 5.82±4.75 | 5.83±4.79 |
| (21.64-23.88) | (11.51-12.44) | (11.51-12.44) | (5.63-6.01) | (5.63-6.03) | |
| Bio_15 | 91.79±4.06 | 83.21±8.25 | 83.21±8.25 | 75.86±3.94 | 75.97±4.04 |
| (91.15-92.42) | (82.88-83.54) | (82.88-83.54) | (75.7-76.02) | (75.8-76.13) | |
| Bio_16 | 1403.28±174.85 | 964.53±446.38 | 964.53±446.38 | 976.63±464.71 | 934.37±449.96 |
| (1375.9-1430.6) | (946.54-982.53) | (946.54-982.53) | (957.89-995.36) | (915.58-953.16) | |
| Bio_17 | 83.54±25.44 | 64.55±47.38 | 64.55±47.38 | 69.67±38.8 | 66.93±38.48 |
| (79.57-87.51) | (62.64-66.46) | (62.64-66.46) | (68.11-71.24) | (65.32-68.53) | |
Del resto de las variables, aquellas con cambios más marcados en ambos escenarios serían las precipitaciones del mes más seco, que disminuirían en 47% y 74% respectivamente. Las temperaturas mínimas del mes más frío, que en los modelos climáticos actuales para esta zona promedian 19.1 °C, llegarían a 24 °C (un 25.5% de cambio). El rango diurno medio de la temperatura se proyecta que se incremente en estas zonas en 2.4 °C en el escenario más drástico.
Los modelos de distribución potencial para R. solanacearum en Ecuador continental muestran varios escenarios de expansión posible del patógeno (Figura 6). En el escenario de cambios más discretos (Figura 6C), para el año 2050, podría esperarse una expansión muy intensa hacia los cultivos del norte de Ecuador, cubriendo una parte significativa de Esmeraldas y Manabí. Un núcleo de zonas muy adecuadas aparecería en medio de Santa Elena. Además, podría presentarse un corredor de invasión hacia el sur, a lo largo de los límites entre Los Ríos y Bolívar, atravesando el sur de Guayas hasta extenderse a lo largo de la costa sur de esta provincia y tocar el extremo superior de El Oro. Sin embargo, en este mismo escenario de cambios, para el 2080 la distribución de las áreas de adecuación se re-estructuraría nuevamente (Figura 6D). La extensión al sur de las áreas de adecuación se recogería al norte, quedando solo afectada de manera significativa la provincia Los Ríos, porciones significativas de Santo Domingo de los Tsáchilas, Pichincha, Esmeraldas y Manabí.
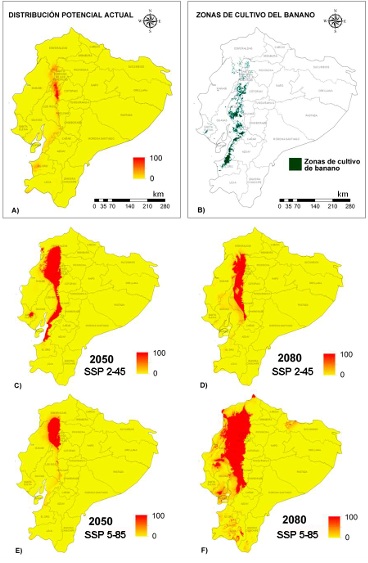
En el escenario 5-85 el comportamiento es diferente, debido a que para el año 2050 debería esperarse una expansión del área potencial de distribución, pero concentrada en los alrededores de la zona de presencia actual real y la modelada para el clima actual (Figura 6). En este escenario, se reduce la extensión de Los Ríos afectada y se expande la distribución potencial a Esmeraldas y Manabí de forma significativa (Figura 6E). Sin embargo, bajo este mismo escenario, la predicción de la extensión de condiciones necesarias para el patógeno sufre de una expansión explosiva, cubriéndose casi totalmente la provincia Los Ríos y Santo Domingo de los Tsáchilas, y alrededor de un tercio de la extensión de las provincias Esmeraldas y Manabí.
Según las áreas clasificadas según su cambio con relación a la distribución actual: si fue una zona de ampliación del rango, disminución o un área que se mantuvo, la mayor parte de las áreas actuales de distribución se mantienen casi sin cambios, con mayores extensiones de nuevas áreas potenciales de presencia del patógeno y muy pocas zonas donde pudiera desaparecer por inadecuadas condiciones climáticas (Figura 7). Al calcular la extensión de estas áreas se observa que, según las modelaciones realizadas, en Ecuador continental en la actualidad existe una extensión superior a los 5800 km2 con condiciones climáticas consistentes con las de sus áreas de presencia conocidas (Figura 8A).
Figura 7: Mapas binarios obtenidos luego de la aplicación del umbral de corte al modelo de distribución potencial por máxima entropía para el patógeno Ralstonia solanacearum en Ecuador continental y sus proyecciones según los cambios climáticos previstos en los escenarios 2-45 y 5-85 para los años 2050 y 2080. Se muestra, además, la distribución en los tipos de cambios (reducción, mantención o ampliación) en la distribución potencial en relación a la actual.
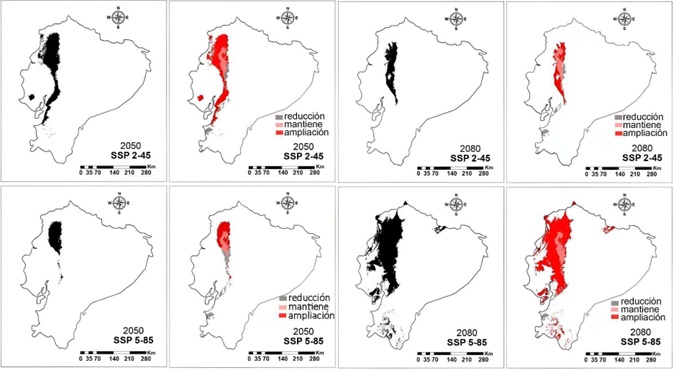
Figure 7: Binary maps obtained after applying the cut-off threshold to the potential distribution model by maximum entropy for the pathogen Ralstonia solanacearum in continental Ecuador and its projections according to the climate changes predicted in the scenarios 2-45 and 5-85 for the years 2050 and 2080. The distribution in the types of changes (reduction, maintenance or expansion) in the potential distribution in relation to the current one is also shown.
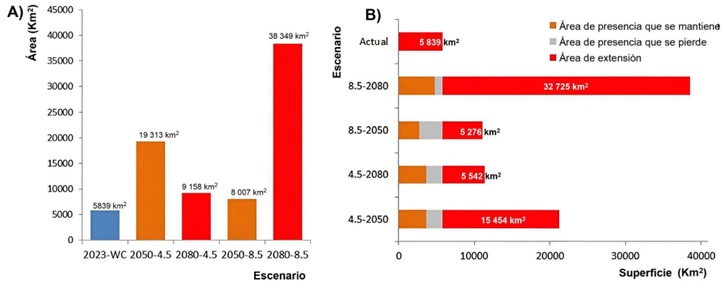
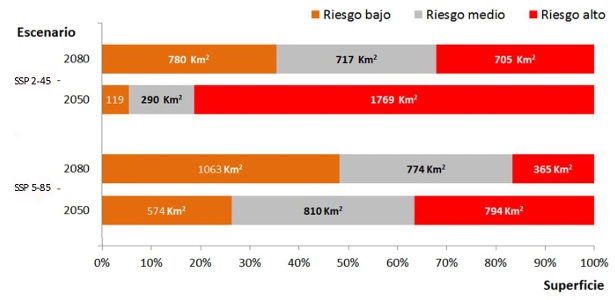
En el modelo proyectado a las predicciones para el 2050, asumiendo el escenario de cambio climático más conservador de los dos analizados, el área se incrementa en más de tres veces, para luego, en la proyección al 2080 disminuir a poco más de 9000 km2. En el escenario de cambio climático 5-85 los cambios se invierten: hacia el 2050 podría esperarse un incremento en un 37% del área adecuada y para el 2080 el área de idoneidad climática crece más de cinco veces (aumenta un 556%).
Casi toda la extensión de presencia actual se mantiene en las proyecciones futuras de cambios climáticos, siendo la tendencia general, en todas ellas, al incremento en las áreas (Figura 8B). En dependencia de la adecuación de los modelos del IPCC a la realidad, se espera que para el 2080 exista una extensión del área potencial del patógeno entre 5500 km2 (escenario más conservador) y 32 725 km2 (escenario más drástico).
Plantaciones Bananeras más Susceptibles a las Condiciones Favorables para el Patógeno en Ecuador Continental
Las áreas de mayor riesgo son los campos de cultivo que se ubican hacia el sur de Ecuador, a pesar de que el área de presencia potencial se extienda más hacia el norte, lo cual es debido a que los índices de idoneidad son muy bajos en esos escenarios (Figura 9). La mayor cantidad de zonas de cultivo de bananos clasificadas como de alto riesgo aparecen para el año 2050 en el escenario 2-45, en el cual existirían unos 2000 km2 de zonas de plantaciones en esta categoría debido a la cercanía entre sus condiciones bioclimáticas generales y las del área de presencia del patógeno (Figura 10). Según este modelo, en el año 2080 existirían menos zonas de riesgo debido a que la expansión en área de las zonas climáticas se debería a cambios en los valores de adecuación en una magnitud relativamente pequeña (Figura 10).
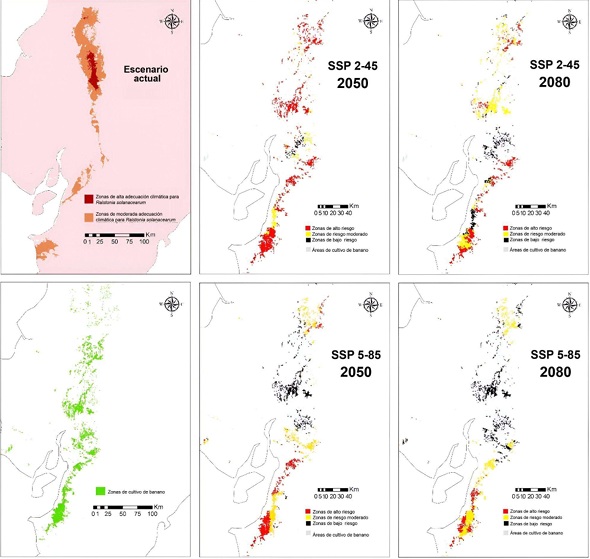
A nivel nacional se han identificado plantas enfermas de moko en Manabí, Guayas, Santo Domingo, Quevedo. También se ha comercializado y trasladado cormos (yemas basales secundarias de la planta de banano) desde la zona afectada a todo el país, sin las medidas de bioseguridad. Larrea (20202) refiere que uno de los mayores problemas en el campo es el mal manejo al detectar una zona con la enfermedad.
La provincia Los Ríos, que es la potencialmente más vulnerable y propensa al desarrollo de R. solanacearum, según INEC (2022) en 2021 produjo el 38.5% de la producción bananera de Ecuador (6 684 916 toneladas). El control de la enfermedad en esta provincia ha sido un problema debido a la falta de tecnologías para la identificación y el manejo; el cual se ha llevado a cabo de una forma tradicional y sin tener protocolos adecuados (Vivas-Cedeño et al., 2022). Aunque no existen protocolos definitivos que permitan la eliminación de la bacteria y de la enfermedad que provoca en un 100%, es necesario desarrollar medidas de manejo integral que pudieran reducir a un mínimo la diseminación de la bacteria en los campos de manera inmediata.
La presencia de malezas o plantas coexistentes con el banano, que actúan como hospederas o reservorios, también influye en el desarrollo epidemiológico de la enfermedad y dificultan su erradicación. Además, hacen más compleja la ecología del patógeno y pueden influir en su capacidad de adaptación a los cambios climáticos. Una forma de aumentar la precisión de los modelos predictivos del patógeno R. solanacearum podría ser incluir en este tipo de estudios los modelos de distribución de las otras especies de plantas que también son hospedantes y que coexisten en los cultivos de banano. Este patógeno posee un amplio rango de hospederos, de los cuales se habían descrito 234 especies para 2010 (Hernández, 2010) y ya en 2020 se habían reportado al menos 160 nuevas especies hospedantes (Lowe-Power et al., 2020). La presencia de malezas o plantas coexistentes con el banano o plátano, que actúan de hospederas o reservorios, influye en el desarrollo epidemiológico de la enfermedad y dificultan su erradicación. Además, hacen más compleja la ecología del patógeno y pueden influir en su capacidad de adaptación a los cambios climáticos.
Conclusiones
El nicho ecológico de R. solanacearum en Ecuador continental muestra una baja centralidad en comparación con el de la especie en el mundo, lo que indica que las condiciones climáticas actuales en Ecuador no suelen ser las más típicas para el patógeno, aunque aún están dentro del rango de variabilidad que puede soportar. Esta distribución casi periférica del nicho se mantiene cuando se ubica dentro del espacio climático general del país, y del espacio climático específico de las áreas de cultivo del banano. La distribución geográfica potencial de R. solanacearum, según la proyección geográfica binarizada del modelo de nicho ecológico asciende a 5839 km2, de la cual la mayor parte se superpone con las zonas de cultivo del banano actualmente. Si se cumpliera el escenario de Cambio Climático predicho por el escenario 2-45, esta extensión aumentaría a 15 454 km2 en el año 2050 y luego disminuiría a 5545 km2 en el 2080. Si se cumpliera el escenario de cambios más drástico (5-85) el aumento para el 2050 sería de 5276 km2, pero para el 2080 llegaría a 32 725 km2. Estos escenarios tan negativos son solo los máximos posibles, debido a que, al final, la magnitud de extensión de presencia del patógeno estará dada por el cumplimiento o no de un estricto protocolo de manejo y control de la enfermedad.
Las plantaciones con mayor probabilidad de riesgo de expansión de la enfermedad están hacia el centro-norte de Ecuador, en las provincias Los Ríos, Santo Domingo de los Tsáchilas, Manabí y Esmeraldas tanto por estar dentro de las zonas de distribución potencial como por la cercanía a los puntos de ocurrencia reales reportados del patógeno. Sin embargo, por similitud a las condiciones bioclimáticas de los puntos de presencia de la especie, zonas bananeras con alto riesgo se encuentran también a lo largo de Los Ríos hasta las provincias Guayas y El Oro.

