












 (pdf)
(pdf)




 SciELO
SciELO  SciELO
SciELO 
 Permalink
PermalinkIntroducción
En Jordania, en 2015, se reportaron síntomas de rugosidad marrón en frutos de jitomate (Solanum lycopersicum), con incidencia de 100 % a nivel invernadero. Estudios etiológicos determinaron un nuevo Tobamovirus denominado Tomato brown rugose fruit virus (ToBRFV) (Salem et al., 2016). Se ha postulado que ToBRFV pudo haber surgido de la recombinación de 314 nucleótidos de la región 534-848 del gen de la replicasa (Maayan et al., 2018). El Tomato mottle mosaic virus (ToMMV) y Tobacco mosaic virus (TMV) se han propuesto como ancestros directos, siendo este último el principal (Salem et al., 2016). El genoma del ToBRFV, provenientes de diferentes países, en general exhiben baja variabilidad, lo que sugiere un proceso evolutivo a partir de un descendiente común (Salem et al., 2016). Adicionalmente, Maayan y colaboradores (2018) mediante estudios filogenéticos en Tobamovirus concluyeron que ToBRFV ha tenido un proceso de evolución divergente con adaptación a diferentes hospederos, pero con baja tasa de mutación en un lapso de 3-4 años. En general, los virus tienen potencial para evolucionar y adaptarse rápidamente bajo la presión de selección natural debido a altas tasas poblacionales resultado de la efectiva replicación intraespecie, ocurrencia de cuasiespecies y ausencia de mecanismos de corrección de genomas (virus de ARN) lo que facilitan la variación genética, y tiempos cortos de generación (Hanssen et al., 2010).
En México, el ToBRFV fue reportado por primera vez en 2018 (Cambrón-Crisantos et al., 2018). Este Tobamovirus se transmite principalmente por semilla. Actualmente se ha reportado en 20 estados de la republica incluyendo las principales entidades de producción de jitomate (Solanum lycopersicum) y chile (Capsicum annuum) (Camacho-Beltran et al., 2019; Cambrón-Crisantos et al., 2018). Los frutos en jitomate se asocian con una coloración amarilla, manchas verdes y deformación, estriado verde y manchas irregulares color marrón. En follaje se observa mosaico, moteado y amarillamiento. Los primeros reportes de virus incluyeron pérdida total de producción (Cambrón-Crisantos et al., 2018). Por lo anterior, el objetivo de esta investigación fue analizar la variabilidad de dos aislados mexicanos de ToBRFV posterior a un proceso de inoculación y multiplicación en diferentes variedades comerciales y criollos mexicanos de jitomate (S. lycopersicum) (15 materiales) y chile (C. annuum) (20 materiales), y evaluar la expresión de síntomas mantenidos bajo condiciones de invernadero hasta etapa de floración.
Inoculación de ToBRFV. Se utilizaron dos aislados del virus proveniente del Estado de México (EM-JI2021) y Colima (C-JI2021). Los aislados se inocularon en un total de 15 genotipos de jitomate, 13 comerciales y dos criollos mexicanos y, 20 de C. annuum, cuatro variedades, 15 genotipos y un criollo. Las inoculaciones se realizaron en condiciones de invernadero en plantas de 56 días de edad. Previo a la inoculación, se aplicó Imidacloprid (1.5 mL L-1) como medida preventiva a insectos vectores. La inoculación se realizó en la segunda hoja más joven, asperjando carborondum y posteriormente se aplicó el buffer de fosfatos con el macerado del tejido de jitomate infectado con el virus (dos aislados). Se consideró cinco plantas por genotipo, 150 plantas de jitomate (para ambos aislados) y 200 plantas de chile. Las plantas testigo (tres plantas por material vegetal) se aislaron para evitar contaminación. Las variables evaluadas fueron: incidencia del virus en plantas y se registró el tipo de síntomas en los materiales de jitomate y chile. Tejido de hojas jóvenes fueron colectadas a los 61 días después de la inoculación, se registró fotográficamente y se preservaron a -20 °C tejidos macerados con nitrógeno líquido para el estudio molecular.
Extracción de ARN y RT-PCR. Se realizó la extracción de los ácidos nucleicos con el método de CTAB 2 % (Yu, 2012; modificado por CP-LANREF, 2021). Las concentraciones y calidades del ARN se midieron en el NanoDrop 2000 (Thermo Fisher Scientific 2000, EUA). Para la RT-PCR se utilizaron los oligos ToBRFV-F 5-AACCAGAGTCTTCCTATACTGGGAA-3 y ToBRFV-R 5’CTCWC-CATCTCTTAATAATCTCCT-3 que amplifica parte de la subunidad pequeña de la replicasa RpRd con 475 pb (SENASICA, 2018). La retrotranscripción y reacción en cadena de la polimerasa (RT-PCR) se realizó en el termociclador T-100 (BioRad). Para la retrotranscripción (RT) se utilizó el oligo R (10uM), agua y 2 µL de ARN (50 ng µL-1) para obtener un volumen de 16.375 µL y se incubó a 85 °C por 3 min. Posteriormente, se adicionó a cada reacción previa, la mezcla de: Buffer-RT (5X), dNTPs (10mM), RNAsin (40 U µL-1) y M-MLV-RT (200 U µL-1) con un volumen de 8.625 µL. La mezcla se incubó a 44 °C por 60 min y 92 °C por 10 min. Para PCR, se utilizó MgCl2, dNTPs, los oligos, Taq polimerasa, agua libre de nucleasas y cDNA en un volumen final de 25 µL. Las condiciones fueron desnaturalización inicial 98 °C por 90 s, desnaturalización 98 °C por 10 s, alineamiento 55 °C por 20 s, extensión a 72 °C por 40 s, extensión final a 72 °C por 5 min y finalmente 72 °C (SENASICA, 2018). Los productos de PCR se analizaron por electroforesis en gel de agarosa al 1 % teñidos con bromuro de etidio y se visualizaron con luz UV en fotodocumentador (UVP, Biolmaging Systems, Epi Chemi II Darkroom).
Análisis filogenético. Productos de PCR (24 muestras) se enviaron a secuenciar a Macrogen® (Seúl, Corea). Las secuencias (ambos sentidos) se limpiaron y eliminaron los extremos con el programa SeqAssem (https://science.do-mix.de/software_seqassem.php). Con las secuencias se realizaron consenso, para identificar y comparar la homología con secuencias del banco de genes (Genbank) del Centro Nacional para la Información Biotecnológica (NCBI). Se seleccionaron secuencias de genomas completas de ToBRFV de México (tres secuencias) y de otros países (30 secuencias) del banco de genes para realizar el alineamiento con secuencias del fragmento del gen empleado en esta investigación. Se realizó el alineamiento con Mega 11.0.13 y Geneious 2023.0.4 (www.geneious.com) para determinar la variabilidad entre secuencias. Se seleccionaron 34 secuencias virales con enfoque epidemiológico. (Cuadro 1).
Síntomas en jitomate y chile asociados a ToBRFV. Todos los materiales comerciales y criollas de jitomate expresaron síntomas a los 10 días después de la inoculación de los aislados EM-JI2021 y C-JI2021. Los síntomas observados en jitomate inducidos por ToBRFV fueron principalmente mosaicos ligeros a severos. También se observó deformación y reducción foliar severa con remanencia de nervadura (Cuadro 2 y Figura 1). En algunas variedades se observó aclaramientos de nervaduras.
Zhi-Yong et al. (2021) evaluó 50 cultivares de jitomate y ningún material expresó resistencia al ToBRFV, mostrando diferentes síntomas como mosaico leve a severo, formación de ampollas en hojas, necrosis en sépalos y pedicelos, deformación y, en fruto manchas amarillas, así como lesiones necróticas con rugosidad marrón. Además, inoculó C. annumm, Nicotiana benthamiana, N. tabacum, Solanum melongena y S. tuberosum cv. Kexin 1, donde observó síntomas de necrosis en hojas inoculadas y tallos, así como enanismo.
En chile, todos los materiales manifestaron una diversidad de síntomas después de la inoculación, principalmente reacción de hipersensibilidad con caída de la hoja inoculada. Inicialmente se observó en hojas inoculadas lesiones necróticas y posteriormente, desprendimiento de la hoja. En algunos materiales se observó
Cuadro 1 Secuencias completas de Tomato brown rugose fruit virus obtenidas del banco de genes (NCBI) utilizadas para el alineamiento y comparados con secuencias de ToBRFV de la investigación.
| No. de accesión | Pares de bases | Hospedante | hTejido | hLugar |
|---|---|---|---|---|
| MZ945420.1 | 6379 | Solanum lycopersicum | Belgica | |
| OQ674194.1 | 6374 | S. lycopersicum | Canada | |
| OQ674195.1 | 6242 | S. lycopersicum cultivar Yari | Canada | |
| OM515230.1 | 6375 | S. lycopersicum | Netherlands | |
| OM515231.1 | 6373 | S. lycopersicum | United Kingdom | |
| OM515232.1 | 6373 | S. lycopersicum | United Kingdom | |
| MZ323110.1 | 6394 | S. lycopersicum | Jordan | |
| MK648157.1 | 6388 | Capsicum annuum | Jordan | |
| KT383474.1 | 6393 | S. lycopersicum | Jordan | |
| MN882030.1 | 6379 | S. lycopersicum | Egipto | |
| MN882031.1 | 6379 | S. lycopersicum | Egipto | |
| MW349655.1 | 6379 | C. annuum cultivar Tampiqueno | México | |
| OM515233.1 | 6369 | S. lycopersicum | Semillas | Peru |
| OM515258.1 | 6376 | S. lycopersicum | Semillas | Peru |
| OM515256.1 | 6361 | S. lycopersicum | Semillas | China |
| MT018320.1 | 6392 | S. lycopersicum | China | |
| OM515237.1 | 6377 | S. lycopersicum | Semillas | Israel |
| OM515234.1 | 6371 | C. annuum | Semillas | Israel |
| OM515257.1 | 6367 | S. lycopersicum | Semillas | Israel |
| OM515266.1 | 6364 | S. lycopersicum | Semillas | Israel |
| OM515250.1 | 6371 | S. lycopersicum | Semillas | Israel |
| OM515261.1 | 6376 | S. lycopersicum | Fruto | Belgium |
| OM515265.1 | 6375 | S. lycopersicum | Fruto | Belgium |
| MN815773.1 | 6354 | S. lycopersicum | Grecia | |
| OM305070.1 | 6386 | S. lycopersicum | switzerland | |
| MT107885.1 | 6386 | S. lycopersicum | Turquía | |
| OM515234.1 | 6371 | C. annuum | Semillas | Israel |
| OM782671.1 | 6356 | S. lycopersicum | Mexico | |
| OM892675.1 | 6384 | S. lycopersicum | Fruto importado | Mexico |
| OM892676.1 | 6381 | S. lycopersicum | Semillas | Peru |
| OM892677.1 | 6392 | S. lycopersicum | Fruto de tienda | USA |
| OM892678.1 | 6393 | S. lycopersicum | Fruto importado | Peru |
| OM892679.1 | 6375 | S. lycopersicum | Fruto de tienda | USA |
| OM892681.1 | 6357 | S. lycopersicum | Hoja | USA |
síntomas severos como deformación en follaje apical, necrosis en tallo, necrosis en las nervaduras y lesiones necróticas en hojas no inoculadas (Cuadro 2, Figura 2). Estos síntomas coinciden con Fidan et al. (2021) donde observaron necrosis en hojas inoculadas, lesiones necróticas en tallo y amarillamiento en hojas.
Variabilidad en aislados ToBRFV. Del total de genotipos de jitomate y chile inoculados, se enviaron a secuenciar cinco muestras de chile, una inoculada con
Cuadro 2 Síntomas de Tomato brown rugose fruit virus en materiales comerciales y criollos de jitomate y chile expresados bajo condiciones de invernadero.
| Nombre | Cultivo | Síntomas | Nombre | Cultivo | Síntomas |
|---|---|---|---|---|---|
| Santawest | Jitomatey | MM | Conga | Chilez | A |
| Citali | Jitomate | MM, LF, LR | Fascinato | Chile | A |
| IR143466 | Jitomate | SM, LF, LR | Felicitas | Chile | NLNI |
| Sicilia | Jitomate | SM, LF, LR | Botaron | Chile | A, SN |
| Sahariana | Jitomate | SM, LF, LR | Godzilla | Chile | NLNI |
| Altius | Jitomate | SM, LF, LR, GI SM, LF, LR, CV | Kathia | Chile | A |
| Don R | Jitomate | MS, LF, LD | Almuden | Chile | LD, NLNI, DAA, SN, NLN |
| Nebula | Jitomate | MM, LF | Bachia | Chile | LNI |
| Volcano | Jitomate | MM, LF, CV | Cannon | Chile | A |
| Ametrino | Jitomate | MM, LF, CV | Gina | Chile | NLNI, SN |
| Angelle | Jitomate | SM, LF, LD, CV | Confidaro | Chile | A |
| Olmeca | Jitomate | SM, LF, LR | |||
| UAM-X | Jitomate | SM, LF, LR, CV | Cayenne | Chile | DAA, NL |
| Rio Grande | Jitomate | SM, LF, CV | |||
| Totolapan, Mor. | Jitomate | MM | mulato | Chile | A |
| Cavanna | Chile | A | Serrano | Chile | SM, GI |
| Shir | Chile | A | Zongolica | Chile | MM, CR |
| Orangela | Chile | A | Chile pasilla | Chile | A |
| Manzano-Ver | Chile | A |
yJitomate. ML: mosaico ligero; MS: Mosaico severo; DF: Deformación foliar; AF: Apollamiento foliar; RF: Reducción foliar; AN: Aclaramiento de nervaduras; IV: Islas verdes.
zChile: A: Asintomática; ML: mosaico ligero; MS: Mosaico severo; DF: Deformación foliar; LN: lesiones necróticas en hojas; LNI: lesiones necróticas en hojas no inoculadas; NT: Necrosis tallo; DFA: Deformación área apical; NN: necrosis nervaduras en hojas; AF: Apollamiento foliar; AN: Aclaramiento de nervaduras; IV: Islas verdes; AC: anillos concéntricos.
C-JI2021 y cuatro inoculadas con EM-JI2021 y, 19 muestras de jitomate, seis muestras con EM-JI2021 y 13 con C-JI2021. Las secuencias en estudio se alinearon con el 80.2 % del total del fragmento del gen parcial del RpRd (475 pb). Las secuencias mostraron una identidad con accesiones del banco de genes con una homología que osciló entre 99.74 al 100 % incluyendo la secuencia del virus originalmente descrito por Salem et al. (2016) (Cuadro 3, Figura 3). Las secuencias que mostraron una identidad 99.74 y 99.75 % fueron muestras inoculadas con EM-JI2021 (Cuadro 3). Todas las secuencias se alinearon con accesiones de USA y Canadá (Cuadro 3). Cinco secuencias con identidad menor al 100 % se alinearon con OQ6741195.1 de Canadá y con una secuencia de USA (MT002973.1).
De acuerdo al análisis de alineamiento basado en el genoma completo de 34 secuencias de ToBRFV y el genoma parcial de la replicasa de las 24 secuencias

Figura 1 Síntomas en materiales comerciales y criollas de jitomate inoculados con Tomato brown rugose fruit virus. Síntomas de mosaico leve (IR143466, Nebula, Ametrino, Rio Grande, Citlali, Altius); mosaico severo, deformación y aclaramiento de nervaduras principales de la hoja (Volcano, Criollo-X y Angelle); planta sana (Angelle testigo).
de esta investigación, se observó variabilidad a nivel nucleótido en seis secuencias obtenidas de plantas inoculadas con EM-JI2021. Se observó dos cambios: en el nucleótido c.2,278A>T y c.2355T>C (Figura 3). Las secuencias en estudio se alinearon entre la posición 2,124 al 2,500 correspondiente al gen de la replicasa. Las substituciones de nucleótidos también se observaron en las accesiones MW349655.1 (tejido de chile), OQ674195.1 (hojas de jitomate), y OM515234.1 (semillas de chile), que corresponden a México, Canadá e Israel. Sin embargo, en otra investigación no se encontraron mutaciones de ToBRFV después de un proceso de inoculación en jitomate y chile (Eichmeier et al., 2023). Estos resultados concuerdan con un trabajo que reporta de 2 a 39 sustituciones de nucleótidos a nivel del genoma completo (Abrahamian et al., 2022). Si bien los cambios de nucleótidos son bajos, estos cambios podrían eventualmente generar variabilidad en el comportamiento patogénico y/o epidemiológico del virus. Esto es sustentado con

Figura 2 Síntomas en chile (Capsicum annumm) inoculados con Tomato brown rugose fruit virus. A-C) Síntomas de deformación apical, necrosis en tallo y nervaduras en hojas de Almuden; D) Síntomas de deformación apical, ligera necrosis en nervaduras en Cayenne; E) Síntomas de deformación apical, mosaico, lesiones necróticas en hojas no inoculadas en Felicitas.
lo observado en plantas de jitomate inoculados con el aislado del Edo. de México (EM-JI2021), que se registró mayor altura y diámetro del tallo que las plantas inoculadas con el aislado de Colima (C-JI2021) (Datos no mostrados). También se observó síntomas en jitomate inoculados con el aislado C-JI2021 se asociaron a los síntomas europeos, los cuales fueron más severos que el aislado del Edo. de México (Salem et al., 2016).
La importancia que tiene ToBRFV desde un diagnóstico temprano del virus hasta el impacto en producción en jitomate y chile es relevante. Se han realizado
Cuadro 3 Porcentaje de cobertura e identidad de dos aislados de ToBRFV inoculados en un total de 35 genotipos de jitomate y chile en condiciones de invernadero.
| Aislado | Variedad / criolloz | Cultivo | Origen aislado | Longitud pb | Cobertura | Identidad | Aislado homologado | |
|---|---|---|---|---|---|---|---|---|
| Accesión | País | |||||||
| EM-JI2021 | Almuden | Chile | Edo. México | 398 | 100 | 100 | OQ674195.1 | Canada |
| Mulato | Chile | Edo. México | 390 | 100 | 99.74 | OQ674195.1 | Canada | |
| Congo | Chile | Edo. México | 391 | 100 | 99.74 | MT002973.1 | USA | |
| Gina | Chile | Edo. México | 381 | 98 | 100 | MT002973.1 | USA | |
| Santawest | Jitomate | Edo. México | 402 | 100 | 99.75 | OQ674195.1 | Canada | |
| Gotzilla | Jitomate | Edo. México | 383 | 100 | 100 | MT002973.1 | USA | |
| Altius | Jitomate | Edo. México | 385 | 100 | 99.74 | OQ674195.1 | Canada | |
| Don R | Jitomate | Edo. México | 407 | 100 | 100 | OQ674195.1 | Canada | |
| Sahariana | Jitomate | Edo. México | 392 | 100 | 99.74 | OQ674195.1 | Canada | |
| Nebula | Jitomate | Edo. México | 408 | 99 | 99.75 | OQ674195.1 | Canada | |
| C-JI2021 | Manzano | Chile | Colima | 402 | 100 | 100 | MT002973.1 | USA |
| Santawest | Jitomate | Colima | 410 | 100 | 100 | MT002973.1 | USA | |
| Altius | Jitomate | Colima | 407 | 100 | 100 | MT002973.1 | USA | |
| Sicilia | Jitomate | Colima | 411 | 100 | 100 | MT002973.1 | USA | |
| IR143466 | Jitomate | Colima | 418 | 100 | 100 | MT002973.1 | USA | |
| Don R | Jitomate | Colima | 408 | 100 | 100 | MT002973.1 | USA | |
| Ametrino | Jitomate | Colima | 384 | 100 | 100 | MT002973.1 | USA | |
| Olmeca | Jitomate | Colima | 399 | 100 | 100 | MT002973.1 | USA | |
| Nebula | Jitomate | Colima | 404 | 100 | 100 | MT002973.1 | USA | |
| Xochimilco | Jitomate | Colima | 384 | 100 | 100 | MT002973.1 | USA | |
| Angelle | Jitomate | Colima | 399 | 100 | 100 | MT002973.1 | USA | |
| Río Grande | Jitomate | Colima | 408 | 100 | 100 | MT002973.1 | USA | |
| Totolapan | Jitomate | Colima | 388 | 100 | 100 | MT002973.1 | USA | |
| Citlali | Jitomate | Colima | 385 | 100 | 100 | MT002973.1 | USA | |
Z Nombres en cursiva son materiales criollos. En negrita secuencias con homología <100%.
estudios de métodos de extracción del ARN para un diagnóstico temprano y preciso en el análisis molecular (Zamora-Macorra et al., 2023), hasta alternativas de manejo del ToBRFV con bajo impacto ambiental como el uso de Beauveria peruviencis, Trichoderma longibrachiatum y Pseudomonas sp. (Ramos-Villanueva et al., 2023). No obstante, estudios sobre la expresión de síntomas en los diferentes materiales disponibles en el mercado, así como conocer la variabilidad de ToBRFV que existe en campo, da la pauta para conocer mejor la epidemiología del virus y su comportamiento de acuerdo al cultivo y condiciones climáticas que se establecen. Por ello, es importante hacer estudios de la diversidad del virus en campo e invernadero a través del tiempo y asociar la expresión de síntomas con la variabilidad a nivel nucleótidos del virus.
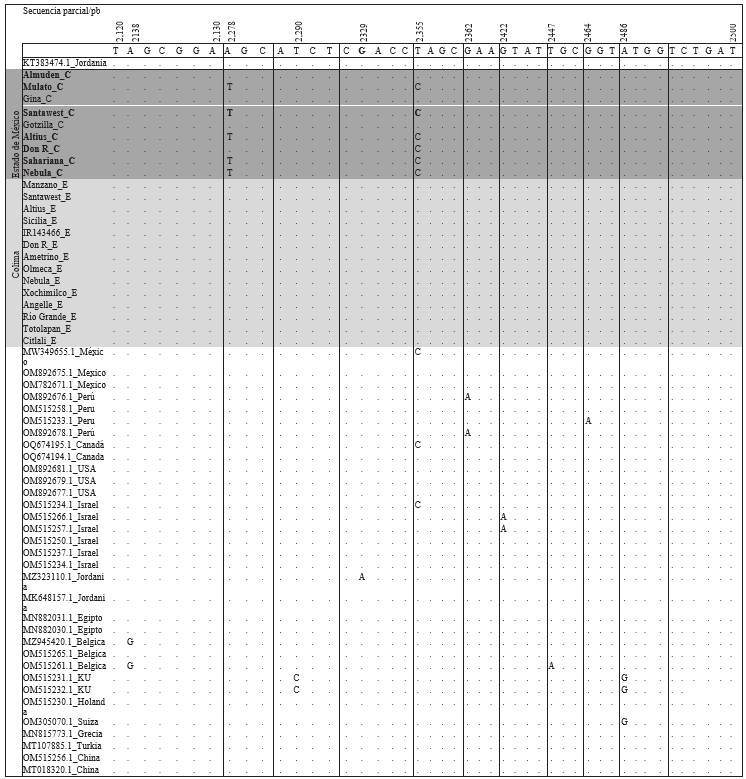
Figura 3 Alineamiento de la secuencia parcial del gen de la replicasa del Tomato brown rugose fruit virus de 24 secuencias de aislados obtenidos de 35 genotipos inculados con los aislados EM-JI2021 y C-JI2021, y de 34 secuencias de diferentes países productores de jitomate y chile. Alineamiento realizado en Geneious.
Conclusiones
Todos los materiales de jitomate expresaron síntomas de mosaicos, pero sin observar síntomas severos como los reportados por Salem et al. (2016), excepto en plantas de jitomate inoculados con el aislado C-JI2021. En chile se registraron lesiones en las hojas inoculadas (reacción de hipersensibilidad), así como lesiones necróticas en tallo, deformación apical, necrosis en nervaduras de las hojas y condición asintomática.
De acuerdo al análisis de alineamiento de dos aislados de ToBRFV (Edo. de México y Colima), comparado con 34 aislados disponibles en el banco de genes del NCBI, se observó en cinco secuencias de aislados del Edo. de México (EMJI2021) la sustitución de dos nucleótidos, nucleótido c.2,278A>T y c.2355T>C. Las accesiones MW349655.1 (México), OQ674195.1 (Canadá) y OM515234.1 (Israel) coincidieron con el aislado EM-JI2021 (inoculado en la variedad Don R) en cambio de un nucleótido c.2355T>C. En este trabajo se encontraron un total de tres haplotipos. Este es el primer trabajo que reporta variabilidad de ToBRFV en un tiempo corto en condiciones controladas. Esta información puede ser relevante para estudios de protección cruzada.

