











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
Introducción
La enfermedad conocida como mancha foliar causada por Corynespora cassiicola (Berk. y Curt.) Wei, es importante a nivel mundial debido a que ocasiona pérdidas significativas en la producción de hortalizas de importancia económica como el tomate (Solanum lycopersicum L.) (Gao et al., 2020). El patógeno afecta el crecimiento de las plantas en cualquier etapa de desarrollo, se observan lesiones necróticas con patrones de halo amarillo circundante en las hojas (Moo et al., 2022). El método más utilizado para el control de la enfermedad es el uso frecuente de fungicidas, sin embargo, además de los costos económicos, la gran dependencia de estos agroquímicos aplicados en dosis más altas, puede ser un riesgo para la salud humana y para el medio ambiente (Santovito et al., 2018), además, aceleran el proceso de resistencia en los patógenos, C. cassiicola está catalogado por el Comité de Acción de Resistencia a Fungicidas como un patógeno de “alto riesgo” debido a su desarrollo de resistencia a fungicidas (Zhu et al., 2018).
Por lo anterior se requieren opciones de manejo amigables con el ambiente sin acelerar la resistencia del patógeno. Una alternativa es el uso de rizobacterias pertenecientes al género Bacillus con potencial biocontrolador contra patógenos, con mecanismos como la competencia de espacio y nutrientes, la producción de lipopéptidos con capacidad de formar biopelículas protectoras y alteración de las membranas celulares de los microorganismos, así como actividades enzimáticas con capacidad de degradar la pared celular de los patógenos, sin efectos colaterales (Leal et al., 2015).
Con respectó a la actividad antagónica por competencia de espacios y nutrientes, Sosa et al. (2012) reportaron que Bacillus subtilis cepa F8 y cepa K47 presentaron más del 56 % en la inhibición in vitro de los patógenos Macrophomina phaseolina, Colletotrichum gloeosporioides, Helminthosporium sp. y Alternaria sp. En cuanto al modo de acción indirecto Bacillus spp. actúa por medio de la producción de lipopéptidos cíclicos (LPC), moléculas anfifílicas activas en la membrana con diversas actividades antimicrobianas y funciones antifúngicas importantes (Ongena y Philippe, 2008).
Bacillus spp. produce comúnmente tres familias biosintéticas principales de LPC utilizados como agentes de control biológico: surfactinas, iturinas y fengicinas. La familia surfactina destaca por la producción de biopelículas y emulsiones, evitando la adhesión y colonización de patógenos, de igual forma puede alterar la membrana celular del hongo provocando su desestabilización llevándolo a la muerte, tambien actúa como secuestrador de iones requeridos en el crecimiento y supervivencia del fitopatógeno (Sałek y Euston, 2019; Dong et al., 2022). Mejía et al. (2016) reportarón que B. subtilis F8 mostró presencia de genes relacionados con la biosíntesis de surfactinas e iturinas y antagonismo frente los patógenos Fusarium equiseti y F. solani, por otra parte, en B. subtilis JN005 se detectaron genes que codifican la síntesis de metabolitos secundarios de surfactina y fengicina, se aislaron los lipopéptidos y se probó in vitro contra Magnaporthe oryzae en plantas de arroz, se reporta una reducción del más del 17 % de la enfermedad causada por el patógeno (Zhu et al., 2023), así mismo en B. pumilis se detectó la presencia del gen de la surfactina y la inoculación de la bacteria en semillas protegió significativamente a las plántulas de sorgo de la infección por Rhizoctonia solani (Kumar et al., 2023).
Además, algunas cepas de Bacillus presentan actividad de enzimas quitinolíticas que hidrolizan los enlaces glicosídicos β -1,4 en la quitina el cual forma el componente principal de la pared celular de los hongos, contribuyendo de manera importante a la actividad antagónica de las cepas bacterianas contra los hongos patógenos de las plantas (Kalai-Grami et al., 2014). B. subtilis str. SV41 suprimió más de un ٨٧ ٪ el marchitamiento por Fusarium oxysporum f. sp. lycopersici en plantas de tomate, la cepa mostró presencia del gen ChiA (Aydi et al., 2017). Por otra parte, Rodas et al. (2009) reportan que las cepas LUM, B001, B003, B013, B015 y B065 pertenecientes al género Bacillus produjeron una actividad quitinolítica de más de 2 cm (halo de hidrólisis) en medio mínimo suplementado con 5 % de quitina coloidal, mientras que Brzezinska et al. (2020)) mencionan que B. subtilis B3 produjo una actividad quitinolítica de más de 4 U mL-1 utilizando como sustrato quitina coloidal. Así mismo, Le Quoc (2023) reportó que 25 aislados de bacterias inhiben el crecimiento de C. cassiicola, entre 39.8 % y 62.6 %, todos los aislados demostraron producir β-1, 3-glucanasa y quitinasas, siendo la cepa B. siamensis TV16 que exhibió mayor actividad antifúngica. El objetivo de este estudio fue evaluar la actividad biosurfactante y quitinolítica de dos cepas de B. subtilis nativas de Yucatán y su antagonismo contra C. cassiicola aislada de tomate.
Materiales y métodos
Material biológico
Se utilizó B. subtilis F8 y B. subtilis K47 aislados previamente de la península de Yucatán (Sosa et al., 2012). Para la obtención de los inóculos las bacterias se cultivaron en caldo nutritivo (CN) durante 10 días en agitación a 180 rpm a 30 °C (Incubator shaker ZWY 200D, Shanghái China), el sobrenadante (filtrado libre de células) y el paquete celular (esporas) se recuperaron por centrifugación a 3000 rpm por 15 minutos (Centrífuga Solbat C600, Puebla México). El filtrado libre de células se almacenó a -18 °C hasta su uso, y el paquete celular se ajustó a una DO de 0.8 a 550 nm en un espectrofotómetro (Ray Leigh, UV2601, China). La cepa de Corynespora cassiicola se aisló de plantas de tomate con síntomas típicos de la enfermedad (Moo et al., 2022), y se reactivó en medio PDA.
Antagonismo in vitro por confrontación directa
La actividad antifúngica in vitro se evaluó mediante la técnica de cultivo dual. Se colocó un disco de 0.5 cm de diámetro de micelio de C. cassiicola en un extremo de la caja Petri con medio PDA, 24 horas después se inoculó con una asa bacteriológica, una colonia aislada de cada cepa bacteriana en el extremo opuesto al hongo. Las cajas se incubaron a 28 °C durante 15 días y se determinó el crecimiento micelial (cm), se evaluó el crecimiento fúngico (cm2) midiendo el área total del crecimiento del hongo utilizando el programa imageJ y se calculó el porcentaje de inhibición utilizando la fórmula reportada por Abbott (1925):
Donde R1: valor promedio del radio de la colonia control y R2: es el valor promedio del radio del micelio fúngico que crece en la placa antagonista.
Ensayo en hojas desprendidas inoculadas con esporas o filtrado libre de células
Se realizó de acuerdo a reportes previos (Bañuelos, 2007; Parra et al. 2011; De la Caridad et al. 2017), se recolectó el segundo foliolo de la tercera hoja a partir del ápice de plantas de tomate variedad Pony express crecidas en condiciones de campo, en una cámara de flujo laminar se trabajó con la técnica de cámara húmeda en cajas Petri estériles de 100 por 15 mm , en la cual se dispuso un foliolo de aspecto sano, previamente desinfectado con hipoclorito de sodio al 2 % y lavada tres veces con agua destilada estéril. Tanto para el ensayo con la solución de esporas (DO = 0.8), como para el filtrado libre de células se asperjaron 200 μL de cada bacteria por tratamiento en la zona adaxial de la hoja y se dejó secar 15 min, posteriormente, se colocó un disco de 0.5 cm de diámetro de micelio de C. cassiicola en dos extremos de la zona media de la parte adaxial de la hoja. Como testigo se utilizaron hojas sin aspersión de esporas o de filtrado libre de células. Se realizaron tres repeticiones por tratamiento (una cámara húmeda como unidad experimental). Las cámaras fueron humedecidas homogéneamente con 1 mL de agua destilada estéril cada dos días. Al quinto día se evaluó el área afectada y área sana mediante el programa Image J 64 bits para Windows.
Evaluación de la capacidad biosurfactante de Bacillus subtilis
Se le evaluó la capacidad de emulsión en B. subtilis cepa F8 y cepa K47 de acuerdo a lo descrito por Bodour et al. (2004), en tubos de ensayo de 13 x 100 mm, se agregó 2 mL del filtrado libre de células y 2 mL de aceite mineral y se mezcló en la velocidad máxima durante 2 min con un vortex (Ika Vortex 3, Madrid España), las muestras se mantuvieron en oscuridad durante 24 h. El índice de emulsión (E24) se calculó de acuerdo a la siguiente ecuación:
Donde AEF: Altura de emulsión formada, ATS: altura total de la solución.
Extracción de ADN bacteriano
Las cepas bacterianas se cultivaron en caldo nutritivo y se mantuvieron 24 horas en agitación a 180 rpm a 30 °C (Incubator shaker ZWY 200D, Shanghái China). La extracción de DNA se realizó utilizando el kit de extracción Wizard Genomic, kit Wizard Genomic (Promega®, E.U.A) de acuerdo a las instrucciones del fabricante. La integridad del DNA se visualizó en gel de agarosa al 1 %. La concentración y pureza se determinó con un Nanodrop 2000c (Termo scientifíc, E.U.A.).
Detección de genes relacionados a la biosintesis de surfactina
Para determinar la presencia de genes relacionados a la biosíntesis de lipopéptidos srfAA y srf3 pertenecientes a la familia de la surfactina en B. subtilis cepa F8 y cepa K47, se realizó la amplificación por la PCR en un termociclador (Techne TC-312 E.U.A.), los iniciadores específicos utilizados se presentan en la Tabla 1. Se realizó a un volumen de reacción de 50 μL, se mezcló buffer PCR 1x, 1.5 mM de MgCl2, 0.2 mM de dNTPs, 0.5 μM de cada iniciador y 2 U de Dream Taq DNA Polymerase (Termo Scientific). Las condiciones de reacción de la PCR fueron con una denaturalización inicial a 95 °C durante 3 min, 35 ciclos: desnaturalización a 94 °C durante 1 min, temperatura de alineamiento de 60 °C por 1 min, extensión a 72 °C durante 1 min, y una extensión final de 72 °C por 5 min. Los productos de la PCR obtenidos se enviaron a secuenciar a la empresa Psomagen, Inc. E.U.A. Las secuencias obtenidas se analizaron con el software BioEdit Sequence Alignment Editor, los resultados se compararon con la base de datos del National Center for Biotechnology Information (NCBI), mediante el programa Basic Local Alignment Search Tool (BLAST).
Table 1: Primers used for detection of surfactin biosynthesis genes
| Gen | Iniciador | Secuencia (5' – 3') | Tamano (pb) | Tm (°C) | Referencia |
| srf3 | SUR3F | ACAGTATGGAGGCATGGTC | 441 | 60 | Athukorala et al., 2009 |
| SUR3R | TTCCGCCACTTTTTCAGTTT | ||||
| srfAA | SRFAF | TCGGGACAGGAAGACATCAT | 201 | 58 | Mora et al., 2011 |
| SRFAR | CCACTCAAACGGATAATCCTGA |
Tm: temperatura de alineamiento, pb: pares de bases del ADN. Tm: Anealling temperature, pb: AND base pairs.
Actividad quitinolítica de Bacillus subtilis F8 y K47
Se utilizó quitina coloidal al 10 % en medio mínimo como medio de inducción de acuerdo a lo reportado por Monreal y Reese (1969). Al sobrenadante obtenido se le determinó azucares reductores liberados por el método de Miller (1959). Las proteínas se cuantificaron por el método Bradford (1976) con una curva estándar de albumina sérica bovina (BSA). La unidad de actividad enzimática específica (UQ) se definió como la cantidad de enzima necesaria para liberar un micromol de N-acetilglucosamina, determinado como azucares reductores, por mg de proteína en las condiciones de la reacción (Chávez y Cruz, 1984). Los valores se interpolan en una curva tipo N-acetilglucosamina (NAG).
Resultados y discusión
Antagonismo in vitro de B. subtilis sobre C. cassiicola
B. subtilis cepa F8 y cepa K47 inhibieron el crecimiento micelial de C. cassiicola, el mayor efecto se observó con B. subtilis F8 (62.2 %) en comparación con el testigo, no se detectó diferencia significativa entre las cepas de bacterianas. Ambas cepas (F8 y K47) disminuyeron un 79.4 % y 75.6 % el área de crecimiento fúngico de C. cassiicola comparadas con el testigo. B. subtilis F8 mostró el porcentaje de inhibición mayor, siendo estadísticamente diferente a B. subtilis K47 y al testigo (Tabla 2).
Tabla 2 Antagonismo in vitro de Bacillus subtilis contra Corynespora cassiicola a los 15 días de confrontación.
Table 2. In vitro antagonism of Bacillus subtilis against Corynespora cassiicola after 15 days of confrontation.
| Tratamiento | Crecimiento micelial (cm) | Crecimiento fúngico (cm2) | Inhibición (%) |
| Bs F8 + C.c. | 2.15 ± 0.04 b | 14.70 ± 0.36 c | 72.96 ± 0.41 a |
| Bs K47 + C.c. | 2.45 ± 0.08 b | 17.48 ± 0.26 b | 69.14 ± 1.01 b |
| C.c. | 7.94 ± 0.11 a | 71.60± 0.25 a | 0 c |
| DMS | 0.385 | 1.833 | 3.472 |
Los datos son expresados como media ± EE. Medias con diferente letra en columnas indican diferencias significativas (Tukey, p ≤ 0.05). Data are expressed as mean ± SE. Means with different letters in columns indicate significant differences (Tukey, p ≤ 0.05).
Debido a la rápida proliferación de ambas bacterias se consume en menor tiempo los nutrientes presentes en el medio de cultivo, limitando los recursos necesarios para el crecimiento del hongo, por otra parte, ambas bacterias cubrieron mayores superficies, impidiendo la proliferación del patógeno y restringiendo su capacidad de expansión (Ghosh y Panja, 2021). En estudios previos, se reportó que B. subtilis F8 inhibió el crecimiento micelial un 63.8 % y 69.9 % de los patógenos Fusarium equiseti y Fusarium solani (Mejía et al., 2016).
Supresión de la mancha foliar usando esporas de B. subtilis en hojas desprendidas
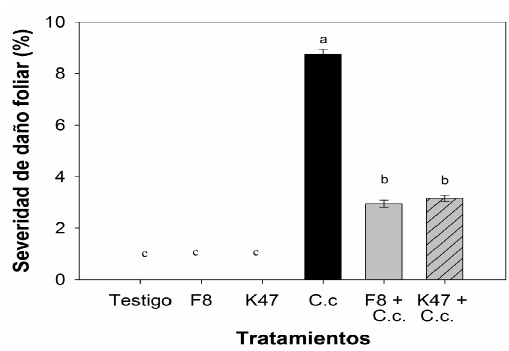
En cuanto a las hojas desprendidas tratadas con esporas de B. subtilis F8 y K47, se observó una reducción de los síntomas de la mancha foliar causada por C. casiicola (Figura 1), la inoculación con la cepa F8 redujo los síntomas de la enfermedad un 66.2 %, mientras que al ser tratadas con la cepa K47 se observó una reducción del 63.8 %, en comparación con el tratamiento infectado con C. casiicola y sin tratar con esporas bacterianas (Figura 2).
Figura 1 Síntomas de la enfermedad mancha foliar causada por C. cassiicola en hojas desprendidas de S. lycopersicum tratadas con esporas de B. subtilis. A) Testigo sin inoculación, B) B. subtilis F8, C) B. subtilis K47, D) C. cassiicola, E) B. subtilis F8 contra C. cassiicola, F) B. subtilis K47 contra C. cassiicola.

Figure 1: Symptoms of leaf spot disease caused by C. cassiicola on detached S. lycopersicum leaves treated with B. subtilis spores. A) Control without inoculation, B) B. subtilis F8, C) B. subtilis K47, D) C. cassiicola, E) B. subtilis F8 against C. cassiicola, F) B. subtilis K47 against C. cassiicola.
Supresión de la mancha foliar usando filtrado libre de células de B. subtilis en hojas desprendidas
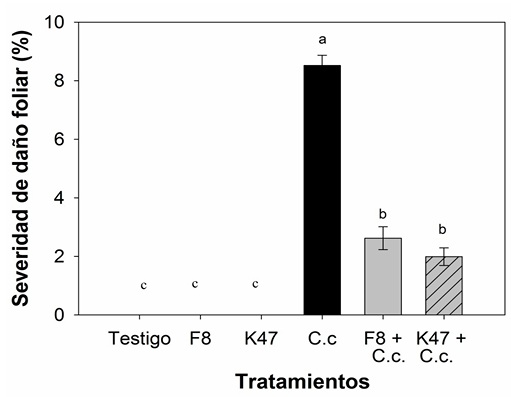
En las hojas desprendidas tratadas con el filtrado libre de células de B. subtilis cepa F8 y cepa K47, se vio una reducción del área afectada por C. casiicola (Figura 3), la inoculación con el filtrado de la cepa F8 redujo los síntomas de la enfermedad un 69.2 %, mientras que con el filtrado de la cepa K47 se observó una reducción del 76.6 %, en comparación con el tratamiento inoculado con C. casiicola y sin tratar con filtrados libres de celulas bacterianas (Figura 4).
Figura 3 Síntomas de la enfermedad mancha foliar causada por C. cassiicola en hojas desprendidas de S. lycopersicum tratadas con filtrado libre de celulas B. subtilis. A) Testigo sin inoculación, B) B. subtilis F8, C) B. subtilis K47, D) C. cassiicola, E) B. subtilis F8 contra C. cassiicola, F) B. subtilis K47 contra C. cassiicola.

Figure 3. Symptoms of leaf spot disease caused by C. cassiicola on detached S. lycopersicum leaves treated with B. subtilis cell-free filtrate. A) Control without inoculation, B) B. subtilis F8, C) B. subtilis K47, D) C. cassiicola, E) B. subtilis F8 against C. cassiicola, F) B. subtilis K47 against C. cassiicola.
Figura 4 Porcentaje de severidad de C. cassiicola en hojas de S. lycopersicum tratadas con filtrado libre de celulas de B. subtilis. F8: B. subtilis F8, K47: B. subtilis K47, C.c: C. cassiicola. Los valores son medias ± EE; barras con literales idénticas son estadísticamente iguales (Tukey, p ≤ 0.05).
En el presente estudio B. subtilis cepa F8 y cepa K47 mostrarón mantener la actividad antagónica in vitro reportada anteriormente contra otros fitopatógenos (Sosa et al., 2016; Mejía et al., 2019), esta actividad antifúngica se presentó en los bioensayos en hojas desprendidas, donde se observó una reducción del área afectada en los tratamientos inoculados con las bacterias, sin embargo, las hojas desprendidas mostraron mayor reducción en los síntomas de la enfermedad con el uso de filtrados libres de células, por lo que su acción antifúngica puede estar relacionado con la presencia de metabolitos secundarios y lipopéptidos cíclicos, algunas bacterias tienen la capacidad de producir sustancias antimicrobianas que pueden interferir con la replicación celular o dañar las membranas celulares del hongo, sin embargo, no todas las cepas presentan los mismos potenciales (Barboza et al., 2003;Becerra y Horna, 2016; Aydi et al., 2015; Trupo et al., 2023).
Detección de genes de la biosíntesis de surfactina
B. subtillis F8 y K47 produjeron por PCR amplicones de aproximadamente 200 y 440 pb, utilizando los iniciadores para los genes srfAA y srf3 respectivamente. La secuenciación de los fragmentos amplificados por la PCR permitió confirmar tres secuencias relacionadas con la biosintesis de surfactina (Tabla 3). Para B. subtilis F8 se identificó el gen SrfAD que codifica para una tioesterasa y el gen SrfAA que codifica para la surfactina sintetasa, en el caso de B. subtilis K47 se identificó el gen SrfAA. Estos genes identificados en B. subtillis cepa F8 y cepa K47 pertenecen a la familia surfactina. Las surfactinas poseen actividades antifúngicas y su interacción con las membranas celulares de los microorganismos patógenos conduce a la desestabilización de la estructura y muerte celular, de igual forma puede secuestrar iones metálicos requeridos para el crecimiento y supervivencia de los hongos lo que lleva a la inhibición de su crecimiento, además, forma una capa protectora que interfiere en la adhesión de los hongos en la superficie, lo que limita su capacidad de colonizar y proliferar (Sałek y Euston, 2019; Yang et al., 2020; Dong et al., 2022). Anteriormente Mejía et al. (2016) reportaron que B. subtilis F8 mostró presencia del gen Srf3, lo cual fue confirmada con las secuencias obtenidas (Tabla 3) de que dichos fragmentos corresponden a genes que codifican enzimas en la sintesis no ribosomal de surfactinas.
Table 3: Identification of genes related to surfactin biosynthesis.
| Cepa | Gen | Tamaño (pb) | Similitud con el GenBank (NCBI) | ||
| Proteína hipotética | Porcentaje de identidad | Accesión | |||
| F8 | srf3 | 433 | Tioesterasa de la biosintesis de surfactina (SrfAD) | 99.77 | CP156029 |
| F8 | srfAA | 202 | Surfactina sintetasa (SrfAA) | 100.0 | CP051463 |
| K47 | srfAA | 202 | Surfactina sintetasa - peptido no ribosomal (SrfAA) | 99.0 | CP046448 |
pb: pares de bases del ADN. pb: ADN base pairs.
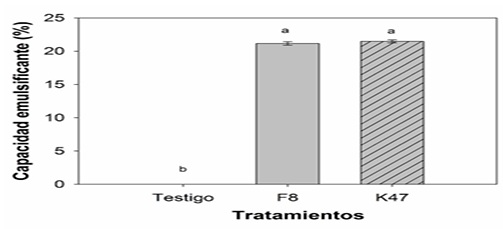
Actividad biosurfactante de Bacillis subtilis
Se detectó que tanto B. subtilis F8 y K47 poseen la capacidad de formar de emulsión de 21.15 % y 21.48 % respectivamente, en comparación al testigo, sin embargo, no se detectó diferencias significativas entre los tratamientos bacterianos (Figura 5). Por otra parte Al-Mutar et al. (2023) reportaron que el filtrado libre de células bacterianas y el extracto de lipopéptido extracelular de B. subtilis DHA41 mostraron inhibir significativamente el crecimiento micelial de Fusarium oxysporum. f. sp. niveum, Didymella bryoniae, Sclerotinia sclerotiorum, Fusarium graminearum y Rhizoctonia solani. El extracto lipopeptídico demostró una actividad emulsionante además inhibió el crecimiento del micelio de F. oxysporum en un 86.4 % a 100 µg/ml. A través de observaciones en microscopio electrónico de transmisión se confirmó que el extracto de lipopéptidos alteraba la integridad celular de F. oxysporum.
Actividad quitinolítica de Bacillus subtilis
Ambas cepas de B. subtilis (F8 y K47) mostraron tener actividad enzimática con capacidad de degradar la quitina coloidal. La cepa K47 mostró tener mayor actividad quitinolitíca especifica (UQE) por proteína en la muestra (Tabla 4).
La actividad de la enzima quitinasa está involucrada en la degradación de la pared celular de los hongos, a través de la hidrólisis de los enlaces glucosídicos entre las unidades de N-acetilglucosamina en la quitina, el cual es el componente principal de la pared célula del hongo (Younes y Rinaudo, 2015; Philibert et al., 2017), comprometiendo su crecimiento y desarrollo, lo que condujo directamente a la reducción de los síntomas de la enfermedad de la mancha foliar en las hojas desprendidas tratadas con filtrado libre de células bacterianas. Un estudio realizado por Arun et al. (2024) se clonó de Bacillus aryabhattai el gen quitinasa en el plásmido pET 23a y se transfirió en E. coli Rosetta pLysS. La actividad quitinasa más alta en el polvo y las escamas de quitina insoluble alcanzó 875 U mg-1 y 625 U mg-1 respectivamente. La quitinasa demostró inhibición del crecimiento de Candida albicans, Fusarium solani y Penicillium chrysogenum.
Conclusiones
Bacillus subtilis F8 y K47 son cepas quitinolíticas productoras de surfactantes que inhibieron el crecimiento micelial y disminuyeron el daño foliar ocasionado por Corynespora cassiicola, por lo que presentan potencial prometedor como alternativa sustentable en el control de C. cassiicola en el cultivo de tomate.

