












 (pdf)
(pdf)




 SciELO
SciELO  SciELO
SciELO 
 Permalink
Permalink
Introducción
La producción de hongos comestibles del género Pleurotus conocidos como setas, ocupa el segundo lugar en la oferta mundial, Pleurotus cornucopie, P. eryngii, P. nebrodensis y P. ostreatus son las especies más cultivadas (Pérez-Moreno et al. 2021). En México, la producción y comercialización de hongos es principalmente por el género de Agaricus, con el 95.3% de la oferta nacional seguido de Pleurotus spp. con 4.6% (Mayett y Martínez-Carrera 2019), esta actividad productiva la desarrollan grandes y pequeños productores, quienes en su mayoría se concentran en estados del centro del país, aunque se han reportado módulos de producción de características rusticas en la mayoría de los estados (Royse y Sánchez 2017). Los hongos Pleurotus son un alimento de alto valor por sus propiedades nutricionales y medicinales que promueven la salud humana (Mayett y Martínez-Carrera 2019), son ricos en proteínas cruda (21-27%), fibras (13-19%), potasio, calcio, magnesio, cobre, hierro, zinc, así como polifenoles con actividad antioxidante (Vega et al. 2022).
De las especies de Pleurotus cultivadas mundialmente destaca P. ostreatus (Royse y Sánchez 2017); sin embargo, estudios revelan que esta especie requiere de ciertas condiciones controladas de temperatura. Por ejemplo, en el desarrollo del hongo en la etapa de incubación se requieren temperaturas entre 24 y 25 °C (Jaramillo-Mejía y Albertó 2019, Roggero-Luque et al. 2021, Roblero-Mejía et al. 2021); mientras que, en la fructificación diversos autores han reportado distintos rangos de temperaturas, de 17 a 20 °C (Abid et al. 2020, Roggero-Luque et al. 2021), de 18 a 20 °C (Jaramillo-Mejía y Albertó 2013), de 20 a 25 °C (Ejigu et al. 2022) y de 22 a 26 °C (Roblero-Mejía et al. 2021). Teniendo en cuenta que, en México existen pequeños productores con módulos de producción de características rústicas y escasamente tecnificados, es difícil mantener el control de las condiciones de temperatura antes mencionadas para el cultivo de P. ostreatus (Huacash-Pale y Ocampo-Guzmán 2021). En la región centro de Veracruz se presentan condiciones de clima cálido húmedo (Am) y cálido subhúmedo (Aw) con temperaturas promedio de 27 a 32 °C en los meses cálidos (García 2004). Por lo tanto, esto limita la posibilidad de cultivar especies comerciales de P. ostreatus. Cepas de P. djamor serían una opción de cultivo cuando se excede de 27 °C, se ha demostrado que esta especie de Pleurotus se desarrolla adecuadamente en condiciones agroclimáticas de tipo tropical; además, de que por sus características genéticas tiene ciclos cortos de cultivo (Salmones 2017).
En México, P. djamor es un recurso genético abundante con respecto a otras especies de Pleurotus y al ser un recurso local presenta mejores adaptaciones que las cepas comerciales procedentes de otras latitudes (Salmones et al. 2022). Así mismo, son abundantes los rastrojos de cultivos agrícolas, los cuales poseen alto potencial para su uso como sustrato en el cultivo de Pleurotus, como residuos del cultivo de la caña de azúcar, rastrojo de maíz, pulpa de café, hojas de plátano, rastrojo de frijol (INEGI 2023). En Veracruz se han realizado producciones exitosas de P. pulmonarius y P. ostreatus en rastrojo de maíz y paja de avena (Gaitán-Hernández y Silva-Huerta 2016). La región de Las Montañas, Veracruz, está siendo atendida mediante transferencias de tecnología y capacitaciones a productores en el cultivo de cepas silvestres de Pleurotus y uso de sustratos locales (Aguilar-Ventura et al. 2024); por tanto, identificar recursos genéticos de Pleurotus y sustratos productivos beneficiará la toma de decisiones en este sistema agroalimentario.
El sistema agroalimentario de Pleurotus contribuye a la seguridad alimentaria de los pueblos, ya que se desarrolla como una actividad económica que genera ingresos y posee alto potencial para la diversificación de las actividades agrícolas, por la reutilización de residuos que se generan después de la cosecha (Huacash-Pale y Ocampo-Guzmán 2021). Los hongos del género Pleurotus tienen la capacidad de desarrollarse en diversos sustratos lignocelulósicos como subproductos agrícolas, agroindustriales y forestales, de estos, en México se reciclan anualmente 500 000 toneladas (Piña-Guzmán et al. 2016). Todo ello contribuye en el reciclaje de este tipo de materiales orgánicos, de los cuales, es posible obtener un alimento funcional de forma sostenible (Salmones et al. 2022). Ante el uso eficiente de estos recursos, en el cultivo de setas, se busca una alta productividad en términos del peso fresco de esporomas, eficiencia biológica, tasa de producción, rendimiento y ciclos cortos de producción, con el sustrato empleado, el cual se puede utilizar solo o combinado (Roblero-Mejía et al. 2021). Para iniciar un sistema productivo de Pleurotus, donde se empleen como insumos cepas silvestres y sustratos locales, es necesario medir la velocidad de crecimiento micelial ya que esto permitirá predecir la capacidad del hongo en formar micelio vegetativo y la posterior formación de cuerpos fructíferos que tendría en un sustrato determinado (León-Avendaño et al. 2013). Por lo anterior, el objetivo del presente estudio fue evaluar la velocidad de crecimiento micelial y la productividad de cepas silvestres de Pleurotus djamor en sustratos locales de la región centro del estado de Veracruz, México.
Materiales y métodos
El estudio se realizó en el Laboratorio de Biotecnología de Cultivo de Hongos Comestibles de la Facultad de Ciencias Biológicas y Agropecuarias, Universidad Veracruzana, región Orizaba-Córdoba, Veracruz, México.
Material biológico
Se emplearon tres cepas silvestres de Pleurotus djamor (MXLD23, MXLD24 y 598), las cuales, fueron recolectadas en 2016 en los municipios de Amatlán de los Reyes y Fortín de las Flores, Veracruz, México, estos municipios se encuentran entre los 720 y 1 000 msnm, temperatura promedio de 18 a 18.8 °C, precipitación de 1 807 a 1 832 mm, con vegetación de bosque templado caducifolio y selva mediana subperennifolia. Actualmente las cepas se encuentran resguardadas en el banco de germoplasma del Laboratorio de Biotecnología de Cultivo de Hongos Comestibles y Medicinales de la Facultad de Ciencias Biológicas y Agropecuarias, Universidad Veracruzana, región Orizaba-Córdoba, Veracruz, México (Figura 1).

Evaluación de la velocidad de crecimiento de las colonias fúngicas
El crecimiento micelial de cada cepa, se determinó en seis tratamientos, con cuatro repeticiones. Las unidades experimentales (UE) fueron placas de Petri de 9 cm de diámetro que contenían 10 g (peso húmedo) de los siguientes sustratos que se consideraron como los tratamientos: paja de cebada (PC), hoja de caña de azúcar (HCA), hojas y tallos de Arachis pintoi (Fabaceae) (AP), rastrojo de maíz (RM), hojas de plátano (HP) y 10 mL de Papa Dextrosa Agar (PDA) (tratamiento control) (Figura 1 e, g, i, f, h y d). La humedad en los sustratos se ajustó al 65% y se esterilizaron en autoclave durante 15 min a 121 °C y 1 055 kg cm-2 de presión. Cuando las UE se enfriaron y permanecieron a temperatura ambiente (25 ± 2 °C), estas fueron inoculadas con un fragmento de 1 cm de diámetro de PDA que contenía micelio de cada cepa desarrollada durante 20 días (Gaitán-Hernández 2021). Posteriormente, las UE se incubaron en oscuridad a 25 ± 2 °C (Jaramillo-Mejía y Albertó 2013), en un periodo de 15 días. Durante la incubación; cada 24 h se midió el diámetro de crecimiento de la colonia fúngica empleando un vernier Mitutoyo® modelo estándar. Se registraron dos mediciones sobre dos rectas perpendiculares (A y B) que previamente se habían trazado en las placas de Petri; para el trazo de las rectas se tomó como punto de intersección el centro de la caja de Petri donde fue colocado el inóculo. La velocidad de crecimiento se expresó en cm día-1, bajo la expresión matemática:
Donde: VC = es la velocidad de crecimiento, Ø = diámetro de crecimiento de la colonia y t = tiempo (Campi et al. 2016).
Evaluación de la productividad
La productividad de esporomas de cada cepa (MXLD23, MXLD24 y 598) se evaluó en cinco formulaciones (tratamientos) de sustrato con cinco repeticiones: PC (100%), HCA (100%), RM (100%), HCA (90%) + AP (10%) y PC (80 %) + HP (20%). El sustrato AP no se usó al 100%, porque en pruebas anteriores se observó que tiende a compactarse y el sustrato HP genera mayor espacio entre partículas. Por lo que, las formulaciones evaluadas permitieron un ajuste adecuado del umbral de humedad requerido en el experimento que fue de 65%.
Los sustratos de cada formulación, se trituraron de forma manual a un tamaño de 2 a 5 cm; con el cual, se formaron paquetes de 1 kg empleando bosas de polietileno. Los paquetes se colocaron dentro de un contenedor cilíndrico de metal que contenía 20 L de agua y después de 24 h se aplicó calor durante 2 h empleando gas LP como combustible. En el interior del contenedor se generó vapor de agua a temperatura de 95 ± 2 °C. Posteriormente, los paquetes se enfriaron a temperatura ambiente y se inocularon con 50 g de semillas de sorgo colonizadas por cada cepa antes mencionada. Los paquetes se incubaron a temperatura ambiente y en ausencia de luz (dos semanas). Cuando se observó una colonización micelial completa y formación de primordios, los paquetes fueron puestos en una cámara de fructificación (Romero-Arenas et al. 2018), la cual se mantuvo a temperatura ambiente y a una humedad controlada del 95%.
De cada paquete, se realizaron tres cosechas de esporomas durante un ciclo de producción (Ejigu et al. 2022), se empleó una balanza de precisión Highland®-HCB1002 para cuantificar el peso fresco total de esporomas (PFTE), PFTE = suma del peso fresco de los esporomas de cada cosecha en gramos, el ciclo de producción (CP) se representó como el total de días transcurridos desde la inoculación del hongo hasta el final de la tercera cosecha (Romero-Arenas et al. 2018). La eficiencia biológica (EB), la tasa de producción (TP) y el rendimiento (R) se calcularon mediante las siguientes fórmulas (Roblero-Mejía et al. 2021):
Diseño de los experimentos y análisis de datos
Los experimentos fueron establecidos mediante un diseño completamente aleatorizado, modelado por: Yij = µ + Ti + Eij. Para cada i = 1, …, n y j = 1, …, n, donde Yij (variable dependiente) en el primer experimento fue el valor de la velocidad de crecimiento micelial (cm día-1) y en el segundo experimento fueron las variables (PFTE, CP, EB, TP y R) del tratamiento i en su j-ésima repetición. μ: es la media general de la variable de respuesta. Ti: es el efecto del tratamiento i. Eij: es el error experimental asociado al tratamiento i en su j-ésima repetición.
Los valores obtenidos de la velocidad de crecimiento micelial (cm día-1), se analizaron mediante un análisis de varianza no paramétrico y con regresión lineal simple (Coello-Loor et al. 2017), y con el método de mínimos cuadrados se obtuvieron los coeficientes del modelo: la pendiente (m) y el intercepto (b) con su respectivo límite inferior (LI) y límite superior (LS), el coeficiente de determinación R2 y el p-valor, con una confianza del 95 % (Balzarini et al. 2008).
Por la naturaleza de las variables estudiadas (continuas) que tienden a ser de distribución normal, se evaluaron con una prueba de Shapiro-Wilks a excepción de la variable CP (variable discreta) (Zar 2010). Las variables que cumplieron con el supuesto de normalidad (Lambda de Wilks > = 0.9 y p-unilateral (D) > 0.05; se les aplicó un análisis de varianza (ANOVA) con α de 0.05 de confianza y contraste de medias de Tukey. Las variables que no cumplieron con el supuesto de normalidad requerido (Lambda de Wilks > = 0.9 y p-unilateral (D) > 0.05) se sometieron a un análisis de varianza no paramétrico de Kruskal-Wallis y prueba de medias de rangos pareadas (p ≤ 0.05). El efecto de las formulaciones de sustratos sobre la EB, R y CP se analizó mediante un análisis de componentes principales (Balzarini et al. 2008). En las salidas estadísticas se muestran los valores: grados de libertad (Gl), para igualdad de varianzas (F), estadístico de prueba de Kruskal-Wallis (H) y p-valor. Los análisis se realizaron con el software estadístico InfoStat® Versión 2020 (Di-Rienzo et al. 2020).
Resultados
Velocidad de crecimiento de la cepa MXLD23
En la Figura 2, se observa el diámetro de crecimiento micelial de la cepa MXLD23 durante 15 días de incubación. Después de la inoculación de la cepa en los medios evaluados, la etapa de latencia se observó hasta el segundo día, en el cual, la cepa mostró diferente adaptación respecto al diámetro de crecimiento en cada tratamiento, se encontraron diferencias Gl = 5, H = 42.57 y p-valor = <0.0001) (Tabla 1). La etapa exponencial fue entre el tercer y sexto día (Figura 2); en esta etapa, el crecimiento de la cepa en los tratamientos RM y HP fue mayor y estadísticamente diferente al resto de los tratamientos: día 3 (Gl = 5, H = 41.31 y p-valor = < 0.0001), día 4 (Gl = 5, H = 40.79 y p-valor = < 0.0001), día 5 (Gl = 5, H = 42.76 y p-valor = < 0.0001) y día 6 (Gl = 5, H = 44.21 y p-valor = < 0.0001) (Tabla 1). Las UE con HP, fueron colonizadas por la cepa en siete días, con RM en ocho días, con PDA en nueve días, con AP en 11 días y con HCA en 14 días. Las UE con PC no fueron colonizadas completamente alcanzando un crecimiento máximo de 7.3 cm de diámetro (Figura 2).
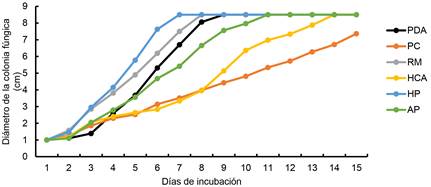
Figura 2 Velocidad de crecimiento (cm día-1) de la cepa MXLD23. PDA: papa dextrosa agar, PC: paja de cebada, RM: rastrojo de maíz, HCA: hoja de caña de azúcar, HP: hoja de plátano y AP: Arachis pintoi. Media de ocho mediciones.
Tabla 1 Diámetro de crecimiento (cm día-1) de la cepa MXLD23 Pleurotus djamor durante ocho días. Prueba no paramétrica de Kruskal Wallis.
| Tratamiento | Día 1 | Día 2 | Día 3 | Día 4 | Día 5 | Día 6 | Día 7 | Día 8 |
|---|---|---|---|---|---|---|---|---|
| PDA | 1 ± 0a | 1.1 ± 0d | 1.3 ± 0.06c | 2.6 ± 0.15cd | 3.6 ± 0.3b | 5.3 ± 0.34bc | 6.7 ± 0.5bc | 8.1 ± 0.5ab |
| PC | 1 ± 0a | 1.3 ± 0.07bc | 1.8 ± 0.13bc | 2.3 ± 0.2d | 2.5 ± 0.32c | 3.1 ± 0.26de | 3.5 ± 0.27de | 3.9 ± 0.32c |
| RM | 1 ± 0a | 1.5 ± 0.09a | 2.8 ± 0.21a | 3.8 ± 0.29ab | 4.9 ± 0.5ab | 6.1 ± 0.54ab | 7.5 ± 0.36ab | 8.5 ± 0a |
| HCA | 1 ± 0a | 1.2 ± 0cd | 2.1 ± 0.12b | 2.4 ± 0.14d | 2.6 ± 0.13c | 2.8 ± 0.15e | 3.3 ± 0.19e | 3.9 ± 0.42c |
| HP | 1 ± 0a | 1.4 ± 0.07ab | 2.9 ± 0.21a | 4.1 ± 0.25a | 5.7 ± 0.15a | 7.6 ± 0.24a | 8.5 ± 0a | 8.5 ± 0a |
| AP | 1 ± 0a | 1.1 ± 0d | 2.04 ± 0.14b | 2.8 ± 0.1bc | 3.5 ± 0.24b | 4.6 ± 0.47cd | 5.4 ± 0.7cd | 6.6 ± 1.14bc |
Los valores representan la media de ocho repeticiones ± desviación estándar. Medias con diferente letra dentro de la columna son significativamente diferentes (p ≤ 0.05). PDA: papa dextrosa agar, PC: paja de cebada, RM: rastrojo de maíz, HCA: hojas de caña de azúcar, HP: hojas de plátano y AP: Arachis pintoi.
El mayor diámetro promedio de crecimiento de la cepa MXLD23, fue en el sustrato HP con 1.23 cm día-1, seguido del RM con 1.11 cm día-1. En el medio PDA, el crecimiento fue de 1.07 cm día-1, en AP de 0.83 cm día-1, en PC de 0.43 cm día-1 y en HCA de 0.4 cm día-1. El modelo de regresión, fue significativo (p ≤ 0.05) y mostró un coeficiente de determinación R2 en intervalo de 0.95 a 0.99 (Tabla 2). De acuerdo a LS de m, en diámetro la cepa podría incrementar en PDA, PC, RM, HCA, HP y AP con crecimiento de 0.26, 0.003, 0.008, 0.007, 0.21 y 0.11 cm día-1, respectivamente (Tabla 2).
Tabla 2 Análisis de regresión lineal simple de la velocidad de crecimiento (cm día-1) de la cepa MXLD23 Pleurotus djamor. m: pendiente, b: intercepto, LI: límite inferior, LS: límite superior y R2: coeficiente de determinación
| Tratamiento | M | b | LI de m | LS de m | LI de b | LS de b | R2 | p-valor |
|---|---|---|---|---|---|---|---|---|
| PDA | 1.07 | -1.11 | 0.82 | 1.33 | -2.39 | 0.17 | 0.95 | < 0.0001 |
| PC | 0.43 | 0.51 | 0.4 | 0.46 | 0.36 | 0.67 | 0.99 | < 0.0001 |
| RM | 1.11 | -0.45 | 1.03 | 1.19 | -0.85 | -0.05 | 0.99 | < 0.0001 |
| HCA | 0.4 | 0.61 | 0.34 | 0.47 | 0.27 | 0.96 | 0.97 | < 0.0001 |
| HP | 1.23 | -0.54 | 1.02 | 1.44 | -1.6 | 0.52 | 0.97 | < 0.0001 |
| AP | 0.83 | -0.34 | 0.72 | 0.94 | -0.91 | 0.24 | 0.98 | < 0.0001 |
PDA: papa dextrosa agar, PC: paja de cebada, RM: rastrojo de maíz, HCA: hojas de caña de azúcar, HP: hojas de plátano y AP: Arachis pintoi.
Velocidad de crecimiento de la cepa MXLD24
El crecimiento micelial de la cepa MXLD24 se muestra en la Figura 3. La etapa de latencia de la cepa se observó hasta las 48 h después de la inoculación de la cepa en los sustratos, a excepción del tratamiento control PDA que fue hasta el día 4, se encontraron diferencias (GL = 5, H = 41 y p-valor = < 0.0001) (Tabla 3). Esta cepa colonizó completamente las placas de Petri que contenían: HP en un tiempo de 7 días, RM en 8 días, HCA en 12 días y el tratamiento control PDA en 14 días. Mientras que, las placas con PC y AP no fueron colonizadas por completo después de los 15 días que duró el experimento (Figura 3). En el día 7 de la evaluación, se observó una sobresaliente colonización de la cepa en los tratamientos HP y RM a diferencia del resto de tratamientos, se encontraron diferencias (Gl = 5, H = 44.18 y p-valor = < 0.0001). (Tabla 3).
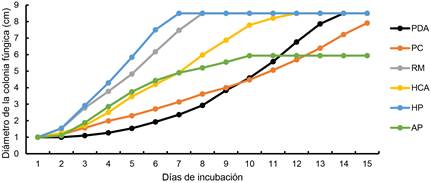
Figura 3 Velocidad de crecimiento (cm día-1) de la colonia fúngica de la cepa MXLD24. Media de ocho mediciones. PDA: papa dextrosa agar, PC: paja de cebada, RM: rastrojo de maíz, HCA: hoja de caña de azúcar, HP: hoja de plátano y AP: Arachis pintoi.
Tabla 3 Diámetro de crecimiento (cm día-1) de la cepa MXLD24 Pleurotus djamor durante ocho días. Prueba no paramétrica de Kruskal Wallis.
| Tratamiento | Día1 | Día 2 | Día 3 | Día 4 | Día 5 | Día 6 | Día 7 | Día 8 |
|---|---|---|---|---|---|---|---|---|
| PDA | 1 ± 0a | 1 ± 0c | 1.1 ± 0c | 1.3 ± 0.1c | 1.54 ± 0.1d | 1.9 ± 0.1d | 2.4 ± 0.2d | 2.9 ± 0.3c |
| PC | 1 ± 0a | 1.1 ± 0.1b | 1.6 ± 0.1b | 1.9 ± 0.3bc | 2.3 ± 0.4cd | 2.7 ± 0.5cd | 3.1 ± 0.4cd | 3.6 ± 0.5bc |
| RM | 1 ± 0a | 1.5 ± 0.1a | 2.8 ± 0.4a | 3.8 ± 0.4a | 4.8 ± 0.4ab | 6.2 ± 0.6ab | 7.5 ± 0.4ab | 8.5 ± 0.0a |
| HCA | 1 ± 0a | 1.2 ± 0.1ab | 1.7 ± 0.3b | 2.5 ± 0.5b | 3.4 ± 0.5bc | 4.2 ± 0.7bc | 4.9 ± 0.8bc | 5.9 ± 1.2b |
| HP | 1 ± 0a | 1.5 ± 0.2a | 2.9 ± 0.4a | 4.3 ± 0.5a | 5.8 ± 0.5a | 7.5 ± 0.4a | 8.5 ± 0a | 8.5 ± 0a |
| AP | 1 ± 0a | 1.1 ± 0bc | 1.6 ± 0.3b | 2.5 ± 0.4b | 3.5 ± 0.4bc | 4.2 ± 0.5bc | 4.8 ± 0.4bc | 5.2 ± 0.3b |
Los valores representan la media de ocho repeticiones ± desviación estándar. Medias con diferente letra dentro de la columna son significativamente diferentes (p ≤ 0.05). PDA: papa dextrosa agar, PC: paja de cebada, RM: rastrojo de maíz, HCA: hojas de caña de azúcar, HP: hojas de plátano y AP: Arachis pintoi.
El mayor promedio de crecimiento de la cepa MXLD24 se observó en el sustrato HP con 1.22 cm día-1; mientras que, el menor valor fue en el tratamiento control PDA con 0.27 cm día-1 (Tabla 4). El valor de R2 del modelo evaluado resultó en intervalo de 0.9 a 0.99. De acuerdo a LS de m, la velocidad de crecimiento de la colonia fúngica de la cepa MXLD24, podría incrementar en PDA, PC, RM, HCA, HP y AP con crecimiento promedio de 0.1, 0.03, 0.09, 0.11, 0.2 y 0.11 cm día-1 respectivamente. Las tasas de crecimiento en HP y RM superaron tres veces la tasa de crecimiento la cepa en PC.
Tabla 4 Análisis de regresión lineal simple de la velocidad de crecimiento (cm día-1) de la cepa MXLD24 Pleurotus djamor. m: pendiente, b: intercepto, LI: límite inferior, LS: límite superior y R2: coeficiente de determinación
| Tratamiento | m | b | LI de m | LS de m | LI de b | LS de b | R2 | p-valor |
|---|---|---|---|---|---|---|---|---|
| PDA | 0.27 | 0.4 | 0.18 | 0.37 | -0.06 | 0.87 | 0.9 | 0.0003 |
| PC | 0.38 | 0.48 | 0.35 | 0.41 | 0.31 | 0.64 | 0.99 | < 0.0001 |
| RM | 1.11 | -0.51 | 1.03 | 1.2 | -0.96 | -0.06 | 0.99 | < 0.0001 |
| HCA | 0.73 | -0.17 | 0.63 | 0.84 | -0.69 | 0.35 | 0.98 | < 0.0001 |
| HP | 1.22 | -0.49 | 1.03 | 1.42 | -1.48 | 0.5 | 0.97 | < 0.0001 |
| AP | 0.68 | 0.09 | 0.57 | 0.79 | -0.46 | 0.63 | 0.98 | < 0.0001 |
PDA: papa dextrosa agar, PC: paja de cebada, RM: rastrojo de maíz, HCA: hojas de caña de azúcar, HP: hojas de plátano y AP: Arachis pintoi.
Velocidad de crecimiento de la cepa 598
Las curvas de crecimiento micelial de la cepa 598 en los diferentes sustratos evaluados, se muestran en la Figura 4. La etapa de latencia de la cepa, se observó durante los primeros dos días posteriores de la inoculación del hongo en los tratamientos PDA, RM, HCA, HP y AP, mientras que en PC fue hasta el día 3, se observaron diferencias (GL = 5, H = 39.68 y p-valor = < 0.0001) Tabla (5). Las placas de Petri que contenían RM, HP y medio PDA fueron colonizadas en un tiempo de 8 días (Figura 4), estos fueron significativamente diferentes al resto de tratamientos (GL = 5, H = 39.18 y p-valor = < 0.0001) Tabla (5). El desarrollo micelial a los 15 días en AP mostró un máximo crecimiento de 5.05 cm de diámetro, en PC el crecimiento máximo fue de 4.7 cm y en HCA 3.5 cm (Figura 4).
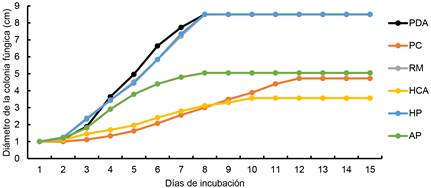
Figura 4 Velocidad de crecimiento (cm día-1) de la colonia fúngica de la cepa 598. Media de ocho mediciones. PDA: papa dextrosa agar, PC: paja de cebada, RM: rastrojo de maíz, HCA: hoja de caña de azúcar, HP: hoja de plátano y AP: Arachis pintoi.
Tabla 5 Diámetro de crecimiento (cm día-1) de la cepa 598 Pleurotus djamor durante ocho días. Prueba no paramétrica de Kruskal Wallis.
| Tratamiento | Día1 | Día 2 | Día 3 | Día 4 | Día 5 | Día 6 | Día 7 | Día 8 |
|---|---|---|---|---|---|---|---|---|
| PDA | 1 ± 0a | 1.1 ± 0.1ab | 1.9 ± 0.3ab | 3.6 ± 0.2a | 4.9 ± 0.2a | 6.6 ± 0.2a | 7.7 ± 0.4a | 8.5 ± 0a |
| PC | 1 ± 0a | 1 ± 0.0c | 1.1 ± 0.1c | 1.3 ± 0.1d | 1.6 ± 0.2d | 2.1 ± 0.3d | 2.6 ± 0.4c | 3 ± 0.4b |
| RM | 1 ± 0a | 1.2 ± 0.1a | 2.4 ± 0.4a | 3.4 ± 0.7ab | 4.5 ± 0.6ab | 5.8 ± 0.6ab | 7.2 ± 0.3ab | 8.5 ± 0a |
| HCA | 1 ± 0a | 1.1 ± 0bc | 1.5 ± 0.2bc | 1.7 ± 0.2cd | 1.9 ± 0.3cd | 2.4 ± 0.4cd | 2.8 ± 0.5c | 3.1 ± 0.6b |
| HP | 1 ± 0a | 1.2 ± 0.1a | 2.3 ± 0.2a | 3.4 ± 0.3ab | 4.4 ± 0.5ab | 5.8 ± 0.9ab | 7.3 ± 0.4a | 8.5 ± 0.0a |
| AP | 1 ± 0a | 1.2 ± 0a | 1.8 ± 0.1b | 2.9 ± 0.1bc | 3.8 ± 0.1bc | 4.4 ± 0.1bc | 4.8 ± 0.2bc | 5.1 ± 0.2b |
Los valores representan la media de ocho repeticiones ± desviación estándar. Medias con diferente letra dentro de la columna son significativamente diferentes (p ≤ 0.05). PDA: papa dextrosa agar, PC: paja de cebada, RM: rastrojo de maíz, HCA: hojas de caña de azúcar, HP: hojas de plátano y AP: Arachis pintoi.
A partir de la velocidad de crecimiento de la cepa 598, se determinó que el mayor valor promedio fue en el medio PDA (1.2 cm día-1), seguido de HP y RM con crecimientos de 1.13 y 1.12 cm día-1 respectivamente. El valor R2 del modelo resultó en intervalo de 0.92 a 0.98 y un p-valor ≤ 0.05 (Tabla 6). Los sustratos PC y HCA fueron menos eficientes en el desarrollo del hongo, presentando una velocidad de crecimiento promedio de 0.3 cm día-1. De acuerdo a LS de m, la velocidad de crecimiento de la colonia fúngica, podría incrementar en RM y HP un crecimiento promedio de 0.14 cm día-1 respectivamente (Tabla 6).
Tabla 6 Análisis de regresión lineal simple de la velocidad de crecimiento (cm día-1) de la cepa MXLD24 Pleurotus djamor. m: pendiente, b: intercepto, LI: límite inferior, LS: límite superior y R2: coeficiente de determinación
| Tratamiento | m | b | LI de m | LS de m | LI de b | LS de b | R2 | p-valor |
|---|---|---|---|---|---|---|---|---|
| PDA | 1.2 | -1 | 1.01 | 1.39 | -1.94 | -0.01 | 0.98 | < 0.0001 |
| PC | 0.3 | 0.38 | 0.21 | 0.38 | -0.06 | 0.81 | 0.92 | 0.0002 |
| RM | 1.12 | -0.8 | 0.98 | 1.26 | -1.48 | -0.06 | 0.98 | < 0.0001 |
| HCA | 0.31 | 0.53 | 0.27 | 0.35 | 0.33 | 0.73 | 0.98 | < 0.0001 |
| HP | 1.13 | -0.8 | 0.98 | 1.27 | -1.55 | -0.04 | 0.98 | < 0.0001 |
| AP | 0.65 | 0.17 | 0.54 | 0.77 | -0.41 | 0.76 | 0.97 | < 0.0001 |
PDA: papa dextrosa agar, PC: paja de cebada, RM: rastrojo de maíz, HCA: hojas de caña de azúcar, HP: hojas de plátano y AP: Arachis pintoi.
Productividad de las cepas MXLD23, MXLD24 Y 598
La EB de la cepa MXLD23 fue estadísticamente similar en las formulaciones: F2, F3 y F4, pero diferente en F1 y F5 (Gl = 4, F = 17.1 y p-valor = < 0.0001). En la F2, se encontró la mayor TP (2.22% día-1), R (6.66%), PFTE (187.8 g) y el menor CP (34.2 días) (Tabla 7). Las formulaciones de sustrato F1 y F5 presentaron los mayores ciclos de producción 46.4 y 48.6 días respectivamente, sin embargo, esto no se reflejó en una mayor productividad en términos de EB.
Tabla 7 Indicadores de producción de las cepas de Pleurotus djamor (MXLD23, MXLD24 y 598). Eficiencia biológica (EB), tasa de producción (TP), rendimiento (R), peso fresco total de esporomas (PFTE) y ciclo de producción (CP).
| Formulaciones de sustratos | Cepa | EB (%) | TP (%/día) | R (%) | PFTE (g) | CP (días) |
|---|---|---|---|---|---|---|
| F1 | MXLD23 | 28.91 ± 5.48b | 0.51 ± 0.1c | 3.58 ± 0.68c | 101.2 ± 19.18b | 46.2 ± 2.39b |
| F2 | 53.66 ± 1.96a | 2.22 ± 0.08a | 6.66 ± 0.24a | 187.8 ± 6.87a | 34.2 ± 3.03a | |
| F3 | 47.03 ± 8.13a | 1.67 ± 0.29a | 4.85 ± 0.84b | 164.6 ± 28.45a | 34.8 ± 2.68a | |
| F4 | 50.29 ± 5.07a | 1.47 ± 0.15ab | 5.15 ± 0.52b | 176 ± 17.76a | 35.8 ± 1.3a | |
| F5 | 36.17 ± 5.53b | 0.67 ± 0.1bc | 5.55 ± 0.85ab | 126.6 ± 19.36b | 48.6 ± 2.88b | |
| F1 | MXLD24 | 26.11 ± 1.33D | 0.46 ± 0.02C | 3.23 ± 0.17A | 91.4 ± 4.67D | 56.4 ± 0.89C |
| F2 | 46.09 ± 2.27A | 1.9 ± 0.09A | 5.72 ± 0.28B | 161.3 ± 7.95A | 24.2 ± 2.68A | |
| F3 | 32.11 ± 4.26C | 1.14 ± 0.15AB | 3.31 ± 0.44A | 112.4 ± 14.93C | 28.2 ± 5.54A | |
| F4 | 45.43 ± 4.19A | 1.33 ± 0.12AB | 4.66 ± 0.43AB | 159 ± 14.68A | 34.2 ± 4.32AB | |
| F5 | 38.4 ± 1.14B | 0.71 ± 0.02BC | 5.89 ± 0.17B | 134.4 ± 3.97B | 54.2 ± 0.45BC | |
| F1 | 598 | 40.67 ± 4.42 c | 0.72 ± 0.08 c | 5.04 ± 0.55 d | 109.4 ± 11.89 c | 53.2 ± 2.59 b |
| F2 | 42.28 ± 3.77 c | 1.75 ± 0.16 ab | 5.25 ± 0.47 cd | 181.8 ± 16.21 b | 39.8 ± 3.11 a | |
| F3 | 59.05 ± 4.96 b | 2.09 ± 0.18 a | 6.09 ± 0.51 c | 224.4 ± 18.85 a | 40.6 ± 1.52 a | |
| F4 | 74.07 ± 4.12 a | 2.17 ± 0.12 a | 7.59 ± 0.42 b | 240 ± 13.34 a | 41.6 ± 2.79 a | |
| F5 | 59.71 ± 4.68 b | 1.10 ± 0.09 bc | 9.16 ± 0.72 a | 166.6 ± 13.07 b | 52 ± 1 b |
Los valores representan la media de cinco repeticiones ± desviación estándar. Medias con diferente estilo de letra dentro de la columna son significativamente diferentes (p ≤ 0.05). F1: paja de cebada (100 %). F2: hoja de caña de azúcar (100 %). F3: hojas de caña de azúcar (90 %) + Arachis pintoi (10 %). F4: rastrojo de maíz (100 %). F5: paja de cebada (80 %) + hoja de plátano (20 %). Los valores de TP y CP de las tres cepas corresponde a la prueba de Kruskal-Wallis, así mismo los valores de R para la cepa MXLD24.
Los valores de EB correspondientes a la cepa MXLD24 se agruparon en cuatro grupos estadísticos: A, B, C y D (Gl = 4, F = 42.03 y p-valor = <0.0001) (Tabla 7). La F2 produjo la mayor EB (46%), seguido de F4 con una EB de 45.4%; ambas formulaciones fueron estadísticamente similares. La menor EB fue en F1 con una EB de 26.11. La TP para la F2 fue de 1.9% día-1, mientras que para F1 fue de 0.46% día-1. EL R en la F2, F4 y F5 fue estadísticamente igual con valores de 5.72, 4.66 y 5.89%, respectivamente. Los valores de PFTE para F2 fue de 161.3 g y para F4 fue de 159 g, estos fueron superiores a las demás formulaciones evaluadas. El menor CP fue en F2 con una duración promedio de 24.2 días y el mayor CP fue en PC con 56.4 días. En cada uno de los parámetros productivos evaluados de la cepa MXLD24 se encontraron diferencias TP: Gl = 4, H = 22.24 y p-valor = 0.0002; R: Gl = 4, H = 21.63 y p-valor = 0.0003; PFTE: Gl = 4, F = 42.03 y p-valor = < 0.0001; CP: Gl = 4, H = 21.04 y p-valor = 0.0003.
Los valores de la productividad de la cepa de 598, obtenidos en las diferentes formulaciones de sustrato, se muestran en la Tabla 4. El mayor valor de EB (74%) fue en F4, este valor fue significativamente diferente (Gl = 4, F = 49.46 y p-valor = < 0.0001) frente a las otras formulaciones de sustrato evaluadas. La TP fue estadísticamente similar en F4, F3 y F2 con valores promedio de 2.17, 2.09 y 1.75% día-1 respectivamente. Un R de 9.16% fue en F5 el cual fue superior a las demás formulaciones evaluadas, esto representa una mayor bioconversión de esporomas (base seca) de la cepa en el sustrato empleado. El mayor PFTE revelado fue en F4 Y F3 con valores de 240 y 224 g respectivamente, esta ganancia de peso representa el doble del PFTE obtenido en F1 (109 g). El menor CP se encontró en F2 presentando una duración promedio de 39.8 días. Se encontraron diferencias significativas (p ≤ 0.05) entre los parámetros productivos evaluados.
El análisis de componentes principales mostró que el 95.9% de la variación producida es explicada por el componente principal 1 y 2. El vector EB presentó mayor aporte al componente principal 1 y el vector CP al componente 2 (Figura 5). Un efecto positivo se encontró entre la cepa 598 cultivada en F4 y F3 en relación a la EB. La EB de la cepa MXLD23 fue sobresaliente en F2. La cepa 598 cultivada en F5 se caracterizó fuertemente con el R, esto representó mayor ganancia en peso seco de esporomas de la productividad obtenida. Las cepas MXLD24 cultivada en F5, MXLD23 cultivada en F5 y 598 cultivada en F1, se caracterizaron por presentar mayores ciclos de producción. La cepa MXLD24 cultivada en F2 y la cepa MXLD23 cultivada en F4 presentaron una relación directa al presentar los menores ciclos de producción.
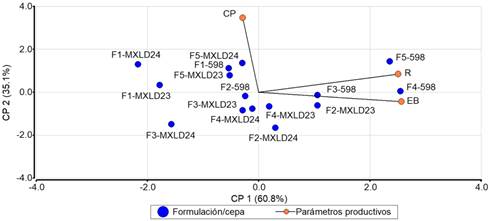
Figura 5 Efecto de las formulaciones de sustrato en la productividad de las cepas MXLD23, MXL24 y 598. Correlación cofenética = 997. F1: paja de cebada (100%). F2: hoja de caña de azúcar (100%). F3: hojas de caña de azúcar (90%) + Arachis pintoi (10%). F4: rastrojo de maíz (100%). F5: paja de cebada (80%) + hoja de plátano (20%). EB: eficiencia biológica. R: rendimiento. CP: ciclo de producción.
Discusión
Velocidad de crecimiento micelial
Las curvas de crecimiento micelial de las cepas estudiadas sobre los diferentes sustratos lignocelulósicos (Figuras 1, 2 y 3), presentaron un adecuado ajuste de crecimiento lineal, con coeficientes de determinación R2 entre 0.9 y 0.99, la forma de crecimiento (radial) fue similar como lo describe Gaitán-Hernández (2021). El desarrollo de las cepas evidenció una etapa de latencia de dos días, esta etapa podría estar asociada a que inicialmente el hongo, sintetiza enzimas para la asimilación de nutrientes (Martínez et al. 2015). La etapa de crecimiento exponencial, se observó entre el tercer y sexto día después de la inoculación de las diferentes cepas en los sustratos, esto concuerda con las características propias del desarrollo microbiano, ya que en esta etapa se alcanza la velocidad máxima de crecimiento de la colonia (Campi et al. 2016). No fue posible determinar la etapa estacionaria del crecimiento de la cepa MXLD23 en HP, RM, PDA, AP y HCA; de la cepa MXLD24 desarrollada en HP, RM, HCA PDA y PC; de la cepa 598 desarrollada en PDA, HP y RM, esto porque la limitante física de la placa de Petri de tamaño de 9 cm de diámetro impidió determinar si la colonia fúngica había llegado a su estado estacionario (Campi et al. 2016). Se observó la etapa estacionaria con una disminución de la velocidad del crecimiento de la cepa MXLD23 en PC; de la cepa MXLD24 en AP; de la cepa 598 en HCA, PC y AP, esta reducción de crecimiento pudo deberse a un agotamiento de nutrientes o por la acumulación de desechos tóxicos (Martínez et al. 2015), también pudo deberse a menor adaptabilidad metabólica del hongo a las fuentes de carbono y nitrógeno presentes en cada sustrato (Coello-Loor et al. 2017). La variada velocidad de crecimiento promedio de las cepas MXLD23, MXLD24 y 598 en los diferentes sustratos, podría estar relacionada con la particular composición química y estructura física de cada sustrato empleado (Vega et al. 2022), como fuentes de carbono, minerales, vitaminas, aminoácidos, nitrógeno y oxígeno (Gaitán-Hernández 2021), así como por la capacidad de síntesis de enzimas de cada cepa para biodegradar los materiales orgánicos empleados (Salmones 2017) o por cualidades genéticas de la cepa (León-Avendaño et al. 2013).
El crecimiento micelial promedio que corresponde al valor de m (pendiente) de la cepa MXLD23 en el medio control PDA fue de 1.07 cm día-1 y para la cepa 598 fue de 1.2 cm día-1, estas tasas de crecimiento fueron similares a las reportadas por Coello-Loor et al. (2017) en la evaluación de P. ostreatus en medio PDA con un valor de m de 0.47 mm h-1 o 1.128 cm día-1; sin embargo, la cepa MXLD24 mostró un valor de m de 0.27 cm día-1. Por su parte Suárez y Holguín (2011) indicaron tasas de crecimiento radial en medio PDA para P. pulmonarius y P. ostreatus de 0.81 y 0.65 mm día-1 respectivamente. El medio de cultivo PDA, por sus propiedades físico-químicas es ampliamente usado para el cultivo de hongos proveyendo un adecuado desarrollo, sin embargo, los estudios mostrados, sugieren un aprovechamiento diferencial de los nutrientes del medio, de acuerdo a la cepa cultivada. Los sustratos RM y HP resultaron con un crecimiento micelial de las cepas evaluadas en un intervalo de 1.11 a 1.23 cm día-1, esto fue similar a lo encontrado en el medio PDA. De acuerdo con Salmones (2017), los hongos comestibles del género Pleurotus, entre estos P. djamor, pertenece a los hongos de pudrición blanca cuya característica particular, es que estos hongos poseen enzimas capaces de degradar la lignocelulosa de donde obtienen fuentes de carbono para su desarrollo. El menor crecimiento promedio de las cepas en el sustrato PC que fue de 0.3 a 0.43 cm día-1 y en el sustrato HCA de 3.3 a 0.7 cm día-1, pudo deberse a una menor disponibilidad de nutrientes en los sustratos (Vega et al. 2022), principalmente fuentes de carbono (Gaitán-Hernández 2021). Los valores de crecimiento micelial en el sustrato AP fueron intermedios respecto a los otros sustratos evaluados con valores entre 0.65 a 0.83 cm día-1; el sustrato AP es una leguminosa y por ende, posee un mayor contenido de proteína frente a los otros sustratos empleados (Romero et al. 2022), sin embargo, esto no favoreció en una mayor velocidad de crecimiento micelial, pero si en una mayor producción de biomasa micelial ya que en este sustrato el micelio fue altamente denso (Gaitán-Hernández 2021).
Productividad de las cepas MXLD23, MXLD24 y 598
La productividad de esporomas de la cepa MXLD23 y MXLD24 fue mayor en F2 mostrando una EB de 53.6 y 46.09%, TP de 2.22 y 1.9% día-1, R de 6.66 y 5.72% PFTE de 187.8 y 161.3 g y CP de 34.2 y 24.2 días respectivamente; para la cepa 598, los valores de productividad más altos fueron en F4 con una EB de 74.07%, TP de 2.17% día-1, R de 7.59% PFTE de 240 g y CP de 41.6 días. Pérez-Chávez et al. (2022) reportaron valores productivos de una cepa de P. djamor cultivada en paja de trigo con EB de 167.2% y TP de 3.49 % día-1, PFTE de 501.6 g y CP de 47.8 días; esta misma cepa cultivada en aserrín de álamo suplementado con salvado de trigo obtuvieron una EB de 109.4%, TP 1.65% día-1, PFTE de 328.3 g y CP de 66.1 días. Por su parte, Roblero-Mejía et al. (2021) indicaron valores productivos de una cepa de P. djamor cultivada en pasto pangola con EB de 38.2% y TP de 0.5% día-1 y en pulpa de café con EB de 20.8% y TP de 0.2% día-1. Estos valores productivos, sugieren una variada bioconversión del sustrato por el hongo en esporomas o cuerpos fructíferos de Pleurotus, lo cual puede deberse a factores intrínsecos como las propiedades genéticas del material biológico empleado (León-Avendaño et al. 2013). Así como propiedades físico químicas del sustrato: como el contenido de nitrógeno y de carbono, pH, humedad, minerales, el tamaño de partícula, secreción enzimática de la cepa para degradar materiales lignocelulósicos; también por factores extrínsecos como la temperatura y humedad ambiental de la cámara de incubación y fructificación, el fotoperiodo, el oxígeno, y la concentración de dióxido de carbono (Bellettini et al. 2019). Además, es posible que las cepas de Pleurotus evaluadas requieran menor contenido de nitrógeno, proteína cruda y relación carbono/nitrógeno para su desarrollo y fructificación, por ejemplo, Romero-Arenas et al. (2018) indicaron una mayor productividad en P. ostreatus usando paja de cebada (EB de 102%) con un contenido de proteína de 5.8% y una menor productividad en rastrojo de maíz (EB de 64.2%) con un contenido de proteína cruda de (4.9%); mientras que Roblero-mejía et al. (2021) obtuvieron una EB de 38% en P. djamor y una EB de 48% en P. ostreatus en un sustrato que contenía 0.5% de nitrógeno y carbono de 51%; por su parte, Vega et al. 2022 reportaron EB de 80 a 96% al cultivar P. djamor en sustratos con un contenido de nitrógeno de 1.4 a 2.2% y relación carbono/nitrógeno de 24 a 37. En el presente estudio, la humedad del sustrato en el momento de la inoculación de las cepas fue de 65%, en el estudio de Pérez-Chávez et al. (2022) la humedad fue de 70% y en lo reportado por Roblero-Mejía et al. (2021) fue de 60%; es posible que una mayor proporción de humedad en el sustrato en la etapa de incubación favorece el desarrollo vegetativo y productividad del hongo, permitiendo mejores condiciones en la biodegradación del sustrato y actividad enzimática (Salmones 2017).
En el cultivo de hongos comestibles del género Pleurotus, las cepas de P. djamor destacan por poseer ciclos cortos de producción, en contraste con cepas de las especies de P. ostreatus y P. pulmonarius (Salmones 2017). El ciclo de producción para las cepas de P. djamor fue de 24.2 a 54.4 días. Al respecto, Abid et al. (2020) cultivaron P. ostreatus sobre paja de arroz, paja de trigo y paja de trigo combinada con Pinus wallichiana en una proporción de 50%, indicaron un CP de 54.5, 47.6 y 62.83 días, respectivamente. Mientras que Jaramillo-Mejía y Albertó (2019) reportan un CP entre 66 a 81 días en la producción de P. ostreatus empleando paja de trigo con diferentes proporciones de inóculo. Por otro lado Rugamas et al. (2022) reportaron un CP de 43 y 48 días en el cultivo de P. tuber-regium y P. sajor-caju. Es evidente que, la especie de P. djamor, presenta periodos más cortos de producción, sin embargo, es notable que este periodo productivo puede verse afectado por el sustrato en el que se cultiva el hongo. El sustrato de hojas de caña de azúcar, mostró los CP más cortos de las cepas evaluadas, lo que sugiere altas expectativas del uso de estos insumos en un sistema productivo de Pleurotus en regiones con clima tropical (Salmones et al. 2022).
La productividad más baja de las cepas estudiadas, encontrada en F1, que corresponde a paja de cebada con EB de 26.1 a 40.6%, pudiera deberse a la porosidad e intercambio de gases que provee el sustrato (Abid et al. 2020), ya que en la formulación F5 que corresponde a paja de cebada (80%) + hoja de plátano (20%) la EB fue de 36.1 a 59.7% y es posible que este incremento productivo se diera por la estructura del sustrato. La EB de la cepa 598 en F2 (hojas de caña de azúcar) fue de 42.2% mientras que en F3 que incluye la combinación de hojas de caña de azúcar (90%) + A. pintoi (10%) fue de 59%. Este incremento productivo pudiera deberse a un mayor aprovechamiento de esta leguminosa y sus fuentes de nitrógeno por el hongo, en la formación de cuerpos fructíferos, ya que en las cepas MXLD23 y MXLD24 la EB no incrementó con el uso de A. pintoi. De acuerdo al análisis de componentes principales, la cepa 598, presentó una relación positiva entre la EB y las formulaciones de sustrato (F3 y F4) el R con F5, esto posiblemente se deba a que la cepa posee a una mayor capacidad de síntesis de enzimas degradadoras de lignocelulosa (Salmones 2017), lo que permite una mayor bioconversión de estos materiales orgánicos en esporomas (Vega et al. 2022).
Conclusiones
El estudio reveló un rápido desarrollo micelial de las cepas MXLD23, MXLD24 y 598 en rastrojo de maíz y hojas de plátano; productividad más alta en la cepa 598 cultivada en rastrojo de maíz con valores de EB de 74%, TP de 2.17% día-1, R de 7.59% y PFTE de 240 g. La paja de cebada combinada con 20% de hoja de plátano incrementó la EB de la cepa 598 en 19%. La leguminosa forrajera Arachis pintoi incrementó la EB de la cepa 598 en 17%; por lo que, el uso de esta leguminosa como suplemento en el cultivo de Pleurotus representa un alto potencial. En la región centro del estado de Veracruz, es viable iniciar un sistema de producción de Pleurotus empleando como insumos cepas silvestres y sustratos locales.

