












 (pdf)
(pdf)




 SciELO
SciELO  SciELO
SciELO 
 Permalink
Permalink
Introducción
La estructura de la comunidad vegetal es uno de los aspectos más importantes para la toma de decisiones en el manejo forestal, ya que de esta depende la productividad y las interacciones intra e interespecíficas dentro de los ecosistemas (Bettinger et al., 2017). Evaluar las diferencias en la estructura de las comunidades forestales es esencial para valorar los cambios que pueden llegar a sufrir estas comunidades por las actividades de gestión (Ramos-Hernández et al., 2024). Los impactos que generan los aprovechamientos forestales son importantes en la modificación de las etapas sucesionales de un ecosistema, debido a que el manejo conlleva a modificaciones significativas en la estructura y la composición de especies (Graciano, 2001). Por lo que el conocer la estructura y composición de los bosques antes de las operaciones del manejo forestal (corta), permite entender los cambios que pueden presentar los componentes bióticos y abióticos de un ecosistema determinado (Luján-Soto et al., 2008).
La complejidad estructural y la diversidad arbórea en la productividad de los ecosistemas forestales generan una sinergia, siendo la diversidad una característica estructural y la variabilidad un elemento de la biodiversidad (Alanís-Rodríguez et al., 2010; Monárrez-González et al., 2018). Los estudios de estructura y biodiversidad generan conocimientos para asegurar de manera perpetua y óptima servicios ecosistémicos para las necesidades de las generaciones futuras, a través de decisiones y actividades pertinentes sobre el manejo forestal (Aguirre-Calderón et al., 2015; Zúñiga-Vásquez et al., 2018; Ramos-Hernández et al., 2024).
Varios trabajos han aportado evidencias de que los esquemas de manejo pueden afectar significativamente la composición de especies (Graciano-Luna, 2001; Corral-Rivas et al., 2005; Solís-Moreno et al., 2006; Castellanos-B, 2008; Návar-Cháidez & González-Elizondo, 2009; Leyva-López et al., 2010; Hernández-Salas et al., 2013; Graciano-Ávila et al., 2017; Silva-González et al., 2021 & Galván-Moreno et al., 2024). La mayoría de estos trabajo se han enfocado en describir la diversidad alfa de manera excelente, sin embargo, en la actualidad, ante los nuevos compromisos internacionales sobre la conservación de la biodiversidad y la incursión para demostrar un buen manejo a través de la certificación forestal, se requiere generar conocimientos sobre aspectos estructurales por unidad de manejo más específicas, considerando los métodos silvícolas y características ecológicas como la orientación geográfica, la pendiente o la altura sobre el nivel del mar. Los estudios para evaluar la diversidad también son muy necesarios para poder comprender las relaciones entre comunidades forestales con las diferentes orientaciones geográficas y los esquemas de manejo forestal que se prescriben. (Flores-Morales et al., 2022).
La región de El Salto, ubicada al sur de la capital del estado de Durango, posee una amplia diversidad vegetal, en la cual los géneros Pinus, Quercus y Juniperus son de gran importancia económica para la región. Los bosques de esta zona se han manejado y aprovechado desde 1926, primero con el Método Mexicano de Ordenación de Montes (MMOM) en 1944 y modificado en 1984 a Método de Ordenación de Bosques Irregulares (MMOBI), aplicando el tratamiento de selección, así como por el Método de Desarrollo Silvícola (MDS) mediante tratamientos de aclareos desde la década de los setentas para tener bosques regulares. Sin embargo, existe la necesidad de evaluar los efectos de los tratamientos silvícolas en la estructura y diversidad de los dos métodos de manejo en función de la orientación geográfica, para comprender de qué manera influyen en la riqueza, abundancia y dominancia de las especies, ya que se argumenta que cuando se aplican tratamientos como aclareos, se eliminan algunas especies raras de poca abundancia y la diversidad se modifica porque esa nueva área es colonizada por las especies dominantes que suelen ser las de mayor interés económico (Silva-González et al., 2021).
Los objetivos del presente estudio fueron evaluar el efecto del MMOBI y del MDS en la estructura horizontal del bosque manejado, mediante la abundancia, frecuencia, dominancia, valor de importancia (IVI), riqueza y diversidad, así como conocer la influencia de la orientación geográfica en la composición de especies en bosques del ejido El Brillante, Pueblo Nuevo, Durango. La hipótesis planteada fue que la estructura y la diversidad es modificada por el Método Mexicano de Ordenación de Bosques Irregulares (MMOBI) con el tratamiento de selección y el Método de Desarrollo Silvícola (MDS) con tratamientos de aclareo y que la orientación geográfica influye en dicha modificación. La información generada contribuirá al conocimiento biológico de los ecosistemas forestales en el Noroeste de México y generará conocimiento para la toma de decisiones del manejo de las masas forestales.
Métodos
Área de estudio
El estudio se realizó en el sistema montañoso de la Sierra Madre Occidental en el estado de Durango, en el ejido El Brillante, del municipio de Pueblo Nuevo, entre las coordenadas geográficas 23°50'44.85" latitud N, 105°22'2.95" longitud O y 23°46'49.38" N, 105°20'28.62" O con una Altura de 2600 m s.n.m. (Figura 1). De acuerdo con la clasificación de Köppen modificada por García (INEGI, 2007), el clima predominante en el área es el tipo C (E) (w), que corresponde a semifrío subhúmedo con lluvias en verano. La precipitación oscila entre los 1 000 a 1 200 mm anuales, con las temperaturas medias anuales de 11.5 ºC, las extremas más frías registradas de 8.3 ºC y las extremas más calurosas de 14.3 ºC (Quiñónez-Barraza et al., 2012). Los tipos de suelo conforman asociaciones entre Regosol, Litosol y Cambisol (UPSE, 2007). Los tipos de vegetación existentes son bosque de pino, bosque de encino, bosque de pino-encino, encino-pino y vegetación secundaria arbustiva en todas las comunidades vegetales (González-Elizondo et al., 2012).
Evaluación en campo
La información dasométrica se obtuvo de 52 sitios de monitoreo forestal de forma circular de 1000 m2, 38 para MMOBI, y 13 de MDS (Tabla 1), ubicados aleatoriamente en diferentes áreas de corta del actual programa de manejo. Los sitios se establecieron donde se aplicaron tratamientos silvícolas con el Método de Desarrollo Silvícola (MDS), incluyendo primero, segundo, tercer aclareo y cortas de liberación, así como en áreas con el Método Mexicano de Ordenación de Bosques Irregulares (MMOBI) bajo el tratamiento de selección. Los sitios se dividieron por orientación geográfica 12 sitios para la orientación Norte, Noreste, Noroeste en el grupo (N NE NO); 12 para Sur, Sureste, Suroeste en el grupo (S SE SO) y 14 para Este-Oeste en otro grupo (EO) para MMOBI, 4 sitios (N NE NO), 8 para (S SE SO) y 4 para (EO) en MDS. Se colectó información silvícola dasométrica de cada sitio, por estratos y por tratamiento. Para los análisis se consideraron el diámetro normal (Dn> 7.5 cm), el cual se midió con cinta diamétrica Ben Meadows; altura total (H), con un clinómetro Suunto Pm5/360pc; diámetro de copa Norte-Sur y Este-Oeste, así como el registro de la especie de cada individuo. Los formatos utilizados fueron los recomendados por el Sistema de Planeación Forestal para Bosques Templados SiPlaFor (Ávila-Márquez, 2021). Los nombres científicos se consultaron en la página webThe World flora On line List(http://www.worldfloraonline.org/).
Tabla 1 Características silvícolas, ecológicas y geográficas de los sitios de estudio.Table 1. Silvicultural, ecological and geographic characteristics of the study sites.
| Sitio | AS.N.M. | Pendiente (%) |
Orientación | Tratamiento | Manejo | Sitio | A.S.N.M. | Pendiente (%) |
Orientación | Tratamiento | Manejo |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2217 | 2839 | 26 | Norte | aclareo | MDS | 1773 | 2710 | 87 | Oeste | selección | MMOBI |
| 2040 | 2792 | 53 | Sur | aclareo | MDS | 2047 | 2785 | 32 | Este | selección | MMOBI |
| 2218 | 2858 | 30 | Norte | aclareo | MDS | 1900 | 2761 | 39 | Oeste | selección | MMOBI |
| 1908 | 2852 | 20 | Oeste | 2 aclareo | MDS | 2713 | 2255 | 64 | Noroeste | selección | MMOBI |
| 1905 | 2882 | 60 | Oeste | 2 aclareo | MDS | 1759 | 2266 | 74 | Noreste | selección | MMOBI |
| 2302 | 2828 | 40 | Sur | 2 aclareo | MDS | 1763 | 2277 | 49 | Noroeste | selección | MMOBI |
| 2305 | 2852 | 60 | Sur | 2 aclareo | MDS | 1767 | 2290 | 45 | Oeste | selección | MMOBI |
| 2216 | 2833 | 17 | Noreste | aclareo | MDS | 1747 | 2302 | 80 | Oeste | selección | MMOBI |
| 2316 | 2708 | 65 | Noreste | aclareo | MDS | 1822 | 2302 | 40 | Norte | selección | MMOBI |
| 2304 | 2830 | 30 | Sur | 2 aclareo | MDS | 1845 | 2507 | 60 | Oeste | selección | MMOBI |
| 2311 | 2753 | 15 | Sur | aclareo | MDS | 2686 | 2658 | 65 | Suroeste | selección | MMOBI |
| 2037 | 2765 | 34 | Sureste | aclareo | MDS | 2032 | 2800 | 68 | Norte | selección | MMOBI |
| 1313 | 2752 | 21 | Este | 4 aclareo | MDS | 1932 | 2683 | 65 | Noreste | selección | MMOBI |
| 2303 | 2804 | 60 | Sur | selección | MMOBI | 2319 | 2865 | 64 | Sureste | selección | MMOBI |
| 1957 | 2560 | 80 | Suroeste | selección | MMOBI | 2048 | 2784 | 29 | Oeste | selección | MMOBI |
| 1747 | 2302 | 80 | Oeste | selección | MMOBI | 1940 | 2517 | 72 | Sureste | selección | MMOBI |
| 2210 | 2893 | 10 | Este | selección | MMOBI | 2309 | 2644 | 67 | Este | selección | MMOBI |
| 1981 | 2572 | 60 | Noroeste | selección | MMOBI | 1899 | 2743 | 64 | Oeste | selección | MMOBI |
| 1952 | 2550 | 80 | Sur | selección | MMOBI | 1948 | 2815 | 62 | Suroeste | selección | MMOBI |
| 1793 | 2298 | 60 | Oeste | selección | MMOBI | 2315 | 2702 | 75 | Sur | selección | MMOBI |
| 1806 | 2272 | 35 | Sur | selección | MMOBI | 1947 | 2750 | 62 | Sur | selección | MMOBI |
| 1932 | 2683 | 64 | Noreste | selección | MMOBI | 2320 | 2808 | 60 | Sureste | selección | MMOBI |
| 1996 | 2758 | 64 | Noroeste | selección | MMOBI | 2760 | 2768 | 24 | Noroeste | selección | MMOBI |
| 1958 | 2682 | 34 | Sur | selección | MMOBI | 2317 | 2736 | 58 | Sur | selección | MMOBI |
| 1926 | 2815 | 20 | Oeste | selección | MMOBI | 1992 | 2760 | 58 | Este | selección | MMOBI |
| 1847 | 2770 | 19 | Noroeste | selección | MMOBI | 1976 | 25666 | 80 | Norte | selección | MMOBI |
Análisis de la información
Con la información de campo y para cada tratamiento por tipo de manejo y
orientación geográfica se realizó un análisis de la composición de especies
entre los métodos de manejo MMOBI y MDS, se calculó el área basal y la cobertura
en función del diámetro normal y diámetro de copa, con la fórmula del cilindro
Para estimar la diversidad en las comunidades forestales evaluadas, se aplicaron los índices de Margalef [DMg], Shannon [H´] y el número efectivo de especies con base en la exponencial del índice de Shannon (exp (H´)) llamado también índice verdadero de Shannon, conocido como (1D), los cuales estiman la diversidad alfa (Moreno, 2001, 2019; Cultid-Medina C & Escobar F., 2019).
Con la identificación de cada especie y su abundancia en ambos tipos de manejo, se calculó el Índice de Dominancia de Simpson (D). Este índice varía de cero a uno, donde 0 significa que no hay dominancia y 1 significa dominancia por especies dentro de la comunidad (Simpson, 1949). Con el índice D, se estimó el Índice de Diversidad de Simpson (IDS = 1 - D), también llamado complemento de Simpson, el cual oscila entre 0 donde solo hay una especie y 1 donde hay mucha diversidad. Se realizó una prueba “t” para determinar diferencias estadísticas entre los índices de diversidad en ambos métodos de manejo. Para los análisis de diversidad se utilizó el Excel 2021 (18.0) y el software PAST 3.22 (Hammer, 2001).
Para evaluar la diversidad entre hábitats (grado de reemplazamiento de especies) o cambio biótico a través de condiciones ambientales, se generó un modelo de ordenación Bray-Curtis, el cual es una representación gráfica de la variación de la composición vegetal (Bray & Curtis, 1957).
El análisis se realizó entre tratamientos (Selección, Selección N NO NE, Selección S SE SO, Selección EO, Aclareo, Aclareo N NO NE, Aclareo S SE SO y Aclareo EO). El modelo es un algoritmo que determina el porcentaje de similitud en una medida de distancia, proporciona valores entre 0 y 100, donde el 100 significa que los sitios tienen la misma composición y 0 significa que no comparten especies entre comunidades (Bray & Curtis, 1957). Esta gráfica se elaboró con el algoritmo UPGMA y 999 permutaciones para respaldar el valor de cada rama (Hammer, 2001). Para determinar la distribución de los datos se realizaron pruebas de normalidad Shapiro-Wilk cuando n ≤ 2000 con α = 0.05.
Resultados
Composición de especies
En cuanto a composición de especies, se registraron 24 especies para sitios con MMOBI y 11 para sitios con MDS, distribuidas en cinco familias (Tabla 2). La familia Fagaceae fue la mejor representada con un 46% (11 especies), seguida por Pinaceae con 42% (ocho especies) y 12% por las familias Cupressaceae, Betulaceae y Ericaceae. La familia Fagaceae fue representada por el género Quercus, mientras que Pinaceae tuvo nueve especies del género Pinus y una de Pseudotsuga; en la familia Cupressaceae aparecen el género Juniperus, en la Betulaceae Alnus y en Ericaceae Arbutus.
Tabla 2 Listado del arbolado del área de estudio y densidades (Árboles ha-1) por tipo de manejo. Los nombres de las especies son acordes a la (WFO’s, 2024).Table 2. List of trees in the study area and densities (Trees ha-1) by management type. Species names are according to (WFO's, 2024).
| Familia | Especie | Densidad (n ha-1) | |||
|---|---|---|---|---|---|
| MMOBI | (%) | MDS | (%) | ||
| Betulaceae | Alnus spp Kunth | 19.2 | 4.34 | 46.9 | 9.65 |
| Cupressaceae | Juniperus deppeana Steud. | 12.9 | 2.92 | 20 | 4.11 |
| Ericaceae | Arbutus xalapensis Kunth | 50 | 11.31 | 28.5 | 5.86 |
| Fagaceae | Quercus candicans Née | 0.5 | 0.11 | 0 | 0.00 |
| Fagaceae | Quercus coccolobifolia Trel. | 1.8 | 0.41 | 0 | 0.00 |
| Fagaceae | Quercus crassifolia Humb. & Bonpl. | 9.2 | 2.08 | 0 | 0.00 |
| Fagaceae | Quercus durifolia Seemen ex Loes. | 5.5 | 1.24 | 0 | 0.00 |
| Fagaceae | Quercus eduardii Trel. | 1.8 | 0.41 | 0 | 0.00 |
| Fagaceae | Quercus emoryi Porter & J.M.Coult. | 0.3 | 0.07 | 0 | 0.00 |
| Fagaceae | Quercus fulvaLiebm. | 3.9 | 0.88 | 0 | 0.00 |
| Fagaceae | Quercus rugosa Née | 35.5 | 8.03 | 21.5 | 4.42 |
| Fagaceae | Quercus sideroxyla Bonpl. | 46.6 | 10.54 | 18.5 | 3.81 |
| Fagaceae | Quercus urbanii Trel. | 11.6 | 2.62 | 0 | 0.00 |
| Fagaceae | Quercus viminea Trel. | 2.9 | 0.66 | 0 | 0.00 |
| Pinaceae | Pinus chihuahuana Martínez | 3.9 | 0.88 | 0.8 | 0.16 |
| Pinaceae | Pinus cooperi C.E. Blanco | 27.9 | 6.31 | 160 | 32.91 |
| Pinaceae | Pinus durangensis Martínez | 136.6 | 30.90 | 149.2 | 30.69 |
| Pinaceae | Pinus engelmannii Carrière | 9.7 | 2.19 | 0 | 0.00 |
| Pinaceae | Pinus herrerae Martínez | 11.3 | 2.56 | 0 | 0.00 |
| Pinaceae | Pinus leiophylla Schiede ex Schltdl. et. Cham. | 8.2 | 1.86 | 6.9 | 1.42 |
| Pinaceae | Pinus lumholtzii B. L. Rob et Fernald | 3.2 | 0.72 | 0 | 0.00 |
| Pinaceae | Pinus strobiformis Engelm. | 27.1 | 6.13 | 33.1 | 6.81 |
| Pinaceae | Pinus teocote Schiede ex Schltdl. et Cham. | 12.1 | 2.74 | 0.8 | 0.16 |
| Pinaceae | Pseudotsuga spp (Mirb.) Franco | 0.3 | 0.07 | 0 | 0.00 |
| TOTALES | 442 | 100 | 457.7 | 100 | |
Se muestra que con las cortas selectivas en el MMOBI existen muchas especies exclusivas (13 especies), principalmente del género Quercus y algunas especies de baja densidad como el Pseudotsuga spp. y otras del género Pinus. En el MDS, las 11 especies que aparecen también se pueden encontrar cuando se aplica MMOBI (Tabla 2).
En cuanto a las densidades, para el MMOBI, el P. durangensis, Arbutus spp y Q. sideroxyla resultaron ser las especies que representan el mayor número de árboles por unidad de superficie, mientras que para MDS el P. cooperi, el P. durangensis y el Alnus spp. Existen otras especies con poca abundancia de los géneros Quercus y Pinus con porcentajes menores al 3%, principalmente en el MMOBI. Se observa que el P. durangensis y P. cooperi son las especies con mayor densidad, aunque en MDS cuando se aplican aclareos se aprecia como el P. cooperi incrementa su densidad. El Q. sideroxyla juega un papel importante en la composición de especies, sin embargo, se observa que con los aclareos del MDS se reduce significativamente su densidad.
Se puede observar también que las especies de interés comercial son las que van dejando los técnicos cuando se aplica MDS, el P. cooperi (160 árboles ha-1) y el P. durangensis (149 árboles ha-1), representando éstas dos especies el 64% de la densidad total. Sin embargo, se aprecia en este método, que se están incluyendo otras especies que cohabitan como el P. leiophylla, P. teocote, Juniperus depeana, Q. sideroxyla, Q. rugosa, Arbutus y Alnus, especies que anteriormente se eliminaban por la técnica llamada chaponeo.
Selección (MMOBI) vs Aclareos (MDS)
En cuanto a la dominancia, en rodales con selección (MMOBI) el P. durangensis y Q. sideroxyla son las especies que presentan mayor área basal con 6.52 m2 ha-1 y 1.84 m2 ha-1 respectivamente.
En densidad, el P. durangensis y A. xalapensis ocuparon los primeros lugares, con 137 y 50 árboles por hectárea en selección (MMOBI), además en el 84% de los sitios con MMOBI se encontraron estas especies, seguidas por Q. sideroxyla y Q. rugosa. (Tablas 3 y 4).
Tabla 3 Parámetros ecológicos relativos para selección y aclareos para los dos métodos de manejo.Table 3. Relative ecological parameters for selection and thinning for the two management methods.
| ESPECIE | Dominancia (Área Basal) | Densidad | Frecuencia | IVI (%) | ||||
|---|---|---|---|---|---|---|---|---|
| MMOBI (%) | MDS (%) | MMOBI (%) | MDS (%) | MMOBI (%) | MDS (%) | MMOBI | MDS | |
| Pinus cooperi | 7.65 | 19.4 | 6.31 | 32.91 | 3.8 | 10.5 | 5.93 | 20.95 |
| Pinus durangensis | 36.13 | 37.5 | 30.89 | 30.70 | 13.7 | 15.8 | 26.90 | 28.00 |
| Pinus leiophylla. | 1.87 | 1.79 | 1.85 | 1.42 | 4.7 | 5.3 | 2.80 | 2.83 |
| Pinus teocote. | 2.54 | 0.44 | 2.74 | 0.16 | 2.6 | 1.3 | 2.61 | 0.64 |
| Pinus engelmannii | 1.62 | 2.20 | 1.7 | 1.84 | ||||
| Pinus lumholtzii | 0.56 | 0.71 | 1.3 | 0.85 | ||||
| Pinus ayacahuite | 3.12 | 4.51 | 6.13 | 6.80 | 10.7 | 13.2 | 6.65 | 8.16 |
| Pinus chihuahuana | 1.10 | 0.03 | 0.89 | 0.16 | 1.3 | 1.3 | 1.09 | 0.50 |
| Pinus herrerae | 2.57 | 2.56 | 2.1 | 2.42 | ||||
| Juniperus deppeana | 2.64 | 2.78 | 2.92 | 4.11 | 3.4 | 11.8 | 2.99 | 6.25 |
| Pseudotsuga spp | 0.11 | 0.06 | 0.4 | 0.20 | ||||
| Quercus sideroxyla | 10.21 | 10.0 | 10.54 | 3.80 | 11.1 | 10.5 | 10.62 | 8.13 |
| Quercus durifolia | 0.42 | 1.25 | 0.4 | 0.70 | ||||
| Quercus coccolobifolia | 0.58 | 0.42 | 1.3 | 0.76 | ||||
| Quercus eduardii | 0.80 | 0.42 | 1.3 | 0.83 | ||||
| Quercus urbanii | 3.78 | 2.62 | 3.8 | 3.42 | ||||
| Quercus crassifolia | 2.53 | 2.08 | 3.0 | 2.53 | ||||
| Quercus viminea | 0.30 | 0.65 | 1.7 | 0.89 | ||||
| Quercus emoryi | 0.06 | 0.06 | 0.4 | 0.18 | ||||
| Quercus fulva | 1.47 | 0.89 | 0.9 | 1.07 | ||||
| Quercus candicans | 0.19 | 0.12 | 0.9 | 0.39 | ||||
| Quercus rugosa | 9.54 | 12.3 | 8.04 | 4.43 | 9.8 | 7.9 | 9.14 | 8.19 |
| Alnus spp | 2.40 | 6.53 | 4.35 | 9.65 | 6.0 | 11.8 | 4.24 | 9.34 |
| Arbutus xalapensis | 7.80 | 4.69 | 11.31 | 5.85 | 13.7 | 10.5 | 10.93 | 7.02 |
| TOTALES | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
La dominancia mayor en aclareos la representa el P. cooperi (7.92 m2 ha-1) y el P. durangesnsis con 4.10 m2 ha-1. Se observa también que con la aplicación del MDS a través de los sitios de aclareo, muchas especies del género Quercus ya no se encuentran, posiblemente por el chaponeo o a factores abióticos o bien a otras actividades complementarias que se realizan. En el 92.3% de los sitios con aclareo se encontró el P. durangesnis y en el 76.9 % el P. strobiformis, en 69.2% el Alnus spp. y el Juniperus depeana, el P. cooperi en un 61.5%, luego Q. rugosa en un 46.2% de los sitios.
El P. durangensis y el P. cooperi presentaron los mayores porcentajes de IVI en sitios con MDS y con el tratamiento de aclareos (Tabla 4), las especies que aparecieron en aclareos con menores IVI entre 9% y 1% fueron: Alnus spp, Q. rugosa, P. strobiformis, Q. sideroxyla, A. xalapensis, Juniperus deppeana, P. leiophylla, P. teocote y P. chihuahuana, estas dos últimas especies se encontraron en un 7% de los sitios.
Tabla 4 Parámetros estructurales por tratamiento para las cinco especies con mayor IVI, ordenadas de manera descendente.Table 4. Structural parameters by treatment for the five species with the highest IVI, ordered in descending order.
| ESPECIE | Dominancia (Área Basal) | Abundancia (Densidad) | Frecuencia | IVI (%) | |||
|---|---|---|---|---|---|---|---|
| Absoluta (m2 ha -1) | Relativa (%) | Absoluta (N ha-1) | Relativa (%) | Absoluta (%) | Relativa (%) | ||
| Selección | |||||||
| Pinus durangensis | 6.52 | 36.13 | 136.6 | 30.89 | 84.2 | 13.7 | 26.90 |
| Arbutus xalapensis | 1.41 | 7.80 | 50.0 | 11.31 | 84.2 | 13.7 | 10.93 |
| Quercus sideroxyla | 1.84 | 10.21 | 46.6 | 10.54 | 68.4 | 11.1 | 10.62 |
| Quercus rugosa | 1.72 | 9.54 | 35.5 | 8.04 | 60.5 | 9.8 | 9.14 |
| Pinus strobiformis | 0.56 | 3.12 | 27.1 | 6.13 | 65.8 | 10.7 | 6.65 |
| Aclareos | |||||||
| Pinus durangensis | 7.92 | 37.50 | 149.2 | 30.70 | 92.3 | 15.8 | 28.00 |
| Pinus cooperi | 4.10 | 19.40 | 160.0 | 32.91 | 61.5 | 10.5 | 20.95 |
| Alnus spp. | 1.38 | 6.53 | 46.9 | 9.65 | 69.2 | 11.8 | 9.34 |
| Quercus rugosa | 2.59 | 12.26 | 21.5 | 4.43 | 46.2 | 7.9 | 8.19 |
| Pinus strobiformis | 0.95 | 4.51 | 33.1 | 6.80 | 76.9 | 13.2 | 8.16 |
| Selección N NE NO | |||||||
| Pinus durangensis | 8.19 | 38.63 | 124.5 | 30.79 | 72.7 | 10.7 | 26.69 |
| Quercus sideroxyla | 2.24 | 10.55 | 64.5 | 15.96 | 54.5 | 8.0 | 11.50 |
| Arbutus xalapensis | 1.27 | 5.97 | 34.5 | 8.54 | 72.7 | 10.7 | 8.39 |
| Pinus strobiformis | 0.77 | 3.64 | 22.7 | 5.62 | 81.8 | 12.0 | 7.09 |
| Quercus rugosa | 1.49 | 7.02 | 19.1 | 4.72 | 36.4 | 5.3 | 5.69 |
| Aclareo N NE NO | |||||||
| Pinus cooperi | 8.49 | 50.71 | 330.0 | 63.46 | 75.0 | 13.6 | 42.60 |
| Pinus durangensis | 3.48 | 20.76 | 75.0 | 14.42 | 75.0 | 13.6 | 16.27 |
| Pinus strobiformis | 1.49 | 8.87 | 47.5 | 9.13 | 100.0 | 18.2 | 12.06 |
| Quercus rugosa | 2.50 | 14.93 | 17.5 | 3.37 | 25.0 | 4.5 | 7.61 |
| Juniperus deppeana | 0.39 | 2.36 | 15.0 | 2.88 | 75.0 | 13.6 | 6.29 |
| Selección S SE SO | |||||||
| Pinus durangensis | 7.09 | 40.63 | 166.2 | 35.01 | 92.3 | 14.0 | 29.86 |
| Quercus sideroxyla | 2.10 | 12.06 | 66.2 | 13.94 | 84.6 | 12.8 | 12.93 |
| Arbutus xalapensis | 1.76 | 10.11 | 53.8 | 11.35 | 100.0 | 15.1 | 12.19 |
| Quercus rugosa | 2.06 | 11.83 | 41.5 | 8.75 | 92.3 | 14.0 | 11.51 |
| Aclareo S SE SO | |||||||
| Pinus durangensis | 11.39 | 49.19 | 200.0 | 42.70 | 100.0 | 16.2 | 36.04 |
| Pinus cooperi | 2.11 | 9.11 | 75.0 | 16.01 | 50.0 | 8.1 | 11.08 |
| Arbutus xalapensis | 1.78 | 7.69 | 50.0 | 10.68 | 83.3 | 13.5 | 10.63 |
| Quercus rugosa | 3.15 | 13.61 | 30.0 | 6.41 | 66.7 | 10.8 | 10.28 |
| Quercus sideroxyla | 2.71 | 11.69 | 28.3 | 6.05 | 66.7 | 10.8 | 9.52 |
| Selección E O | |||||||
| Pinus durangensis | 4.75 | 25.75 | 132.5 | 25.85 | 87.5 | 14.89 | 22.16 |
| Arbutus xalapensis | 1.67 | 9.05 | 78.8 | 15.37 | 87.5 | 14.89 | 13.10 |
| Quercus rugosa | 2.34 | 12.69 | 70 | 13.66 | 50 | 8.51 | 11.62 |
| Pinus cooperi | 1.96 | 10.62 | 40 | 7.80 | 12.5 | 2.13 | 6.85 |
| Quercus sideroxyla | 1.21 | 6.56 | 8.8 | 1.71 | 62.5 | 10.64 | 6.30 |
| Aclareo E O | |||||||
| Pinus durangensis | 6.16 | 28.57 | 126.7 | 28.57 | 100.00 | 17.65 | 24.93 |
| Alnus spp | 5.21 | 24.15 | 126.7 | 28.57 | 100.00 | 17.65 | 23.46 |
| Pinus cooperi | 2.16 | 10.01 | 100 | 22.56 | 66.67 | 11.76 | 14.78 |
| Quercus sideroxyla | 3.63 | 16.84 | 6.7 | 1.50 | 66.67 | 11.76 | 10.04 |
| Pinus strobiformis | 0.68 | 3.15 | 33.3 | 7.52 | 66.67 | 11.76 | 7.48 |
En la Tabla 4 se presentan los parámetros de dominancia, densidad, frecuencia y Valor de Importancia para las primeras 5 especies con mayor IVI por orientación geográfica y por método de manejo.
Selección N NE NO vs Aclareos N NE NO
En el tratamiento Selección N NE NO (MMOBI) se encontraron 21 especies y 10 en aclareos orientación N NE NO (MDS). Se observó que varias especies del género Quercus como Q. viminea, Q. urbanii, Q. candicans, Q. eduardii, Q. coccolobifolia y Q. crassifolia y P. lumholtzi y el P. teocote que se encuentran en Selección N NE y NO ya no aparecen en el Aclareo N NE y NO. El Pseudotsuga spp. se ve afectado quizás por la falta de sombra y humedad de la orientación norte, ya que no aparece cuando se aplican aclareos.
La densidad de A. xalapensis disminuyen de 34 árboles ha-1 en Selección N NE NO a 10 árboles por hectárea en Aclareo N NE NO. El P. cooperi y el P. durangensis se encontraron en el 75% de los sitios en Aclareo N NE NO, sin embargo, resulta interesante que el P. strobiformis aparece en el 100% de los sitios con Aclareo N NE NO pero con área basal pequeña (1.49 m2 ha-1). Los valores mayores de IVI en Selección N NE NO los presenta el P. durangensis con 26.69%, Q. sideroxyla con 11.50%, A. xalapensis con 8.39% y P. strobiformis con 7.09%. Es importante que los técnicos pongan especial cuidado cuando apliquen cortas de selectivas con P. engelmannii, P. lumholtzii y Pseudotsuga spp, ya que observaron valores de IVI muy pequeños debido a sus bajas abundancias. En Aclareo N Ne NO esta dominando el P. cooperi con 8.49% y con más del 50% de las áreas basales (8.5m2 ha-1), con una densidad de 330 árboles ha-1 y con un IVI de 42.69%. El P. durangensis ocupó el segundo lugar con un IVI de 16.27 y el P. strobiformis con 12.02%. En Aclareo N Ne NO las especies con índices de valor de importancia menores fueron: P. strobiformis, Q. rugosa, Juniperus deppeana, Q. sideroxyla, Alnus spp., A. xalapensis, P. leiophylla, y P. chihuahuana.
Selección S SE SO vs Aclareo S SE SO
En el tratamiento de Selección S SE SO se encontraron 15 especies y con Aclareos S SE SO nueve especies, proliferan las hojosas, pero cuando se aplican los tratamientos también cambian las especies. En selección S SE SO el P. durangensis ocupa el primer lugar en el IVI con 29.86%, se presenta en el 92.3% de los sitios analizados, tiene densidades de 166 árboles ha-1, con 7 m2 ha-1, ocupando el 40% de área basal, le sigue el Q. sideroxyla apareciendo en el 84.6% de los sitios y con un IVI de 12.93%, luego Arbutus xalapensis con 53.8 árboles ha-1y un IVI de 12.19% pero se encuentra en todos los sitios muestreados con área basal de 1.76 m2 ha-1. El Q. rugosa también es importante en los sitios de Selección Sur con 41 árboles ha-1 y con un IVI de 11.51% encontrándose en el 92% de los sitios de muestreo con Selección S SE SO.
Resulta interesante saber que en este tratamiento (Selección S SE SO) aparecen muchas especies de hojosas y algunas coníferas de bajas abundancias; en orden descendente por su valor del IVI del quinto lugar en adelante son: P. strobiformis, P. cooperi, P. teocote., J. deppeana, Q. crassifolia, Alnus spp, P. leiophylla, P. engelmannii, Q. fulva, Q. candicans y Q. urbanii. El Juniperus deppeana, Alnus spp. y P. leiophylla.
En el Aclareo S SE SO es dominado por el P. durangensis con 200 árboles ha-1, apareciendo en el 100% de los sitios y con el máximo IVI de 26.04%, además el 49.19% de la áreas basales es ocupada por ésta especie, sigue el P. cooperi con 76 árboles ha-1, estando presente en el 50% de los sitios y un IVI de 11.08%, las otras especies tienen un IVI que va de 10 a 3% y con densidades promedio de 50 a 21 árboles ha-1, esas especies son el Arbutus xalapensis apareciendo en el 83% de los sitios, el Quercus rugosa con 13.61% del área basal con 3.15 m2 ha, Q. sideroxyla, Juniperus deppeana. Alnus spp., P. strobiformis y P. leiophylla apareciendo en el 60% de los sitios y con IVI de 9.5, 6.68, 6.3, 6,1 y 3.35% respectivamente.
Selección EO vs Aclareo EO
Para el tratamiento Selección EO y Aclareo EO, el IVI más alto fue para el P. durangesnis, con 132.5 árboles ha -1 en Selección EO y 126 en Aclareo EO, sin embargo, para Selección EO siguen el Arbutus xalapensis y el Q. rugosa, mientras que para el Aclareo EO el Alnus spp. y el P. cooperi. El P, durangensis se presentó en 87 de los sitios, el Arbutus 78% en Selección EO y el P. durangensis y el Alnus spp. en el 100% de los sitios con Aclareo EO. Para Selección EO se presentaron 18 especies y nueve para el Aclareo EO.
Diversidad Alfa
Con respecto a los índices de riqueza y diversidad, para el tratamiento Selección, Selección N NE NO, Selección S SE SO y Selección EO; el índice de riqueza de Margalef (Dmg) mostró promedios de 2.85 ± 0.48, para Shannon (H+) 2.32 ± 0.15 y para la diversidad de Simpson (1-D) 0.85±0.02. Estos valores del MMOBI, fueron superiores que para MDS con Aclareo, Aclareo N NE NO, Aclareo S Se SO y Aclareo EO, donde, el índice de Margalef fue en promedio de 1.57±0.12, el de Shannon de 1.6±0.24, y para la diversidad de Simpson un promedio de 0.72±0.11. Con el tratamiento de selección del MMOBI se observó que la riqueza es mayor, encontrando 23 especies y tan solo 11 con el MDS, se aprecia la diferencia de riqueza y diversidad entre tratamientos, observando una diversidad media para los tratamientos de Selección (MMOBI) y baja para Aclareos (MDS). Con los valores anteriores, se muestra poca dispersión entre los índices sobre todo para los tratamientos de selección, donde presenta una desviación estándar muy baja, como es el caso del índice de Simpson. Los índices para los aclareos muestran una diversidad baja y poca variación entre ellos (Tabla 5).
Tabla 5 Valores de índices de riqueza y diversidad por tipo de manejo MMOBI (selección) y MDS (Aclareos) en diferentes orientaciones geográficas.Table 5. Richness and diversity index values by management type MMOBI (selection) and MDS (thinning) in different geographic orientations.
| Tratamiento | Manejo | Especies (S) |
Margalef (Dmg) |
Diversidad de Shannon (H´) |
Número de especies efectivas (1D) |
Dominancia de Simpson (D) |
Diversidad de Simpson (1-D) |
|---|---|---|---|---|---|---|---|
| Selección | MMOBI | 24 | 3.09 | 2.43 | 11.36 | 0.14 | 0.86 |
| Selección N NE NO | MMOBI | 21 | 3.28 | 2.37 | 10.65 | 0.15 | 0.85 |
| Selección S SE SO | MMOBI | 15 | 2.18 | 2.1 | 8.2 | 0.17 | 0.82 |
| Selección E O | MMOBI | 18 | 2.83 | 2.38 | 10.88 | 0.12 | 0.87 |
| Aclareo | MDS | 11 | 1.55 | 1.77 | 5.9 | 0.22 | 0.78 |
| Aclareo N NE NO | MDS | 10 | 1.68 | 1.28 | 3.6 | 0.44 | 0.56 |
| Aclareo S SE SO | MDS | 9 | 1.42 | 1.77 | 5.9 | 0.24 | 0.76 |
| Aclareo E O | MDS | 9 | 1.64 | 1.71 | 5.52 | 0.22 | 0.78 |
Existen diferencias entre tratamientos de selección y aclareos, mostrando una mayor diversidad el tratamiento de selección en exposiciones N NE NO que en S SE SO. Se observa también como el Aclareo N NE NO presentó la menor diversidad (Figura 2).
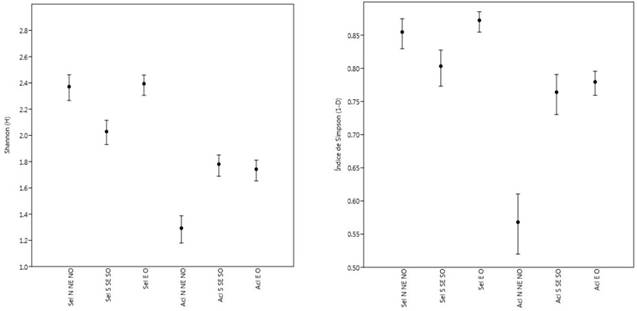
Figure 2 Shannon-Weiner and Simpson diversity indices by treatment for the two study methods (MMOBI Selection and MDS Thinning). Values plotted represent the mean ± standard error.
Se muestra también en la figura anterior que en el índice efectivo de especies (1D) se observan valores altos de las especies más abundantes para las diferentes orientaciones geográficas en el MMOBI, en cambio para el MDS valores muy bajos, lo que se puede demostrar que en este tipo de manejo y con tratamientos con aclareos la diversidad se reduce y pocas especies ocupan las mayores abundancias (P. cooperi, P. durangensis y Arbutus xalapensis ó Q. sideroxyla). El índice de dominancia de Simpson (D) representa poca probabilidad de que dos individuos seleccionados al azar sean de la misma especie en tratamientos de selección, habiendo mayor probabilidad en aclareos y sobre todo mayor en Aclareos N NE NO. El complemento de Simpson muestra que los rodales manejados con selección en las diferentes orientaciones presentan mayor diversidad como lo es el de Selección E O (0.86), Selección (0.85) y Selección N NE NO (0.82). El Aclareo N NE NO fue el que mostró menor diversidad de acuerdo al complemento de Simpson de 0.56.
Similitud entre comunidades
Los resultados obtenidos del análisis de similitud mediante la técnica de Bray-Curtis, muestran un 45% de similitud entre ambos métodos de manejo forestal. Se observan cuatro grupos de comunidades vegetales bien definidos, considerando un valor de referencia de 0.675, el primero conformado por Aclareo S SE SO, Selección S SE SO y Selección N NE NO, estas comunidades presentaron la mayor similitud de especies. El tratamiento Selección EO forma otro grupo. el tercer Aclareo EO y el cuarto grupo el Aclareo N NE NO que representó la comunidad vegetal con menor similitud de especies ocurriendo por separado del resto de las comunidades evaluadas (Figura 3).
Las especies generalistas que se registraron en todos los tratamientos analizados fueron:P. cooperi, P. durangensis, P. leiophylla, P. strobiformis, J. deppeana, Q. sideroxyla, Q. rugosa, Alnus spp. yA. xalapensis, mientras que las especies especialistas se presentaron en el MMOBI y fueron: P. engelmannii, P. lumholtzii, P. herrerae, Pseudotsuga spp, Q. durifolia, Q. coccolobifolia, Q. eduardii, Q. urbanii, Q. crassifolia, Q. viminea, Q. emoryi, Q. fulva, Q. candicans.
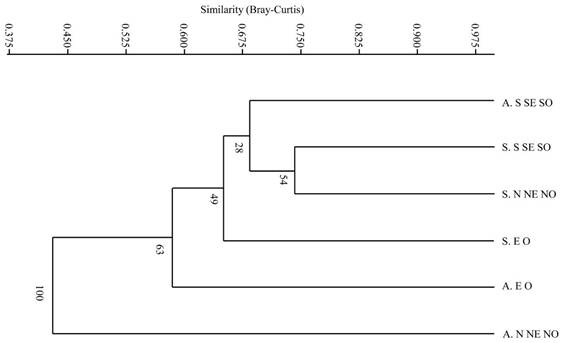
Figure 3 Dendrogram of similarity based on Bray Curtis analysis between treatments with different orientations. A: thinning and S is selection; N, S, E, W, W, NE, NW, SE, SW refer to the exposures where the studied plant communities were found. The numbers near each branch are the percentage of permutations supporting each split node in the final clustering dendrogram.
La correlación cofenética (Cophen. corr. 0.901) obtenida mostró una buena medida del grado de ajuste para la clasificación de nuestro conjunto de datos (Saraçli et al., 2013).
Discusión
Composición de especies
En el manejo de recursos forestales, la diversidad y la estructura son variables que aportan bases para la toma de decisiones en la silvicultura, manejo y aprovechamiento, así lo mencionan (García & Sánchez, 2005); López-Hernández et al. (2017); Flores-Morales et al. (2022). Estos últimos autores, reportan 24 especies y siete familias en su estudio “Diversidad y estructura arbórea de un bosque templado bajo manejo en el municipio de Pueblo Nuevo, Durango, México”, resultados semejantes en el tratamiento de selección del presente trabajo. ya que el de referencia también está manejado con MMOBI con el tratamiento de selección.
Las familias representadas en los métodos de manejo analizados coinciden con las reportadas por García Arévalo & González Elizondo (1998) Graciano-Ávila et al. (2017) para las cadenas montañosas de Durango, dominadas por Pinaceae y Fagaceae. Las especies con mayor abundancia pertenecen al género Pinus, concordando con López-Hernández et al. (2017) que mencionan a este género como el de mayor abundancia en los bosques templados. El número de especies resultó mayor que el reportado por Graciano-Ávila et al. (2020) de 16 especies en 4 familias, en el estudio de cambios estructurales de la vegetación arbórea en un bosque templado de Durango, México con MMOBI.
Los más de 40 estudios revisados por Monárrez-González et al. (2018) con respecto al manejo de ecosistemas y la diversidad, donde se destaca que los tratamientos silvícolas modifican la estructura y diversidad, coinciden con los resultados de este trabajo, ya que se demuestra que al aplicar MDS se modifica la riqueza de especies, aunque faltaría comparar con rodales donde no se ha realizado ningún tipo de manejo. Algunos autores encontraron una relación entre el manejo forestal y la diversidad arbórea (Solís-Moreno et al., 2006; Alanís-Rodríguez et al., 2010; Leyva-López et al., 2010 y Graciano-Ávila et al., 2020), donde se modifica la riqueza, pero se incrementa después del manejo por reclutamiento de rodales adyacentes o por los tipos de propagación vegetativa de algunas especies (Graciano, 2001 y Hernández-Salas et al., 2013).
Parámetros estructurales
La diferencia estructural encontrada entre sitios con MMOBI y MDS es consistente con la estructura documentada por diversos autores comoGraciano (2001),Solís-Moreno et al. (2006), Hernández-Salas et al. (2013), Graciano-Ávila et al. (2017, 2020), Flores-Morales et al. (2022) y Silva-González et al. (2021) en bosques templados. Estas conclusiones semejantes se deben a que el área de estudio está localizada en la llamada Región de El Salto, donde a nivel paisaje comparten factores abióticos como algunas características geofísicas, por lo que se prescriben los mismos tratamientos silvícolas, bajo los dos tipos de manejo analizados.
Los resultados obtenidos en densidad coinciden con los citados por Delgado Zamora et al. (2016) y Graciano-Ávila et al. (2017) de 565 y 575 árboles ha-1, en bosques templados de Durango. Silva-García et al. (2022), en su trabajo sobre la influencia de la altitud y orientación geográfica en la estructura y composición de un bosque templado reportan densidades y áreas basales que coinciden con los resultados de este trabajo en un intervalo altitudinal de 2400 a 2500 m s.n.m. Flores-Morales et al. (2022), reporta a P. cooperi con 158 árboles ha-1, Q. Sideroxyla con 130 árboles ha-1 y al P. durangensis con 75 árboles ha-1 como las especies de mayor densidad en un trabajo similar en bosques de Durango. Por el contrario, (López-Hernández et al. (2017)registraron valores más bajos, de 389 árboles ha-1y 11 especies en bosques del estado de Puebla, lo cual puede atribuirse a que los bosques evaluados presentan actividades más intensivas de aprovechamiento, en comparación con las del presente estudio.
Silva-García et al. (2022) reportan que, al aumentar la altitud, el área basal aumenta, asimismo, también mencionan que la orientación y la pendiente pueden influir en la productividad de diversas áreas. Los resultados de este estudio muestran que el área basal total es mayor en aclareos con 21.12 m2 ha-1. esto se debe a que, desde el punto de vista fisiológico, al abrir la masa, se incrementa la cantidad de luz solar que detona una mayor tasa fotosintética y por lo tanto mayor desarrollo, sin embargo, no hay diferencias significativas por orientación geográfica. Estos autores concluyen que las características topográficas influyen en la estructura y riqueza de especies, que la mayor diversidad florística se encuentra en altitudes bajas y exposiciones sur. Los resultados son similares a los estimados por (Graciano-Ávila et al. (2017), quienes calcularon un área basal de 23.54 m2 ha-1 en bosques templados de Durango con MMOBI. El área basal estimada es mayor a la encontrada por Caballero-Cruz et al. (2022) quienes obtuvieron valores de (22.21, 19.30 y 32.28 m2ha-1) en el análisis de la estructura y diversidad arbórea de bosques templados en la ladera oriental del volcán Iztaccíhuatl, México. También es superior al reportado por Dávila-Lara et al. (2019), para un bosque templado en San Luis Potosí y similar a lo registrado para bosques templados de Nuevo León por Ramos-Hernández et al. (2024)y en Chihuahua por Hernández-Salas et al. (2013), es decir que los bosques de Durango presentan mayores dominancias (área basal total por especies).
Los resultados del IVI, muestran que en selección con MMOBI P. durangensis presenta la mayor importancia relativa (26%), sin embargo, en sitios con aclareos del MDS, se observó que P. cooperi llega a tener hasta más del 40% de IVI. Graciano-Ávila et al. (2017) reportan que P. Cooperi, P. durangensis y Q. sideroxyla ocupan el 26, 23 y 15% respectivamente en un bosque de Durango con MMOBI. En 2020 en el estudio “Cambios estructurales de la vegetación en un bosque templado de Durango, México”, estos mismos autores coinciden con las especies de mayor porcentaje de IVI (P. durangensis 46%, Q. sideroxyla 16% y P. cooperi con 11%). Lo anterior obedece a que los principios del MDS es uniformizar la estructura con las especies de mejor calidad de la madera y de mayor productividad, por lo que P. cooperi y P. durangensis son las especies ideales para dichos fines. Cuando se aplica un tratamiento, se dejan espacios donde ecológicamente serán ocupados por individuos que se reclutarán para aprovechar los recursos y generalmente lo van a hacer las especies con mejor capacidad de reproducción, de competencia y de dispersión. Flores-Morales et al. (2022) también coinciden con que P. cooperi con 20%, Q. sideroxyla con 16% y P. durangensis con 11% son las especies que presentan mayores valores de IVI en los bosques templado de Durango. Domínguez-Gómez et al. (2018) realizaron un estudio en la zona norte de la región de El Salto, donde las condiciones de calidad de sitio son pobres y ahí encuentran que Q. sideroxyla con 34%, P. teocote y Arbutus jalapensis ocupan los primeros lugares en el IVI. En este estudio la orientación geográfica influyó en la riqueza de especies, así como en su productividad, ya que las orientaciones sur, sureste y suroeste ofrecen mayor cantidad de luz, favoreciendo la fotosíntesis y la oportunidad de ocupar espacios más rápidamente. La consociación deP. cooperiy la asociación deP. durangensiscon otras especies dePinusyQuercus,se cuentan entre las más representativas de la vegetación de la Sierra Madre Occidental en Durango (Delgado Zamora et al., 2016).
Diversidad
Con respecto a la diversidad alfa, el índice de diversidad de Shannon de 2.3 para tratamientos de selección fueron semejantes a los estimados en bosque templados por Solís-Moreno et al. (2006) de 2.05, y a los de Domínguez-Gómez et al. (2018) quienes reportaron 2.53, fue inferior al de López-Hernández et al. (2017) de 3.94, para bosques templado de Puebla, también a los reportados por Graciano-Ávila et al. (2017) en bosques de la misma región que el área de estudio que fueron de 5.75. Estos resultados muestran que el MMOBI permite que, con el tratamiento de selección, se mantenga la biodiversidad, ya que se observó que la composición florística no se ve afectada, faltaría estudiar dicha diversidad a través de rasgos funcionales para comprobar que se conservan los procesos y servicios ecosistémicos. El índice de Shannon para aclareos de 1.6, fue muy similar a los 1.91 y 1.51 encontrados por Dávila-Lara et al. (2019), en bosques templados de San Luís Potosí. Los resultados también muestran mayor diversidad de la Sierra Madre Occidental que la Oriental, ya que Ramos-Hernández et al. (2024) muestran índices de Shannon-Wiener de 1.6, 1.24 y 1.15 para bosques de pino, encino pino y encino respectivamente en la Sierra Madre Oriental. El número efectivo de especies (1D) fue más alto para selección con MMOBI (11.36) que en aclareos con MDS (5.9), estos valores concuerdan con los trabajos reportados por Solís-Moreno et al. (2006), Graciano-Ávila et al. (2017) y Domínguez-Gómez et al. (2018) donde los valores más altos son para bosques irregulares y más antiguos, tal como el patrón general para matorrales encontrado en estudios donde las comunidades de plantas más antiguas tienen mayor riqueza y diversidad de especies (Alanís-Rodríguez et al., 2018, 2023). El complemento de Simpson se utilizó para describir cual tratamiento es el más diverso, encontrando valores que van desde 0.56 a 0.87, más altos que los que reporta Solís-Moreno et al. (2006) y parecidos a los de López-(Hernández-Salas et al., 2013 y Graciano-Ávila et al., 2017).
Los resultados de este trabajo coinciden con los de Flores-González et al. (2021), donde demuestran que no existen diferencias significativas en la dominancia y abundancia, sin embargo, a diferencia de los resultados de este autor donde no encontró diferencias entre la riqueza con aclareos, en este estudio si presentaron diferencias, reduciendo el número de especies.
Similitud Florística
A través del algoritmo de Bray Curtis se muestra que algunos tratamientos comparten varias especies entre orientaciones, quizás por la intensidad de luz y la cantidad de sombra y humedad, tal como lo consideran Martínez-Pasturet al.(2007) donde mencionan que la prescripción de métodos silvícolas se basa para mantener un equilibrio entre la apertura del dosel para la llegada de la lluvia al suelo y favorecer el ingreso de luz, así como para mantener una cobertura arbórea para la protección y la generación de semillas. Siles et al. (2017) mencionan que la orientación, la inclinación y la altitud, tienen gran influencia en la composición florística y la riqueza de especies. Para orientaciones sur se presentó mayor riqueza en los tratamientos de Selección, donde aparecieron más especies de hoja ancha que en comparación con las orientaciones Norte donde P. cooperi y P. durangensis son los dominantes en Aclareos, esto se debe a que las condiciones geográficas y factores abióticos del área de estudio son favorables para el desarrollo de estas especies, principalmente en orientaciones norte. La asociación es mayor a la obtenida por Ramos-Hernández et al. (2024) que fue de 21% en bosques de la Sierra Madre Oriental. Los resultados también prueban que la similitud entre comunidades está ligada a la altitud y orientación geográfica, como lo dice Chust et al. (2006) y Silva-García et al. (2022) que va de 46 a 65%, donde estos factores pueden determinar las especies que deberán de adaptarse a diferentes comunidades. La similitud depende en gran medida de la adaptación de los taxones (Hernández-Salas et al. 2013) y a las condiciones naturales y de disturbio del hábitat (Delgado-Zamora et al., 2016). Luna-Bautista et al. (2015) mencionan que la similitud de especies aumenta con el tiempo de aplicación de tratamientos silvícolas, debido a los factores bióticos y abióticos asociados al ecosistema. Domínguez-Gómez et al. (2018) reportan baja similitud en el dendrograma de ordenación Bray-Curtis al estudiar la estructura y composición de la vegetación en cuatro sitios de la Sierra Madre Occidental, y esto se atribuye a las condiciones del sitio y a los métodos de manejo.
MMOBI y MDS
Solís-Moreno et al. (2006) en su estudio sobre el efecto de dos tratamientos silvícolas en la estructura de ecosistemas forestales en Durango, México, concluyen también como los resultados de este trabajo que en los tratamientos de selección (MMOBI) existe mayor mezcla y diversidad de especies que en aquellos sitios donde se aplican cortas por aclareos (MDS). Silva-González et al. (2021), realizaron un estudio del efecto de tratamientos silvícolas en la diversidad y estructura forestal en bosques templados bajo manejo en Durango, en el cual concluyen que el aprovechamiento forestal no modifica la diversidad y estructura de especies del estrato arbóreo, esto contrasta con los resultados obtenidos en el presente trabajo donde no se observan diferencias en ciertos parámetros estructurales como en la dominancia y abundancia, pero si en la diversidad de especies.
Hernández-Salas et al. (2013), realizaron una investigación sobre los cambios en la estructura y composición del bosque bajo dos tratamientos silvícolas en Oaxaca, los resultados demostraron que en las zonas de aprovechamiento forestal, después de la aplicación de los tratamientos, se formó un sólo tipo de bosque del género Pinus, por lo que las tendencias de los resultados del presente trabajo demuestran que si se sigue utilizando el MDS a través de aclareos, se debe considerar la composición de especies, dado que pueden desaparecer especies diferentes a las pináceas que comparten hábitat de manera natural.
Bhat et al. (2020) concluyen que la orientación geográfica influye en la riqueza, abundancia y dominancia de las especies, así como en la distribución en las cadenas montañosas. Es necesario seguir evaluando dichos cambios a nivel rodal, haciendo énfasis en las principales orientaciones, ya que las comunidades vegetales difieren de acuerdo a su orientación geográfica (Santiago et al., 2019).
La aplicación del MDS en superficies mayores, requiere de evaluaciones para entender los cambios en la estructura a largo plazo. Los resultados también muestran que se requiere el conocimiento de factores ecológicos que propicien una buena diversidad de especies no compartidas entre MMOBI y MDS para el apoyo de acciones efectivas de regeneración natural de las especies sensibles a la aplicación del MDS, tal como lo menciona Ramírez-Marcial et al. (2001) y Pérez-López et al. (2020) en su estudio sobre los efectos del “Método de Desarrollo Silvícola” sobre la diversidad arbórea en bosques húmedos de montaña del norte de Chiapas, México, donde registran principalmente efectos de bajo impacto sobre los principales parámetros de la estructura y diversidad.
Conclusiones
El método de manejo es determinante para la composición florística, la riqueza y la estructura silvícola dasométrica, ya que el MMOBI a través del tratamiento de selección, se puede considerar como una técnica conservadora que promueve la permanencia de la diversidad y mantiene una estructura heterogénea y el MDS a través de los aclareos reduce la riqueza y modifica los parámetros estructurales de dominancia, densidad, frecuencia y valor de importancia, manteniendo una estructura homogénea.
Los resultados indican que dentro de las áreas evaluadas en las diferentes orientaciones geográficas y por método de manejo se presentaron alunas diferencias en cuanto a la diversidad, lo que hace suponer que la orientación influye en la riqueza y diversidad, densidad e IVI pero no así en los parámetros de área basal y cobertura. Es importante tomar medidas de conservación de las especies con valores de IVI bajos al momento de hacer el manejo para que se conserve la riqueza de los bosques. Estas especies son: Q. candicans, Pseudotsuga spp en Selección y Selección N NE NO al igual que el P. lumholtzii, Q. fulva, Q. candicans y Q. urbanii en Selección S SE SO.
El impacto de los dos tipos de manejo MMOBI y MDS, así como de sus tratamientos sobre la conservación de la diversidad vegetal y la estructura está relacionado con la intensidad y la calidad productiva del bosque, ya que de esto depende su resiliencia para mantener dicha estructura.

