












 (pdf)
(pdf)




 SciELO
SciELO  SciELO
SciELO 
 Permalink
PermalinkINTRODUCTION
Electroma vexillum (Reeve, 1857) is a bivalve native to the Indo-Pacific, which is distributed in the Red Sea, the Persian Gulf, and southern India (Dekker and Orlin 2000, Zenetos 2010, Asha et al. 2016, Oliver et al. 2023, GBIF 2024). It differs from other bivalve species due to its distinctive morphological characteristics, such as small (<10 mm length), fragile and translucent valves, and the absence of hinge teeth (Çevik et al. 2008, Tëmkin 2010). Furthermore, E. vexillum inhabits shallow waters associated with macroalgae, seagrass beds, and artificial structures (Asha et al. 2016). This species has been reported as exotic in several locations in the Mediterranean Sea, such as Turkey (Çevik et al. 2008, Albayrak 2010, Çinar et al. 2021), and after almost 20 years of these records, its presence has been confirmed with a recent report in Israel (Albano et al. 2024). The presence of an introduced species identified as Electroma sp. has been reported in the western Atlantic, specifically in 1983 and 1995 on the coasts of Colombia and Venezuela (Borrero and Díaz 1998). However, this region lacks updated data, so the current status of this record is unknown (Álvarez-León et al. 2007).
In the Gulf of Mexico, which is the region closest to the southern Caribbean, the presence of E. vexillum as an introduced species has not been documented (Okolodkov et al. 2007, Mendoza-Alfaro et al. 2014, León-González et al. 2021, CONABIO 2024). Furthermore, E. vexillum does not appear in the taxonomic lists of marine and coastal ecosystems (Pérez-Rodríguez 1997, Hicks et al. 2001, García-Cubas and Reguero 2007, Tunnell et al. 2007, Turgeon et al. 2009, Correa-Sandoval and Rodríguez-Castro 2013, Suárez-Mozo et al. 2024), nor has it been reported in recent studies on the benthic macrofauna of the Veracruz coast (Vassallo et al. 2014, De la Cruz-Francisco et al. 2017, Rodríguez-Muñoz et al. 2023).
Northern Veracruz is characterized by a predominantly sandy coastline, although artificial protection structures, such as breakwaters and groins, are also present, as are rocky sandstone promontories to a lesser extent (López-Portillo et al. 2023, Rodríguez-Muñoz et al. 2023). These solid substrates provide habitats for macroalgae and marine macrofauna to establish; these have been previously documented in the region (De la Cruz-Francisco et al. 2017; Rodríguez-Muñoz et al. 2023; Mateo-Cid et al. 2024; De la Cruz-Francisco 2025a, b). However, studies have indicated that these artificial structures can also facilitate the establishment of introduced or exotic species (Vaselli et al. 2008, Mineur et al. 2012), a phenomenon that has been scarcely explored in this area.
Therefore, the aims of this study were to document the first records of the Indo-Pacific species E. vexillum in the Gulf of Mexico, describe the external morphological characteristics of specimens collected from 2 coastal areas in northern Veracruz, and contribute to the understanding of the distribution and habitat of this species in the region. The information provided by this study will serve as a basis for future studies evaluating the ecology of E. vexillum, its impact on local ecosystems, and its interaction with native species.
MATERIALS AND METHODS
Study sites: coastline of northern Veracruz
The coast of Tuxpan, located in the municipality of Tuxpan, is primarily sandy with a gentle slope. The Tuxpan River flows into the central part of the coastline, flanked by two ~1-km long breakwater structures, as well as 16 straight groins, each 60 m long, located perpendicular to the shoreline. Communities of benthic algae and marine invertebrates grow on the rocks of the Tuxpan coastline. This coastline is particularly active in national and international maritime traffic, due to the presence of an industrial and commercial seaport, with cargo moving overseas to the United States, Cuba, Russia, Canada, China, Latvia, and Ukraine (López-Portillo et al. 2023) (Fig. 1b; site A).
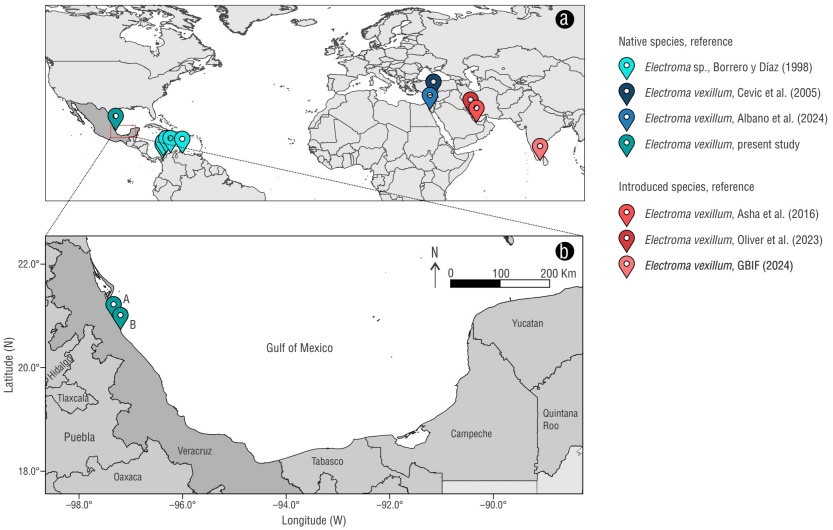
Figure 1 Distribution of Electroma vexillum as a native and introduced species (a). Coasts of Tuxpan (A) and Cazones (B) in the Gulf of Mexico (b).
The coast of Cazones, located in the municipality of Cazones de Herrera and bordering the Tuxpan municipality to the north, is sandy and has rocky sandstone promontories and a ~950-m-long and 40-m-wide abrasive platform, known as Playa El Pulpo, which is is inhabited by algae, grasses, and marine invertebrates (De la Cruz-Francisco et al. 2017, Rodríguez-Muñoz et al. 2023) (Fig. 1b; site B).
These coastlines experience seasonal changes throughout the year. During autumn (September-December) and winter (December-March), the water is cold (22-23 °C) and surface currents flow from north to south; in spring (March-June) and summer (June-September), the water is warmer (27-28 °C) and currents flow from south to north (Zavala-Hidalgo et al. 2003, Mendelssohn et al. 2017).
Sampling
The indirect method was used for organism sighting and collection. This method consisted in searching for small bivalves attached to the benthic macroalgae on the artificial structures along the coastline of Tuxpan and the rocky coastline of Cazones. Samplings were conducted during the lowest tides, walking along artificial and rocky substrates. The first collections occurred in November 2018 on the rocky coastline of Cazones, during samplings aimed at studying the benthic community (Rodríguez-Muñoz et al. 2023). Collected organisms were preserved in 70% ethyl alcohol; however, due to the small size and fragility of the valves, it was not possible to identify them taxonomically during that period. Subsequently, in November 2019 and 2021, sightings of bivalves attached to algae continued in the same area, and more individuals with similar characteristics were collected in June 2022. Regarding the groins along the Tuxpan coast, during searches for sponges and sea slugs between May and June 2023 (De la Cruz-Francisco 2025a, b), more bivalves were sighted associated with macroalgae. These sightings continued in July 2024, and more individuals were collected for taxonomic identification (Fig. 1).
All bivalves collected and identified as E. vexillum were preserved in ethyl alcohol and deposited in the biological collection of the Facultad de Ciencias Biológicas y Agropecuarias of the Universidad Veracruzana. The taxonomic identification of the species was based on the size, coloration, and morphology of the valves; these characteristics were described in Borrero and Díaz (1998), Çevik et al. (2008), and Tëmkin (2010). The updated taxonomic classification for the species was verified in MolluscaBase (MolluscaBase 2025). A Nikon optical microscope (Tokyo, Japan) was used to observe the external characteristics of the valves. Length and height measurements of the collected bivalves were also recorded. From these data, the following descriptive parameters of the valves were calculated: minimum, mean ± standard deviation, and maximum.
RESULTS
A total of 54 E. vexillum individuals were collected. In the rocky intertidal zone of Cazones, 7 individuals were collected in November 2018 on green algae, and 12 more individuals were collected in June 2022 (20°44′20.22″N, 97°11′36.36″W), associated with 2 species of green algae. Finally, 35 specimens were collected in July 2024 on the coastal groins of Tuxpan, Veracruz (20°59′34.38″N, 97°19′8.25″W), associated with several species of green and red algae. Regarding bivalve dimensions, the average length and height values were slightly higher in the few individuals collected along the Cazones coast in November 2018, compared to those of individuals collected in June 2022 and July 2024 (Table 1). The systematics, morphology, and habitat of E. vexillum are described below.
Table 1 Comparative data of the length and height of Electroma vexillum collected on the coasts of Tuxpan and Cazones, Veracruz, Mexico. Descriptive measures: minimum, mean ± standard deviation, maximum, and number of individuals (n).
| Cazones 2018 (n = 7) | Cazones 2022 (n = 12) | Tuxpan 2024 (n = 35) | |||||||
| Dimensions | Minimum | Mean ± SD | Maximum | Minimum | Mean ± SD | Maximum | Minimum | Mean ± SD | Maximum |
| Length | 4.2 | 4.7 ± 0.3 | 5.2 | 3.9 | 4.5 ± 0.4 | 5.3 | 2.7 | 4.2 ± 0.8 | 5.6 |
| Height | 2.8 | 3.7 ± 0.6 | 4.3 | 2.4 | 3.8 ± 0.6 | 4.5 | 1.7 | 3.1 ± 0.7 | 4.5 |
Systematics
Order Ostreida Férussac, 1822. Family Vulsellidae Gray, 1854. Genus Electroma Stoliczka, 1871. Species Electroma vexillum (Reeve, 1857). Synonyms: Avicula vexillum Reeve, 1857.
Morphology
Small, wedge-shaped bivalves, measuring 2.5 to 5.6 mm in length and 1.7 to 4.5 mm in height (Table 1). The valves are fragile, thin, and convex, and lacked auricles, scales, and radial ribs. Concentric lines can be seen in some individuals (Fig. 2a, b). The valves are unequal (inequivalve); the ventral valve is smaller and slightly convex, with a groove toward the posterior margin; the dorsal valve is larger and convex anteriorly, with an umbonal carina (Fig. 2c, d). The umbo is elevated and displaced forward (prosocline). The hinge is straight, but toothless. In most specimens, external coloration was amber with a dark brown zigzag stripe pattern. Some individuals had a brown band toward the posterior part of the umbonal carina (Fig. 2a). The specimens had fibrous filaments that attach them to benthic algae fronds (Fig. 2d, e).

Figure 2 Two individuals of Electroma vexillum collected from benthic marine algae on the groins of Tuxpan, Veracruz. The first individual is 4 mm long: dorsal valve (a) and ventral valve (b). The second individual is 3 mm long: dorsal valve (c) and ventral valve (d). The byssus filaments are visible (white arrow). Aggregation of E. vexillum on red algae (e).
Habitat
All bivalves were found associated with different species of green and red algae (Fig. 3). In the Tuxpan coastal groins, E. vexillum was observed as an epibiont on 4 species of green algae: Ulva lactuca Linnaeus, 1753 (Fig. 3a); Bryopsis pennata J.V. Lamouroux, 1809 (Fig. 3b); Caulerpa sertulariodes (S.G. Gmelin) M. Howe, 1905 (Fig. 3c); and Caulerpa mexicana Sonder ex Kützing, 1849 (Fig. 3d). In addition, it was observed as an epibiont on 7 species of red algae: Acanthophora spicifera (M. Vahl) Børgesen, 1910 (Fig. 3e); Alsidium triquetrum (S.G. Gmelin) Trevisan, 1845; Hypnea musciformis (Wulfen) J.V. Lamouroux, 1813; Agardhiella subulata (C. Agardh) Kraft & M.J. Wynne, 1979; Gracilaria blodgettii Harvey, 1853; Gracilaria domingensis Sonder ex Dickie, 1874 (Fig. 3f), and Solieria filiformis (Kützing) P.W. Gabrielson, 1985 (with high abundance) (Fig. 3g-h). Meanwhile, in the rocky intertidal zone of Cazones, aggregations of E. vexillum were recorded on 2 species of green algae: U. lactuca and Caulerpa racemosa. It is worth mentioning that other groups of invertebrates associated with benthic algae, such as hydras, polychaetes, pycnogonids, amphipods, isopods, tanaidaceans, decapods, gastropods, bryozoans, and ascidians, were also observed.

Figure 3 Aggregations of Electroma vexillum on benthic marine algae on the groins of the Tuxpan coast, Veracruz, Mexico, in July 2024. Benthic algae species: Ulva lactuca (a), Bryopsis plumosa (b), Caulerpa sertularioides (c), Caulerpa mexicana (d), Acantophora specifiera (e), Gracilaria dominguensis (f), and Solieria filiformis (g-h).
DISCUSSION
Morphologically, E. vexillum is distinguished from species native to the Gulf of Mexico by the absence of hinge teeth, auricles, scales, and radial ribs, characteristics that are present in species of the genera Isognomon, Pteria, Ischadium, and Modiolus, which are larger in size (12-75 mm) and adhere mainly to rocks, mangrove roots, oysters, and octocorals (García-Cubas and Reguero 2007). On the other hand, individuals of E. vexillum are small (<6 mm) and were found exclusively in benthic macroalgae. This type of association has also been reported in the southern Caribbean (Borrero and Díaz 1998), the Mediterranean Sea (Bakır et al. 2012), and in India, where E. vexillum has also been found associated with seagrasses (Asha et al. 2016). In addition, E. vexillum has been reported to colonize rocks and artificial substrates in Colombia and Turkey, the Mediterranean Sea (Borrero and Díaz 1998, Bakır et al. 2012), and soft bottoms in Israel (Albano et al. 2024).
Although E. vexillum abundance was not estimated in this work, it is important to note that dense aggregations were observed on red and green algae on the Tuxpan coast (July 2024), which could signal the invasive capacity of this exotic bivalve in the region. A similar case was reported for the species Electroma sp. in the southern Caribbean, where it was considered to hinder the collection of seeds from commercially important bivalves in the Colombian region (Borrero and Díaz 1998). The presence of E. vexillum could alter the habitat due to competition with the native benthic community, which is composed of diverse groups of invertebrates, such as hydras, polychaetes, pycnogonids, crustaceans, and gastropods, which are also associated with native benthic algae.
Electroma vexillum was found exclusively on intertidal benthic algae growing on artificial structures (groins) and natural rocky promontories (rocky intertidal zones). However, this does not rule out the possibility that the species also occurs in deeper coastal waters, as it has been reported at depths of 2-10 m in other locations (Borrero and Díaz 1998) and up to 20 m (Albano et al. 2024). Furthermore, its dispersion most likely expands in other ecosystems of the northern region of Veracruz, such as coral reefs located 12 km from the coast, where benthic algae are common and abundant (González-González et al. 2016, De la Cruz-Francisco et al. 2020).
Maritime transport has been suggested as the main route of introduction of E. vexillum into the Mediterranean Sea (Çevik et al. 2008, Albano et al. 2024) and of Electroma sp. into the southern Caribbean (Borrero and Díaz 1998). The accidental introduction of E. vexillum into the Gulf of Mexico is most likely linked to maritime transport, which is frequent along the Tuxpan coast due to its proximity to the seaport (6 km away), where national and international vessels dock. Maritime navigation has also been linked to the dispersal of other exotic species into the Gulf of Mexico, such as the encrusting coral Oculina patagonica Angelis D’Ossat, 1908 and the regal demoiselle fish Neopomacentrus cyanomos (Bleeker, 1856), which are established on the coasts of Tuxpan and Cazones and have spread along the coast of Veracruz (González-Gándara and De la Cruz-Francisco 2014; De la Cruz-Francisco et al. 2015, 2017; Colín-García et al. 2018; Rodríguez-Muñoz et al. 2023).
The preference of E. vexillum for inhabiting different species of benthic algae could be an indication of its invasive potential. This threat could extend throughout the Gulf of Mexico, given that benthic algae are widely distributed in this region (Mateo-Cid et al. 2024). However, the invasive capacity of E. vexillum and its effect on native species are still unknown; therefore, it is necessary to analyze its impact on the morphology, growth, photosynthesis, diversity, and abundance of basibiont algae. Furthermore, the community structure of the benthic fauna associated with these algae should be evaluated to determine possible changes driven by the presence of this exotic species. In addition, studies of the biology of E. vexillum directed toward topics related to its life cycle, reproductive period, population growth, dispersal mechanisms, and tolerance to environmental factors are essential.
Finally, it is important to recognize that the identification of this exotic species was based exclusively on the external morphology of the valves, using morphological characteristics that coincide with the taxonomic descriptions of E. vexillum. Therefore, it is essential to conduct molecular analyses, such as sequencing of the 18S rRNA and COI (cytochrome c oxidase subunit 1) genes. These techniques will allow us to more precisely validate the identity of E. vexillum, identify its geographic origin, and determine the routes of its introduction into the Gulf of Mexico.
CONCLUSIONS
After 6 years of observations and records in the coasts of Tuxpan and Cazones, Veracruz, E. vexillum is considered an established exotic species in the Gulf of Mexico. Thus, it is essential to conduct studies focusing on its ecology and distribution in the region because it inhabits intertidal benthic algae, which serve as habitat for many native marine invertebrates and epiphytic algae. Furthermore, molecular analyses are needed to validate the identification of this exotic species with greater precision and determine its geographic origin.

