












 (pdf)
(pdf)




 SciELO
SciELO  SciELO
SciELO 
 Permalink
Permalink
Introducción
El sistema de producción de aves de traspatio se caracteriza por el manejo a pequeña escala de gallinas, pavos, guajolotes y otras aves de diferente edad dentro o alrededor del patio familiar (Pym 2010). Este sistema, común en comunidades rurales e indígenas, integra actividades agrícolas y pecuarias con el objetivo de proveer alimentos básicos para el autoconsumo (Cuca-García et al. 2018). Dentro de este modelo productivo destacan las aves criollas, también denominadas locales, de rancho o autóctonas, que se caracterizan por su adaptabilidad al entorno, su capacidad de sobrevivir con pocos insumos y su resiliencia frente a variaciones en la disponibilidad de alimento (Zaragoza et al. 2011). Estas aves, además de carecer de un patrón fenotípico definido, cumplen un papel relevante en las prácticas culturales y sociales de las comunidades.
A pesar de la amplia presencia de gallinas criollas en el medio rural mexicano, persiste un conocimiento limitado sobre sus condiciones de crianza y características fenotípicas. El interés creciente por estas aves ha motivado estudios que documentan su diversidad morfológica y subrayan la necesidad de profundizar en su investigación. Ejemplos de ello incluyen investigaciones de Luis-Chincoya et al. (2016), Hernández-Ortega et al. (2017, 2020) y Vega et al. (2018), quienes proponen investigaciones de tipo molecular para identificar genes asociados con adaptación y productividad. Este enfoque permitiría desarrollar líneas genéticamente rústicas, reforzando la conservación de sistemas tradicionales, cultural y socialmente sostenibles (Zaragoza et al. 2013, Camacho-Escobar et al. 2016).
En el estado de Guerrero, el gobierno ha implementado programas sociales dirigidos a combatir la inseguridad alimentaria mediante el impulso de la producción pecuaria (Hersch-Martínez y Pisanty-Alatorre 2016). Entre estas acciones, la introducción de aves de alta producción, a través de paquetes tecnológicos, ha desplazado los recursos genéticos locales, generando erosión genética al sustituir genotipos adaptados por razas exóticas. En este contexto los estudios de caracterización fenotípica son fundamentales para documentar la variabilidad morfológica presente en las poblaciones criollas, lo cual constituye un primer paso hacia su conservación y aprovechamiento sustentable. La caracterización permite identificar atributos asociados con la adaptación local, rusticidad y potencial productivo, ofreciendo una base para programas de mejora genética orientados al contexto rural. Ante este panorama, el objetivo del presente estudio fue caracterizar fenotípicamente a las gallinas criollas de traspatio criadas en comunidades indígenas del municipio de Ayutla de los Libres, Guerrero, México.
Materiales y métodos
Ubicación del estudio
El estudio se realizó en la región Costa Chica del estado de Guerrero, en cinco comunidades del municipio de Ayutla de los Libres (Atocutla, El Camalote, El Timbre, Plan de Gatica, Tecomulapa), participantes de manera voluntaria (Figura 1). El municipio posee una superficie de 1 030 km2. Aproximadamente, 40% de la población del municipio puede considerarse indígena, en la que predomina la lengua mixteca y tlapaneca (INEGI 2020). El clima predominante es cálido subhúmedo con lluvias en verano, con temperatura media anual de 28 °C; las lluvias se presentan de junio a octubre, con mayor intensidad en septiembre, la precipitación oscila entre 1 500 y 2 000 mm (INEGI 2021).
Población animal
Dado el desconocimiento del número de gallinas y unidades de producción presentes en las comunidades participantes, se utilizó un muestreo no probabilístico tipo bola de nieve (Hernández et al. 2014). La recolección de datos se llevó a cabo de agosto 2021 a julio 2022, considerando un total de 419 gallinas adultas (107 machos y 312 hembras) criadas en 149 unidades de producción de traspatio. No se excluyeron animales en función de su edad o etapa productiva; sin embargo, únicamente se incluyeron individuos que presentaban un desarrollo anatómico completo. La alimentación de las gallinas estaba basada en maíz blanco y amarillo, producto de las siembras bajo condiciones de temporal, y se complementa con residuos de otros productos como calabaza, ajonjolí, masa, desechos de cocina, más los insectos que las gallinas pudieran cazar y plantas silvestres que cosechen durante el día.
El número de aves muestreadas por comunidad fue: 65 gallinas en Atocutla, 73 en El Camalote, 39 en El Timbre, 190 en Plan de Gatica y 52 en Tecomulapa. Cabe destacar que este estudio se realizó con la participación voluntaria de los productores; en cada unidad de producción, los propietarios seleccionaron de manera aleatoria los animales a evaluar a partir de las aves disponibles en sus traspatios.
Características evaluadas
El peso vivo (PV, kg) de los animales se determinó utilizando una báscula de mesa con capacidad de 20 kg (NOVAL modelo ECO 15TN; precisión: 0.001 kg). Las variables circunferencia de pechuga (CP), longitud del cuerpo (LC), de ala (LA), de tarso (LT), y de muslo (LM) se midieron con una cinta métrica flexible y se expresaron en cm, siguiendo las recomendaciones de la FAO (2012). Para reducir posibles sesgos, todas las mediciones fueron realizadas por un responsable con el apoyo de dos asistentes.
Las medidas corporales se definieron según Fayeye et al. (2006) y Francesch et al. (2011): CP, circunferencia de la región pectoral a la altura de la quilla, pasando la cinta métrica detrás de las alas; LC, distancia desde la base del pico hasta el extremo caudal, sin considerar plumas; LA, desde la articulación de la escápula, hasta el extremo distal del ala; LT, longitud desde la articulación tibio-tarsiana hasta la punta del tarso, manteniendo los dedos a 90° respecto al mismo; y LM, longitud desde la tibia hasta la articulación femoral. Mediante observación directa (FAO 2012), también se registraron el color de plumaje, tarsos y piel, así como el tipo de cresta.
Análisis estadístico
Todos los análisis estadísticos se realizaron con el programa R (R Core Team 2023). Las variables morfométricas (PV, LC, CP, LA, LT, LM) se analizaron bajo un diseño de bloques completamente al azar. El modelo estadístico fue:
Donde: yijk, Variable de respuesta; μ, Media general; βj, Efecto del i-ésimo sexo (i = macho, hembra); τi, Efecto de la j-ésima comunidad (j = Atocutla, El Camalote, El Timbre, Plan de Gatica, Tecomulapa); y εijk, Error aleatorio.
Los valores promedio se compararon mediante la prueba de Tukey (α = 0.05). La armonía morfológica de las aves se evaluó a través de coeficientes de correlación de Pearson, calculados de forma independiente para machos y para hembras, y posteriormente considerando el conjunto total de gallinas evaluadas. Asimismo, se realizó un análisis de componentes principales (ACP) utilizando la instrucción prcomp, considerando todas las aves evaluadas. Se analizó el porcentaje de varianza explicada para identificar el número de componentes relevantes. Para las variables cualitativas (color de plumaje, tarsos y piel, y tipo de cresta) fue empleada la estadística descriptiva para determinar la dispersión de los datos obtenidos.
Resultados
Los valores promedio ± error estándar de las variables morfométricas estudiadas, agrupados por sexo, se presentan en la Tabla 1. El efecto del sexo resultó significativo (p < 0.05) en todas las características analizadas, con los machos mostrando mayor peso y dimensiones corporales en comparación con las hembras. Las medidas de las hembras fueron entre 8.3 y 34.1% menores que las de los machos. En todas las variables, los coeficientes de variación (CV) superaron el 10%, indicando heterogeneidad en la población estudiada.
Tabla 1 Efecto del sexo sobre el peso vivo y medidas corporales de gallinas de traspatio de comunidades indígenas
| Variable | Machos | Hembras | |||
|---|---|---|---|---|---|
| Prom ± EE | CV (%) | Prom ± EE | CV (%) | ||
| Peso vivo (kg) | 2.24 ± 0.06a | 27.38 | 1.67 ± 0.02b | 23.45 | |
| Longitud del cuerpo (cm) | 44.89 ± 0.57a | 13.22 | 40.41 ± 0.39b | 16.84 | |
| Circunferencia de la pechuga (cm) | 29.56 ± 0.58a | 20.13 | 27.30 ± 0.32b | 20.39 | |
| Longitud del ala (cm) | 19.87 ± 0.33a | 16.99 | 17.19 ± 0.16b | 15.97 | |
| Longitud del tarso (cm) | 11.35 ± 0.17a | 15.56 | 9.57 ± 0.08b | 15.61 | |
| Longitud del muslo (cm) | 13.22 ± 0.28a | 22.21 | 11.78 ± 0.15b | 21.83 | |
Prom: Promedio; EE: Error estándar; CV: Coeficiente de variación. a, b Medias con distinta literal por fila son diferente estadísticamente (p < 0.05).
La influencia de la localidad sobre las variables morfométricas evaluadas se resume en la Tabla 2. Excepto para CP, todas las variables evaluadas fueron significativamente (p < 0.05) afectadas por el sitio de origen. Las aves de “El Camalote” presentaron los valores más altos de PV, LC, CP y LT, mientras que las mayores medidas de LA y LM se observaron en “El Timbre” y “Tecomulapa”, respectivamente. El factor de la localidad explicó hasta 17.6% de la variabilidad observada.
Tabla 2 Efecto de la comunidad sobre el peso vivo y medidas corporales de gallinas de traspatio de comunidades indígenas
| Variable | Atocutla | Camalote | El Timbre | Plan de Gatica | Tecomulapa |
|---|---|---|---|---|---|
| Promedio ± Error Estándar | |||||
| PV (kg) | 1.77 ± 0.07ab | 1.94 ± 0.05a | 1.78 ± 0.09ab | 1.83 ± 0.04ab | 1.65 ± 0.06b |
| LC (cm) | 39.70 ± 0.61b | 43.56 ± 1.22a | 41.21 ± 1.08ab | 41.69 ± 0.44ab | 40.79 ± 0.64ab |
| CP (cm) | 27.36 ± 0.81a | 28.92± 0.76a | 26.96 ± 0.94a | 27.69 ± 0.39a | 28.46 ± 0.68a |
| LA (cm) | 18.28 ± 0.40ab | 18.03 ± 0.38ab | 18.91 ± 0.49a | 17.66 ± 0.23ab | 17.14 ± 0.37b |
| LT (cm) | 9.26 ± 0.16c | 10.59 ± 0.23a | 10.04 ± 0.32ab | 10.22 ± 0.12a | 9.44 ± 0.22bc |
| LM (cm) | 11.99 ± 0.26ab | 12.23 ± 0.31ab | 11.33 ± 0.35b | 12.20 ± 0.22ab | 12.66 ± 0.37a |
PV: Peso vivo; LC: Longitud del cuerpo; CP: Circunferencia de la pechuga; LA: Longitud del ala; LT: Longitud del tarso; LM: Longitud del muslo. a, b Medias con distinta literal por fila son diferente estadísticamente (p < 0.05).
Los coeficientes de correlación de Pearson entre las variables morfométricas, estimados por separado para machos y hembras se presentan en la Tabla 3. En los machos, el 80% de las correlaciones fueron significativas, de las cuales 17% mostro una asociación fuerte (r > 0.5), el 50% moderada (r > 0.3 r ≤ 0.5) y el 33% débil (r ≤ 0.3). Las correlaciones más consistentes se observaron entre el PV, LC y LT, reflejando un patrón armónico en la estructura corporal. En las hembras, el 67% de las correlaciones fueron significativas, observando asociaciones de intensidad débil (75%) y moderada (25%). Estos resultados sugieren una menor uniformidad estructural en las hembras en comparación con los machos.
Tabla 3 Coeficientes de correlación de Pearson para variables morfológicas cuantitativas para machos (arriba de la diagonal) y hembras (debajo de la diagonal)
| PV | LC | CP | LA | LT | LM | |
|---|---|---|---|---|---|---|
| PV | - | 0.51** | 0.17NS | 0.45** | 0.59** | 0.31* |
| LC | 0.30** | - | 0.28* | 0.27* | 0.49** | 0.15NS |
| CP | 0.16* | 0.31** | - | -0.14NS | 0.22* | 0.31* |
| LA | 0.17* | 0.10NS | -0.21* | - | 0.33* | -0.20* |
| LT | 0.26** | 0.29** | 0.17* | 0.23** | - | 0.31* |
| LM | 0.11NS | 0.00NS | 0.10NS | -0.35** | 0.06NS | - |
PV: Peso vivo; LC: Longitud del cuerpo; CP: Circunferencia de la pechuga; LA: Longitud del ala; LT: Longitud del tarso; LM: Longitud del muslo. * (p < 0.05); **(p < 0.01); NS(p > 0.05).
La Tabla 4 muestra los coeficientes de correlación de Pearson estimados para la población total evaluada. Estos resultados confirman el patrón de armonía corporal previamente identificado al analizar los datos por sexo. En conjunto, las correlaciones presentaron magnitudes de medianas a bajas (<0.55) y fueron estadísticamente significativos. Del total de asociaciones significativas, el 60% correspondió a relaciones débiles, 33% a moderadas y el 7% restante a fuertes. Las correlaciones más consistentes se observaron nuevamente entre el PV, LC y LT, lo que resalta la estrecha relación entre estas dimensiones en la población estudiada.
Tabla 4 Coeficientes de correlación de Pearson para variables morfológicas cuantitativas de gallinas de traspatio (datos agrupados).
| PV | LC | CP | LA | LT | LM | |
|---|---|---|---|---|---|---|
| PV | - | 0.43** | 0.22** | 0.41** | 0.52** | 0.27** |
| LC | - | 0.34** | 0.23** | 0.42** | 0.10* | |
| CP | - | -0.11* | 0.24** | 0.19** | ||
| LA | - | 0.39** | -0.19* | |||
| LT | - | 0.22** | ||||
| LM | - |
PV: Peso vivo; LC: Longitud del cuerpo; CP: Circunferencia de la pechuga; LA: Longitud del ala; LT: Longitud del tarso; LM: Longitud del muslo. * (p < 0.05); **(p < 0.01); NS(p > 0.05).
El análisis de componentes principales mostró que los dos primeros componentes explican conjuntamente el 61.7% de la variación total observada en las gallinas de traspatio (CP1 = 39.7% y CP2 = 22.0%). En el biplot presentado en la Figura 2, el CP1 está principalmente asociado con PV y LT, lo que indica que estos atributos son determinantes en la variabilidad morfométrica. Por su parte, el CP2 está definido principalmente por la LA y LM, características que reflejan diferencias en el desarrollo de las extremidades.
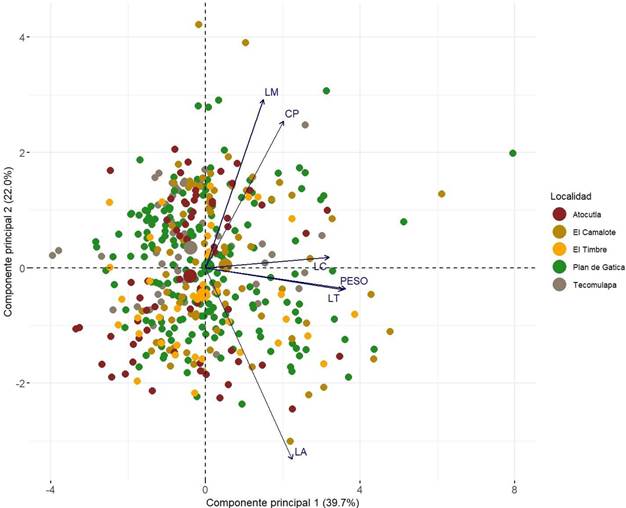
Figura 2. Distribución de individuos y variables morfométricas en el espacio definido por los dos primeros componentes principales. PV: Peso vivo; LC: Longitud del cuerpo; CP: Circunferencia de la pechuga; LA: Longitud del ala; LT: Longitud del tarso; LM: Longitud del muslo
La distribución del color de plumaje, tarsos y piel en la población evaluada se detalla en la Tabla 5. Más de la mitad de las aves presentaron plumaje oscuro, principalmente negro y rojo; en machos predomino el color rojo, mientras que en hembras fue el negro. Aproximadamente una quinta parte de la población, tanto en hembras como en machos, exhibió plumaje blanco. En cuanto a los tarsos, el color amarillo fue el más frecuente. Alrededor del 70% de las aves presentó piel blanca. En términos generales, el color azul en plumaje, tarsos y piel se registró en baja frecuencia.
La Tabla 6 presenta las frecuencias de los distintos tipos de cresta observados en las gallinas de traspatio. La cresta simple o sencilla mostró dominancia.
Tabla 5 Color de plumaje, tarsos y piel en gallinas de traspatio en comunidades indígenas del municipio de Ayutla de los Libres, Guerrero.
| Variable | Categoría | General (%) | Fm (%) | Fh (%) |
|---|---|---|---|---|
| Color de plumaje | Blanco | 20.29 | 18.7 | 20.8 |
| Trigueño | 18.38 | 18.7 | 18.3 | |
| Rojo | 26.25 | 35.5 | 23.1 | |
| Azul | 9.79 | 7.5 | 10.6 | |
| Negro | 25.30 | 19.6 | 27.2 | |
| Color de los tarsos | Amarillo | 47.49 | 71.0 | 39.4 |
| Blanco | 20.53 | 13.1 | 23.1 | |
| Negro | 10.74 | 2.8 | 13.5 | |
| Verde | 17.66 | 11.2 | 19.9 | |
| Azul | 3.58 | 1.9 | 4.2 | |
| Color de piel | Amarillo | 24.34 | 34.6 | 20.8 |
| Blanco | 68.50 | 63.6 | 70.2 | |
| Azul y/o negro | 7.16 | 1.9 | 9.0 |
Fm (%): Frecuencia en machos; Fh (%): Frecuencia en hembras
Tabla 6 Tipo de cresta en gallinas de comunidades indígenas del municipio de Ayutla de los Libres, Guerrero
| Categoría | General (%) | Fm (%) | Fh (%) |
|---|---|---|---|
| Sencilla o Simple | 89.02 | 86.0 | 90.1 |
| Guisante | 4.30 | 4.7 | 4.2 |
| Rosa | 2.63 | 4.7 | 1.9 |
| Nuez, cojín o fresa | 3.56 | 4.6 | 3.2 |
| En V | 0.48 | . | 0.6 |
Fm (%): Frecuencia en machos; Fh (%): Frecuencia en hembras
Discusión
Considerando un nivel de significancia del 5%, la FAO (2012) recomienda entre 100 y 300 hembras y alrededor de 30 machos maduros para estudios de caracterización fenotípica. En el presente estudio, el número de gallinas y gallos muestreados superó esta recomendación, respaldando la representatividad de los resultados para la región evaluada. No obstante, futuras investigaciones con mayores tamaños de muestra podrían aportar estimaciones más robustas y minimizar los sesgos asociados a la heterogeneidad poblacional y al método de muestreo. Los valores obtenidos (Tabla 1) sugieren que las aves evaluadas pertenecen a un biotipo medio, de origen mediterráneo, alemanes y holandeses (Campo 2009), con PV promedio de alrededor de 2.5 kg en machos y de 2.0 kg para hembras. Este fenotipo es característico de poblaciones de doble propósito, útiles tanto para producción de huevo como de carne (Montes et al. 2019). Desde una perspectiva biológica y funcional, dimensiones como la CP y LT no sólo son indicadores de desarrollo somático, sino también de la capacidad para soportar demandas energéticas asociadas a la postura o engorda. Estas medidas pueden relacionarse con la eficiencia metabólica y a la capacidad locomotora, rasgos críticos en sistemas extensivos donde las aves deben recorrer distancias para obtener su alimento. El CV refleja la heterogeniedad morfoestructural (Herrera y Luque 2009); los valores relativamente altos observados en la Tabla 1 indican una población fenotípicamente diversa, situación esperable en sistemas de traspatio sin programas formales de selección o mejoramiento genético (Villacís et al. 2016). Esta variabilidad, lejos de ser un inconveniente, puede constituir una ventaja adaptativa, ya que asegura la presencia de genotipos con distintos grados de resiliencia frente a factores ambientales y sanitarios, contribuyendo así a la estabilidad productiva en contextos rurales. El dimorfismo sexual observado - machos de mayor preso y dimensiones que hembras, es un fenómeno universal en aves (Espinosa et al. 2021), atribuido a la acción diferencial de andrógenos y a estrategias reproductivas en las que los machos más grandes pueden tener ventajas en competencia sexual (Montes-Vergara et al. 2022). En término productivos, este rasgo puede ser aprovechado en programas de selección para optimizar la producción cárnica en machos y la eficiencia reproductiva en hembras.
Las diferencias entre localidades (Tabla 2) parecen asociarse a la adaptación de las aves a microclimas locales. En Ayutla, el clima cálido subhúmedo con temperaturas de hasta 34 °C (CONAGUA 2022) podría afectar el crecimiento, ya que a partir de los 24.3 °C se activan mecanismos fisiológicos de termorregulación que elevan las demandas energéticas y reducen el consumo de alimento (Pishnamazi et al. 2015). Aves de comunidades al norte, donde las temperaturas son hasta 2 °C menores, presentan un desarrollo somático superior, lo que sugiere que se encuentran en un rango térmico más favorable. Estas diferencias reflejan la interacción genotipo-ambiente y subrayan la importancia de considerar factores ambientales en estrategias de conservación y mejoramiento.
En sistemas de traspatio, la alimentación de las aves se basa en productos agrícolas locales como maíz blanco y amarillo, residuos de calabaza y ajonjolí, además de desechos de cocina. Esta dieta, si bien refleja prácticas culturales arraigadas suele ser nutricionalmente deficiente (Luis-Chincoya et al. 2016), limitando el potencial productivo. Las diferencias en frecuencia y cantidad de alimento ofrecido, así como el acceso a forraje y fauna local, pueden contribuir a la variabilidad intra e intercomunidad. El escaso uso de alimento balanceado, motivado por costos y disponibilidad, refuerza la dependencia de recursos locales y la necesidad de aves rústicas con alta capacidad de adaptación.
El patrón estructural observado en las hembras (débilmente armónico, Tabla 3 y 4) sugiere una morfología más orientada a la reproducción que a la producción de carne, mientras que los machos muestran un modelo medianamente armónico, con mayor potencial cárnico. Esta información es relevante para diseñar programas de selección comunitarios enfocados en mejorar características funcionales sin comprometer la adaptabilidad local.
Respecto a los caracteres cualitativos, el predominio de plumajes oscuros (rojo y negro) coincide con estudios previos en poblaciones de traspatio (Andrade-Yucailla et al. 2018, Montes et al. 2019) y podría tener una función adaptativa. Plumajes oscuros incrementan la resistencia al calor y la protección frente a la radiación solar (Akumbugu et al. 2023), factores críticos en climas cálidos. Por el contrario, la presencia de plumaje blanco puede reflejar introgresión genética de estirpes comerciales (Cobb, Ross), derivada de la compra de aves para engorda o de programas gubernamentales como PESA (SAGARPA 2013). Este fenómeno representa un riesgo para la conservación del germoplasma local, ya que podría reducir la rusticidad y la diversidad genética afectando la capacidad de las aves para sobrevivir en condiciones de traspatio.
La coloración amarilla predominante en tarsos y piel probablemente se asocia al consumo de maíz amarillo rico en carotenoides, cuyos metabolitos se depositan en tegumentos. Finalmente, la alta frecuencia de cresta simple, además de coincidir con estudios en otras poblaciones rurales (Apuno et al. 2010, Picardal et al. 2015, Andrade-Yucailla et al. 2017, 2018), podría tener ventajas funcionales. Este tipo de cresta se ha relacionado con mayor fertilidad y una mejor capacidad de disipar calor en ambientes cálidos, favoreciendo la adaptación y supervivencia.
La caracterización de estas poblaciones no solo aporta información biológica y productiva, sino que es esencial para estrategias de conservación in situ y para el diseño de programas de mejoramiento participativo. Mantener estas poblaciones adaptadas a sistemas de traspatio también contribuye a la seguridad alimentaria y a la preservación de prácticas culturales locales, donde la cría de gallinas no solo tiene valor económico, sino también social y simbólico en las comunidades rurales.
Conclusiones
Las gallinas pueden considerarse de talla mediana con variabilidad en todas las características, siendo afectadas por el sexo y la localidad de origen. En general, en machos predomina un modelo moderadamente armónico, en tanto que el modelo para las hembras es débilmente armónico. El peso vivo y la longitud del tarso fueron las variables que más influyeron sobre el tamaño. Abundan las aves de plumaje oscuro con tarsos de color amarillo, piel blanca y cresta simple. Los resultados obtenidos podrían ser base para el desarrollo de programas de conservación, mejoramiento genético y establecer planes de manejo productivo, potencializando la adaptación de la población avícola estudiada, sin embargo, se deberán desarrollar más estudios que relacionen los niveles productivos con el componente genético para logran un mayor aprovechamiento de los recursos animales de las comunidades participantes.

