











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
En Ecuador, en la región Amazónica, la reforestación resulta una de las principales actividades que se promueven para recuperar e incrementar la cobertura arbórea; es un tema de preocupación para el sector forestal pues se registran tasas de deforestación anual de 0.67% y parte de las áreas de bosques nativos son empleadas para otras actividades económicas más rentables (Mejía et al., 2021). Son innumerables los esfuerzos que se realizan para gestionar proyectos con este propósito, en el que la producción de plantas de calidad en vivero tiene una participación determinante en la aclimatación y el establecimiento de las plantaciones forestales (Ureta et al., 2019).
El proceso de repoblación forestal es complejo ya que existen varios factores que condicionan el éxito de las plantaciones: preparación del terreno, cuidados culturales, ejecución adecuada y control de las actividades y, en gran parte, la calidad de la planta utilizada (Serrada-Hierro et al., 2005; Trejo y Contreras, 2004). El éxito de una plantación también está relacionado con la correcta selección de las especies y procedencias, así como con el manejo durante su ciclo productivo (Fontana et al., 2015).
La procedencia de la semilla es reconocida como el área geográfica y ambiental de donde se obtiene el germoplasma, dentro de la cual se ha desarrollado su constitución genética. La relevancia de la procedencia resulta del control genético que esta tiene sobre caracteres de comportamiento tales como supervivencia, crecimiento y producción de plantas (Fontana et al., 2015). Los factores genéticos de una especie pueden ser parámetros determinantes en la calidad de las plantas forestales en vivero y la selección de procedencias constituye una herramienta útil para lograr la máxima adaptación de la especie al sitio de plantación (Navarro y Palacios, 2004). La caracterización de materiales de diferentes orígenes geográficos brinda información necesaria para la selección de las fuentes de semilla superiores para los programas de mejora, así como en proyectos de forestación y/o repoblación (Fontana et al., 2015).
La calidad de una planta está determinada por el conjunto de atributos morfológicos y fisiológicos que le confieren mayor supervivencia, resistencia y crecimiento en campo, lo cual facilita información clave para seleccionar material idóneo con alta variabilidad genética para cada condición de sitio (Birchler et al., 1998; Rueda-Sánchez et al., 2013; Ureta et al., 2019). Se reconoce que la calidad de la planta puede comprometer la estabilidad de la repoblación a lo largo del tiempo y que son pocos los productores que realizan una evaluación para mejorar los sistemas de producción en vivero (Luis et al., 2004; Ortega et al., 2004; Rueda-Sánchez et al., 2013). De ahí que determinar los factores morfológicos y fisiológicos de calidad de las plantas producidas en vivero, debe ser una prioridad de los actores involucrados en los programas de establecimiento de plantaciones forestales.
Este trabajo se enfoca en Ochroma pyramidale (Cav. ex Lam.) Urb, conocida como balsa, debido a que es una especie nativa del trópico americano, que se distribuye desde el sureste de México, Centroamérica, Colombia, Venezuela, Ecuador, Perú, Bolivia y Las Antillas hasta Brasil (Toledo et al., 2019), de importancia por ser una especie pionera, de rápido crecimiento, que prospera en suelos degradados y que tiene alto valor comercial (Guerrero et al., 2021). La madera es muy valorada en el mercado internacional, es extremadamente liviana y ha adquirido interés comercial para la fabricación de piezas de aviones, juguetes y artesanías, siendo Ecuador el principal productor mundial de materia prima (Jiménez et al., 2017; Morstadt y Maridueña, 2015; Toledo et al., 2019). Por lo anterior, esta especie presenta altas potencialidades para fomentar el establecimiento de plantaciones forestales con fines comerciales en la región amazónica ecuatoriana.
Objetivos
El objetivo de este trabajo fue evaluar el efecto de tres procedencias (Pastaza, Napo y Orellana) sobre la morfología de las semillas, parámetros germinativos y características morfofuncionales de plántulas de Ochroma pyramidale cultivadas en vivero.
Materiales y métodos
Localización geográfica
El vivero se ubicó en la matriz principal de la Universidad Estatal Amazónica, km 2 1/2 vía Puyo-Tena Cantón Pastaza, provincia de Pastaza, donde predominan temperaturas máximas y mínimas promedio de 28 °C y 17 °C, respectivamente, con humedad relativa promedio de 85.2% (Estación meteorológica Davis Vantage Pro 2).
Procedencias
Se emplearon semillas de la especie O. pyramidale de tres procedencias de la Amazonía ecuatoriana: 1) Pastaza (1° 14' 36.1"S - 77° 37' 10.4" O, 950 m de altitud), ubicado en la Amazonía central, cantón Arajuno, Parroquia Curaray, Recinto San José de Curaray, con predominio de bosque siempreverde piemontano; 2) Napo (0° 56' 25.8" S 77° 39' 37.4" O, 577 m de altitud), ubicado en la Amazonía central, cantón Archidona, Parroquia San Pablo, Recinto El Para, con predominio de bosque siempreverde piemontano y 3) Orellana (0° 43' 05.2" S 76° 55' 23.0" O, 245 m de altitud), ubicado en la Amazonía baja, cantón Loreto, Parroquia Puerto Murialdo, Recinto Murialdo, con predominio de vegetación de tierras bajas (Fig. 1).
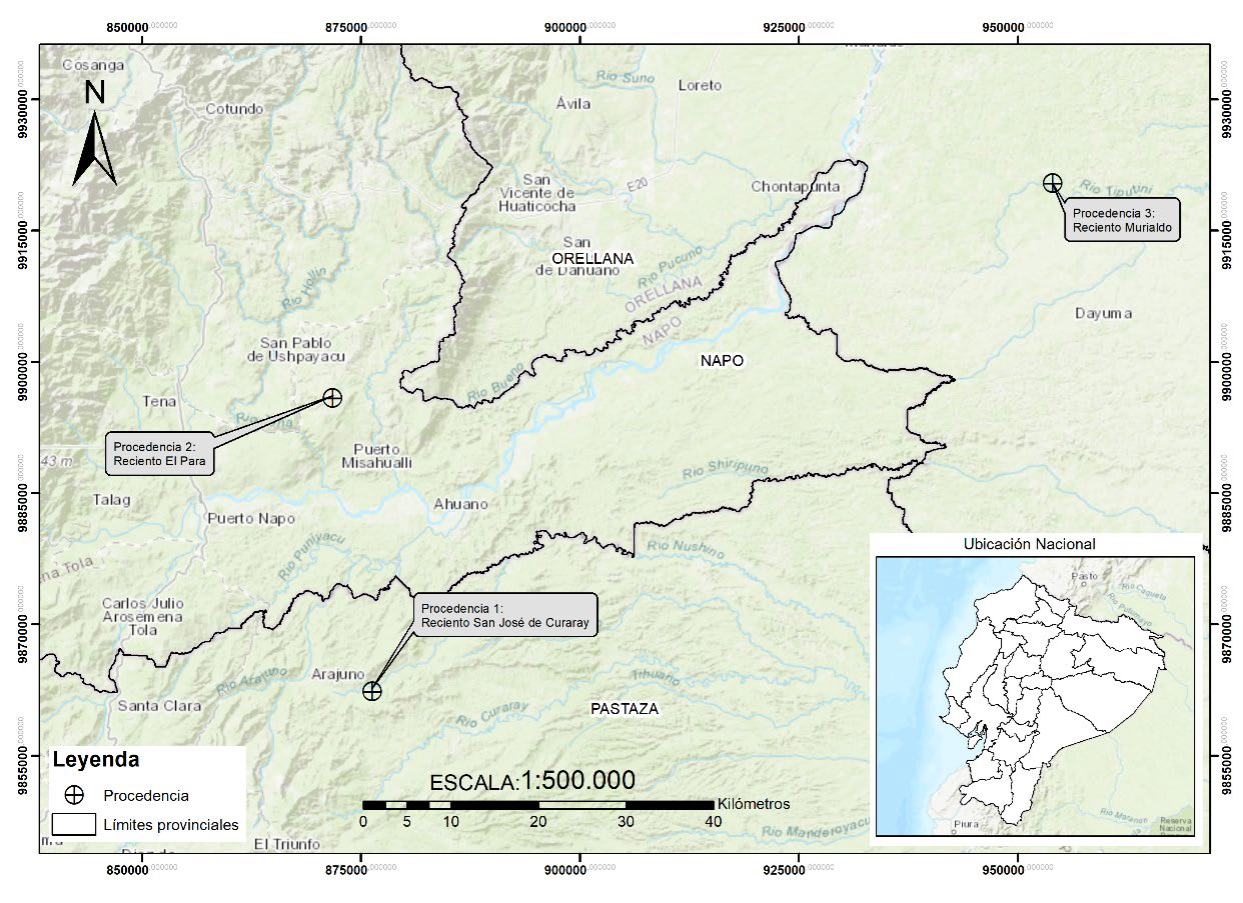
Recolección de semillas y ambiente edáfico
La recolección de las semillas se realizó a partir de la selección de diez individuos de cada procedencia (de Andrade et al., 2018), los cuales se encontraban aislados en remanentes de bosques naturales con presencia de frutos con semillas al momento de la recolección. La selección de los individuos se realizó tomando como criterio la evaluación de árboles semilleros, considerando los parámetros fenotípicos: fuste recto, árboles no bifurcados, dominancia del eje principal, ángulo de inserción de las ramas de 90°, forma de la copa circular y diámetro de copa promedio de 12 m (Ontaneda et al., 2017).
Se caracterizó el ambiente edáfico de cada procedencia a partir de las propiedades físicas y químicas del suelo: clase textural (% de arena, limo y arcilla), materia orgánica (M.O), pH, Ca2+, Mg2+, K+, P, N, Al3+ y acidez intercambiable (Al3+ H+). Las muestras de suelo fueron tomadas alrededor de cada árbol seleccionado, en cuatro puntos cardinales, a una distancia aproximada de 1 m desde la base. Una muestra fue tomada a diferente profundidad (cm): 0 - 10, 10 - 30 y 30 - 60, estas se mezclaron para obtener una sola muestra compuesta por cada árbol (Novoa et al., 2018; Bravo et al., 2017). El contenido de Ca2+, Mg2+, K+, P y Al3+ se realizó mediante la metodología de Olsen modificada (Bertsch, 1998), la acidez intercambiable (Al3+ H+) extraída con HCl (Molina, 2007), el pH se determinó a través de potenciometría (Cury, 2006), la materia orgánica por el método de digestión húmeda de Walkley & Black (Gonzalez-Pedraza et al., 2016) y la textura del suelo por el método de Bouyoucos (Bouyoucos, 1951).
Morfología de las semillas
Se realizaron mediciones morfológicas a una muestra masal por procedencias, conformada por tres submuestras al azar de 150 semillas de cada procedencia. El tamaño de la muestra se seleccionó a partir del criterio de selección de 100 semillas mínimo por muestra según investigaciones previas (Yu et al., 2017; de Andrade et al., 2018; Pozo-Gómez et al., 2019; Intanon et al., 2020). Se midió largo (L), ancho (W) y espesor (T), utilizando un calibrador digital o pie de rey marca Stanley con una resolución de ±0.02 mm.
Se determinaron los parámetros morfológicos de las semillas relacionados con el diámetro medio geométrico (Dg), relación largo/ancho (RLA) y área superficial (S) (Koocheki et al., 2007), según las ecuaciones 1, 2 y 3:
donde:
Dg: diámetro medio geométrico (mm)
L: largo (mm)
W: ancho (mm)
T: espesor de las semillas (mm)
RLA: relación largo/ancho
S: área superficial (mm2)
Manejo en vivero
Se emplearon bandejas plásticas de germinación de 42.8 cm × 61.5 cm que contenían 96 tubetes de plásticos rígido de 115 cm3 de capacidad. Se empleó un sustrato compuesto por una mezcla homogénea que contenía 80% de humus de lombriz y 20% de tamo de arroz. Las propiedades físicas y químicas del sustrato se muestran en la Tabla 1.
Tabla 1 Propiedades físicas y químicas del sustrato empleado en vivero para el cultivo de plantas de O. pyramidale.
| Sustrato | pH | MO (g/100g) | Nt (g/100g) | P (ppm) | K (g/100g) | PT (%) | Hum (g/100g) | Da (g/cm 3 ) |
| 80% Humus de lombriz+20% de tamo de arroz | 5.86 | 14.00 | 0.48 | 1.40 | 0.37 | 83.00 | 34.50 | 0.50 |
pH (potencial de hidrógeno); MO (materia orgánica); Nt (nitrógeno total); P (fósforo); K (potasio); PT (porosidad total); Hum (humedad) y Da (densidad aparente).
Se empleó un tratamiento pregerminativo que consistió en remojo en agua fría durante 24 horas, comúnmente utilizado para romper la latencia física (Camacho et al., 2018), ya que a pesar de estar en condiciones favorables son incapaces de germinar (Jiménez et al., 2017).
El diseño utilizado fue completamente al azar con tres tratamientos (procedencias) y la unidad experimental consistió en 100 semillas por tratamiento y tres repeticiones.
Se realizó el control de la germinación diariamente desde el día de la siembra hasta la finalización del período germinativo (30 días), el cual se definió al momento de no presentarse más emergencia. Se consideró una semilla germinada cuando comenzó a emerger la plúmula (Rossini et al., 2006). En el proceso de germinación se observó que la especie presentó germinación epígea. Se aplicaron tres riegos complementarios a capacidad de campo por semana.
Parámetros de germinación
Se calculó el porcentaje de pureza (P) como indicador físico de calidad de las semillas y un conjunto de parámetros de germinación relacionados con: capacidad germinativa (CG), tiempo medio de germinación (TMG), índice de Timpson (IT), valor útil (VU) y vigor de germinación (VG) ( Pece et al., 2010; Rodriguez et al., 2008; Staniak et al., 2021), según las ecuaciones 4, 5, 6, 7, 8 y 9.
donde
P: porcentaje de pureza (%)
p: peso de semillas puras (g)
I: peso de semillas impuras (g)
donde:
CG: capacidad germinativa (%)
ni: número de semillas germinadas
N: número total de semillas de la muestra
donde:
TMG: tiempo medio de germinación (días)
Tn: número de días transcurridos desde el inicio de la germinación hasta el día n
Nn: número de semillas germinadas en el día n
donde:
IT: índice de Timpson (%/día)
G: porcentaje de germinación de semillas en intervalos de dos días (%/día)
t: germinación total
donde:
VU: valor útil (%)
P: porcentaje de pureza (%)
CG: capacidad germinativa (%)
donde:
VG: vigor germinativo (%)
UM: valor máximo o pico que se presenta entre los valores producto de la división del porcentaje acumulado de germinación y la cantidad de días que tardó en obtenerse (%/día).
GDM: germinación media diaria, calculada como la razón entre el porcentaje final de germinación (PG) y el número de días transcurridos hasta llegar a ese valor (%/día)
Variables morfológicas y fisiológicas de calidad de plantas
Para el análisis de calidad de planta se midieron 15 plantas por procedencia a la edad de tres meses (Basave-Villalobos et al., 2020). Las variables morfológicas fueron: altura (H; cm) y diámetro en el cuello de la raíz (DCR; mm); la primera se determinó con una regla milimétrica y la segunda con un calibrador digital pie de rey; el peso seco de la parte aérea (PSA; g), de la raíz (PSR; g) y el peso seco total (PST; g) se registraron con una balanza analítica digital Sartorius con resolución de 0.1 mg previo secado de cada fracción de la planta en una estufa con circulación forzada de aire a 60 °C hasta peso constante. Se determinaron los índices morfológicos de calidad de la planta: relación parte aérea/radical (PA/PR), índice de calidad de Dickson (IQ), coeficiente de esbeltez (EZ), índice de lignificación (IL) y balance hídrico (BAP) (Birchler et al., 1998; Dickson et al., 1960; Rueda et al., 2014), a partir de las ecuaciones 10, 11, 12, 13 y 14.
donde:
PA/PR: relación parte aérea-radical
PSA: peso seco aéreo (g)
PSR: peso seco radical (g)
donde
EZ: coeficiente de esbeltez
H: altura de las plantas (cm)
DCR: diámetro en el cuello de la raíz (mm)
donde:
IL: índice de lignificación
PST: peso seco total (g)
PSH: peso húmedo total (g)
donde: BAP: balance hídrico
PSA: peso seco aéreo (g)
DCR: diámetro en el cuello de la raíz (mm)
PSR: peso seco radical (g)
donde:
IQ: índice de calidad de Dickson
PST: peso seco total (g)
EZ: coeficiente de esbeltez
PA/PR: relación peso seco aéreo-radical
Las variables fisiológicas se determinaron con un sistema portátil integrado de medidas fotosintéticas y de fluorescencia de clorofila con control de microclima (fluorómetro integrado iFL/Cpro-SD, ADC BioScientific Ltd., Herts, UK). Las mediciones se realizaron entre las 7 h 00 y las 10 h 00 (Mejía De Tafur et al., 2014). Se programó la cámara del equipo con a temperatura (Tch), humedad relativa (H2O Ref) y concentración de CO2 (CO2 Ref) en condiciones ambientales y con un pulso de luz de (QLeaf) de 1200 µmol m-2 s-1. Las variables fisiológicas que se midieron fueron: tasa de transpiración [E] (mmol H2O m-2 s-1), conductancia estomática [gs] (mol H2O m-2 s-1), tasa fotosintética [A] (μmol CO2 m-2 s-1), concentración de CO2 intracelular [Ci] (μmol CO2 mol-1), conductancia mesofílica [gm], rendimiento cuántico de PSII [phiPSII], velocidad de transporte de electrones [ETR] (μmol m-2 s-1), coeficiente de fluorescencia fotoquímica [qP], coeficiente de extinción de fluorescencia no fotoquímica [qN] y medida de disipación de calor no fotoquímico [NPQ].
Análisis estadísticos
Con los datos de las propiedades del suelo, los parámetros de germinación y atributos de calidad de las plantas de cada procedencia se realizó un ANOVA de rangos de KruskalWallis seguido de pruebas Z de comparación múltiple. Se realizó una matriz de correlación de Pearson (alfa = 0.05) y análisis de componentes principales para demostrar la relación entre las variables morfológicas y fisiológicas de calidad de las plantas y comprobar el efecto de las procedencias en la morfología de las semillas, la germinación y la calidad de plantas. Estos análisis se procesaron con el apoyo del software Origin Pro 2022.
Resultados
De acuerdo con el análisis de Kruskal-Wallis, existieron diferencias significativas en las propiedades del suelo de los árboles donde se seleccionaron semillas de O. pyramidale de las tres procedencias de estudio. El porcentaje de arena fue diferente entre procedencias (p = 0.03), con valores superiores en Napo y Pastaza (88.00% ± 2.31% y 77.67% ± 2.03%, respectivamente), lo cual se corresponde con la clase textural arenoso franco. El porcentaje de limo y arcilla también presentó diferencias significativas entre procedencias (limo, p = 0.03; arcilla, p = 0.03). Se encontraron valores superiores en la procedencia Orellana (28.00% ± 1.00% de limo y 21.00% ± 1.73% de arcilla), con una clase textural franco arenoso (Fig. 2).
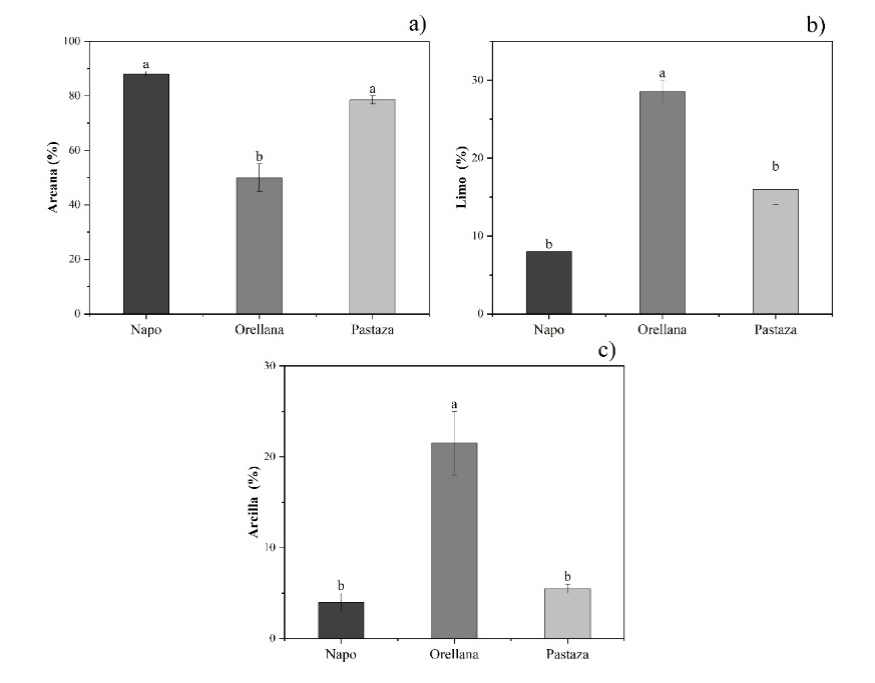
En las barras, letras desiguales indican diferencias significativas entre procedencias (p ≤ 0.05) mediante ANOVA de rangos de Kruskal-Wallis, seguidas de pruebas Z de comparación múltiple.
Figura 2 Porcentaje de arena, limo y arcilla del suelo de tres procedencias (Napo, Orellana y Pastaza) donde se seleccionaron semillas de árboles de O. pyramidale.
El pH resultó con variaciones entre procedencias (p = 0.03), con valores superiores en Orellana y Pastaza (5.65 ± 0.10 y 6.25 ± 0.03, respectivamente), valorado en la categoría de ligeramente ácido (Fig. 3a). El contenido de Al+3 H+ fue similar entre los tres orígenes geográficos (p = 0.17) y resultó con valores bajos (< 0.40 cmol/kg) (Fig. 3b). En cuanto al Al+3, no se presentaron variaciones (p = 0.06) y fue bajo en las tres procedencias (< 0.25 cmol/kg) (Fig. 3c). No hubo diferencias significativas en cuanto al contenido de N entre procedencias (p = 0.29) y los valores fueron bajos (< 0.3%) (Fig. 3d). La disponibilidad de P fue diferente entre procedencias (p = 0.04), con valores mayores en Pastaza (6.85 × 106 ± 0.04 × 106; 6.85 ppm ± 0.04 ppm) y fue calificado de bajo en todas las procedencias (Fig. 3e). El contenido de Mg2+ resultó con diferencias significativas entre las procedencias (p = 0.04), los valores mayores se obtuvieron en la procedencia de Orellana y Pastaza ([0.90 ± 0.06 y 0.75 ± 0.09] meq/100 ml, respectivamente) y fueron calificados de bajos (Fig. 3f). El contenido de M.O presentó diferencias significativas (p = 0.03) entre Orellana y Napo, teniendo esta última procedencia mayor porcentaje de M.O (18.6% ± 0.23%), casi el doble del valor porcentual registrado en Orellana, y en todos los casos fue alto (Fig. 3g). El contenido de Ca2+ y K+ fue diferente entre procedencias (p = 0.03), con valores superiores en Pastaza ([12.5 ± 0.21 y 0.42 ± 0.03] meq/100ml, respectivamente), valorado con alta disponibilidad (Fig. 3h,i).
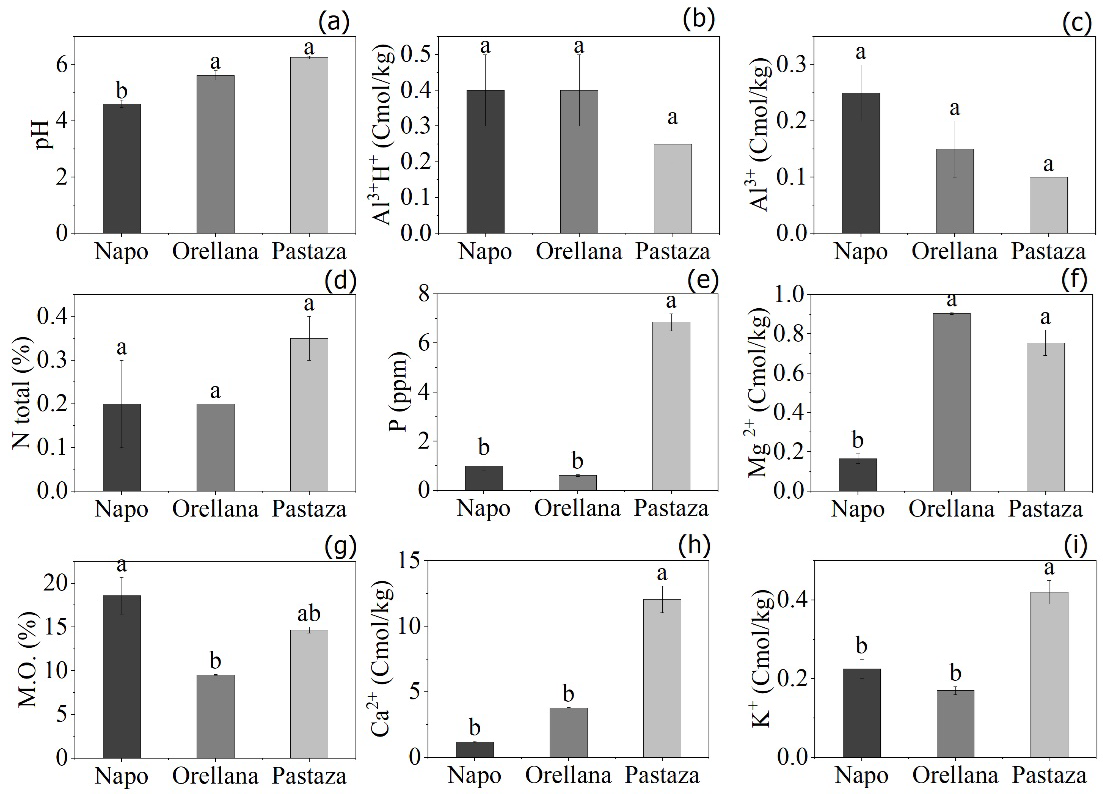
a) pH (potencial de hidrógeno); b) Al3+H+ (acidez intercambiable) c) Al3+ (aluminio); d) N total (nitrógeno total); e) P (fósforo); f) Mg2+ (magnesio); g) M.O (materia orgánica); h) Ca2+ (calcio); i) K+ (potasio). En las barras letras, desiguales indican diferencias significativas entre procedencias (p ≤ 0.05) mediante ANOVA de rangos de Kruskal-Wallis, seguidas de pruebas Z de comparación múltiple.
Figura 3 Propiedades químicas del suelo donde se seleccionaron semillas de tres procedencias (Napo, Orellana y Pastaza) de árboles de O. pyramidale.
El análisis de Kruskal-Wallis reflejó que no existieron diferencias significativas en el largo de las semillas (p = 0.33) entre las procedencias de estudio, mientras que el diámetro geométrico (p = 0.00), ancho (p = 0.02), espesor (p = 0.02), relación largo-ancho (p = 0.04) y área superficial fueron distintos (p = 0.01). Las semillas de las procedencias de Napo y Pastaza presentaron mayor ancho ([1.71 ± 0.35 y 1.60 ± 0.35] mm, respectivamente) y espesor ([1.75 ± 0.03 y 1.48 ± 0.04] mm, respectivamente) que las pertenecientes a Orellana (ancho de 1.40 mm ± 0.08 mm; espesor de 1.48 mm ± 0.04 mm). La relación largo/ancho fue superior en las semillas de la procedencia Orellana (2.23 ± 0.16), mientras que en Napo los valores fueron inferiores (1.80 ± 0.06). El área superficial fue superior en las semillas de la procedencia Napo (15.13 mm2 ± 0.79 mm2), mientras que en Orellana se encontró valores menores (10.05 mm2 ± 0.74 mm2) (Tabla 2).
Tabla 2 Parámetros morfológicos de las semillas de tres procedencias (Napo, Orellana y Pastaza) de la especie O. pyramidale.
| Parámetros morfológicos | Procedencia | ||
| Pastaza | Orellana | Napo | |
| L (mm) | 3.11a±0.11 | 3.03a±0.05 | 3.09a±0.06 |
| W (mm) | 1.60a±0.02 | 1.40b±0.08 | 1.71a±0.03 |
| T (mm) | 1.62a±0.07 | 1.48b±0.04 | 1.75a±0.03 |
| RLA | 1.94ab±0.08 | 2.23a±0.16 | 1.80b±0.06 |
| Dg (mm) | 2.73b±0.20 | 2.10c±0.15 | 3.10a±0.11 |
| S (mm2) | 13.60ab±1.36 | 10.05b±0.74 | 15.13a±0.79 |
L (largo); W (ancho); T (espesor de las semillas); RLA (relación largo-ancho); Dg (diámetro medio geométrico) y S (área superficial). En la tabla, letras desiguales indican diferencias significativas entre procedencias (p ≤ 0.05) mediante ANOVA de rangos de Kruskal-Wallis, seguidas de pruebas Z de comparación múltiple.
Conforme el análisis de Kruskal-Wallis, se comprobó que existieron diferencias significativas en los parámetros de germinación de las tres procedencias, a excepción del porcentaje de pureza como indicador de calidad de semillas (Tabla 3). La capacidad germinativa presentó valores superiores en las procedencias de Pastaza y Napo ([92.00 ± 0.57 y 89.00 ± 0.57]%), presentando 17.66% de semillas germinadas por encima de la procedencia de Orellana (p = 0.03). El tiempo medio de germinación indicó que la germinación ocurrió en menor tiempo en la procedencia de Napo y Pastaza, alrededor de los 7 días, sin embargo, en Orellana la germinación ocurrió en el doble del tiempo (alrededor de los 14 días) (p = 0.04). El índice de Timpson fue inferior en Pastaza y Napo ([0.60 ± 0.06 y 0.70 ± 0.06] %/días, respectivamente), mientras que en Orellana se encontró un valor porcentual de 1.60%/día ± 0.12%/día (p = 0.04), lo cual está relacionado con la velocidad de germinación. El valor útil presentó un porcentaje significativamente inferior en la procedencia de Orellana (69.66% ± 0.33%), con una diferencia del resto de alrededor de 20% (p = 0.03). El vigor germinativo presentó valores superiores en las procedencias de Pastaza y Napo (4.65% ± 0.27% y 4.16% ± 0.08%, respectivamente) (p = 0.04), como reflejo de la vigorosidad de las semillas.
Tabla 3 Parámetros germinativos de tres procedencias (Napo, Orellana y Pastaza) de la especie O. pyramidale
| Parámetros germinativos | Procedencia | ||
| Pastaza | Orellana | Napo | |
| P (%) | 98.33a±0.88 | 96.33a±0.08 | 97.33a±1.20 |
| CG (%) | 92.00a±0.57 | 71.66b±1.20 | 89.00a±0.57 |
| TMG (días) | 7.66b±0.38 | 14.00a±1.14 | 8.66b±0.38 |
| IT(%/días) | 0.60b±0.06 | 1.60a±0.12 | 0.70b±0.06 |
| VU (%) | 91.00a±0.57 | 69.66b±0.33 | 89.33a±0.33 |
| VG (%) | 4.65a±0.27 | 3.17b±0.07 | 4.16a±0.08 |
P (porcentaje de pureza); CG (capacidad germinativa); TMG (tiempo medio de germinación); IT (índice de Timpson); VU (valor útil); VG (vigor germinativo). En la tabla, letras desiguales indican diferencias significativas entre procedencias (p≤ 0.05) mediante ANOVA de rangos de Kruskal-Wallis, seguidas de pruebas Z de comparación múltiple.
La morfología de las plantas también resultó afectada por el origen geográfico, a excepción de: relación peso seco aéreo/peso seco radical, coeficiente de esbeltez y balance hídrico de las plantas, que fueron similares entre procedencias (Tabla 4). Se observaron valores inferiores en la morfología de las plantas para la procedencia de Orellana. Se evidenció una diferencia de 1.30 cm en la altura de las plantas (p = 0.00). El diámetro en el cuello de la raíz presentó una diferencia de 0.82 mm (p = 0.00). El área foliar resultó con una diferencia de alrededor de 8 cm2 (p = 0.02). El peso seco aéreo tuvo una diferencia de casi 0.20 g (p = 0.02). El peso seco radical fue 0.9 g menor (p = 0.01). El peso fresco total fue de 0.32 g inferior (p = 0.00). El volumen de raíz presentó una diferencia mínima de 0.36 cm3 (p = 0.02). El índice de calidad de Dickson resultó con 0.10 unidades diferentes al resto de las procedencias (p = 0.00). El índice de lignificación fue de 0.6 unidades inferior en la procedencia de Orellana (p = 0.02).
Tabla 4 Atributos e índices de calidad morfológica de plantas cultivadas en vivero de tres procedencias (Napo, Orellana, Pastaza) de O. pyramidale
| Atributos | Procedencia | ||
| Pastaza | Orellana | Napo | |
| H (cm) | 4.91a±0.19 | 3.53b±0.23 | 4.95a±0.26 |
| DCR (mm) | 3.35a±0.05 | 2.53b±0.04 | 3.38a±0.06 |
| AF (cm2) | 30.08a±0.92 | 22.15b±1.92 | 31.34a±2.70 |
| PSA (g) | 0.50a±0.01 | 0.31b±0.01 | 0.49a±0.02 |
| PSR (g) | 0.31a±0.02 | 0.21b±0.004 | 0.29a±0.002 |
| PST (g) | 0.81a±0.04 | 0.50b±0.04 | 0.78a±0.02 |
| PFT (g) | 4.61a±0.30 | 3.95b±0.12 | 4.27a±0.12 |
| VR (cm3) | 1.44a±0.02 | 1.02b±0.05 | 1.38a±0.03 |
| PA/PR | 1.61a±0.226 | 1.63a±0.147 | 1.68a±0.133 |
| EZ | 1.46a±0.065 | 1.40a±0.098 | 1.46a±0.086 |
| IQ | 0.26a±0.002 | 0.16b±0.002 | 0.25a±0.003 |
| IL | 0.18a±0.007 | 0.12b±0.004 | 0.18a±0.008 |
| BAP | 0.75a±0.015 | 0.73a±0.011 | 0.70a±0.019 |
H (altura de las plantas); DCR (diámetro en el cuello de la raíz); AF (área foliar); PSA (peso seco aéreo); PSR (peso seco radical); PST (peso seco total); PFT (peso fresco total); VR (volumen radical); PA/PR (relación parte aérea-radical); EZ (coeficiente de esbeltez); IQ (índice de calidad de Dickson); IL (índice de lignificación); BAP (balance hídrico). En la tabla, letras desiguales indican diferencias significativas entre procedencias (p≤ 0.05) mediante ANOVA de rangos de Kruskal-Wallis, seguidas de pruebas Z de comparación múltiple.
En cuanto a la respuesta fotosintética, las variables de mayor influencia fueron: tasa de asimilación, concentración de CO2 intracelular, rendimiento cuántico de PSII, velocidad de transporte de electrones y coeficiente de fluorescencia fotoquímica. En cambio, la tasa de transpiración, la conductancia estomática, la conductancia mesofílica, el coeficiente de fluorescencia no fotoquímica y la medida de disipación de calor no fotoquímico fueron similares entre los orígenes geográficos. La tasa de asimilación presentó una diferencia de casi 7 μmol CO2 m−2s−1, con valores superiores para las procedencias de Pastaza y Napo (p = 0.01). La concentración de CO2 intracelular tuvo una variación de alrededor de 69 μmol mol−1, con valores superiores para Orellana (p = 0.00). El rendimiento cuántico de PSII presentó una variación entre procedencias de 0.6 unidades, con medias superiores para Napo y Pastaza (p = 0.03). La velocidad de transporte de electrones presentó 23 μmol m−2 s−1 de variación entre procedencias, donde Orellana resultó con valores inferiores (p = 0.03). El coeficiente de fluorescencia fotoquímica tuvo una variación de 0.1 unidades entre procedencias, con valores inferiores para Orellana (p = 0.03) (Tabla 5).
Tabla 5 Atributos e índices de calidad fisiológica de plantas cultivadas en vivero de tres procedencias (Napo, Orellana, Pastaza) de O. pyramidale.
| Atributos | Procedencia | ||
| Pastaza | Orellana | Napo | |
| A (μmol CO2 m−2 s−1) | 13.37a±0.73 | 7.71b±0.12 | 12.14a±0.32 |
| E (mmol H2O m−2 s−1) | 0.93a±0.10 | 0.91a±0.15 | 0.98a±0.16 |
| Ci (μmol mol−1) | 399.90b±13.74 | 468.80a±8.45 | 421.10b±15.20 |
| gs (mmol H2O m−2 s−1) | 0.19a±0.02 | 0.13a±0.02 | 0.18a±0.04 |
| gm (mmol H2O m−2 s−1) | 0.01a±0.00 | 0.01a±0.00 | 0.01a±0.00 |
| phiPSII | 0.16a±0.01 | 0.10b±0.01 | 0.15ab±0.01 |
| ETR (μmol m−2 s−1) | 68.63a±7.10 | 45.05b±4.69 | 66.40ab±7.13 |
| qP | 0.28a±0.03 | 0.18b±0.01 | 0.27ab±0.02 |
| qN | 0.57a±0.02 | 0.60a±0.02 | 0.61a±0.02 |
| NPQ | 0.80a±0.06 | 0.93a±0.08 | 0.91a±0.06 |
A (tasa fotosintética); E (tasa de transpiración); Ci (concentración de CO2 intracelular); gs (conductancia estomática); gm (conductancia mesofílica); phiPSII (rendimiento cuántico de PSII); ETR (velocidad de transporte de electrones); qP (coeficiente de fluorescencia fotoquímica); qN (coeficiente de extinción de fluorescencia no fotoquímica); NPQ (medida de disipación de calor no fotoquímico). En la tabla, letras desiguales indican diferencias significativas entre procedencias (p ≤ 0.05) mediante ANOVA de rangos de Kruskal-Wallis, seguidas de pruebas Z de comparación múltiple.
La matriz de correlación de Pearson indicó que más de 80% de los atributos e índices morfológicos y fisiológicos presentaron una fuerte correlación (> 0.5). Se evidenció una correlación positiva alta entre las variables de morfología y fisiología de las plantas (> 0.7), lo que demuestra el grado de relación lineal existente entre estas variables. Sin embargo, hubo algunos parámetros fisiológicos (BAP, Ci, qN, NPQ) que presentaron una correlación negativa baja (< 0.3) (Fig. 4).
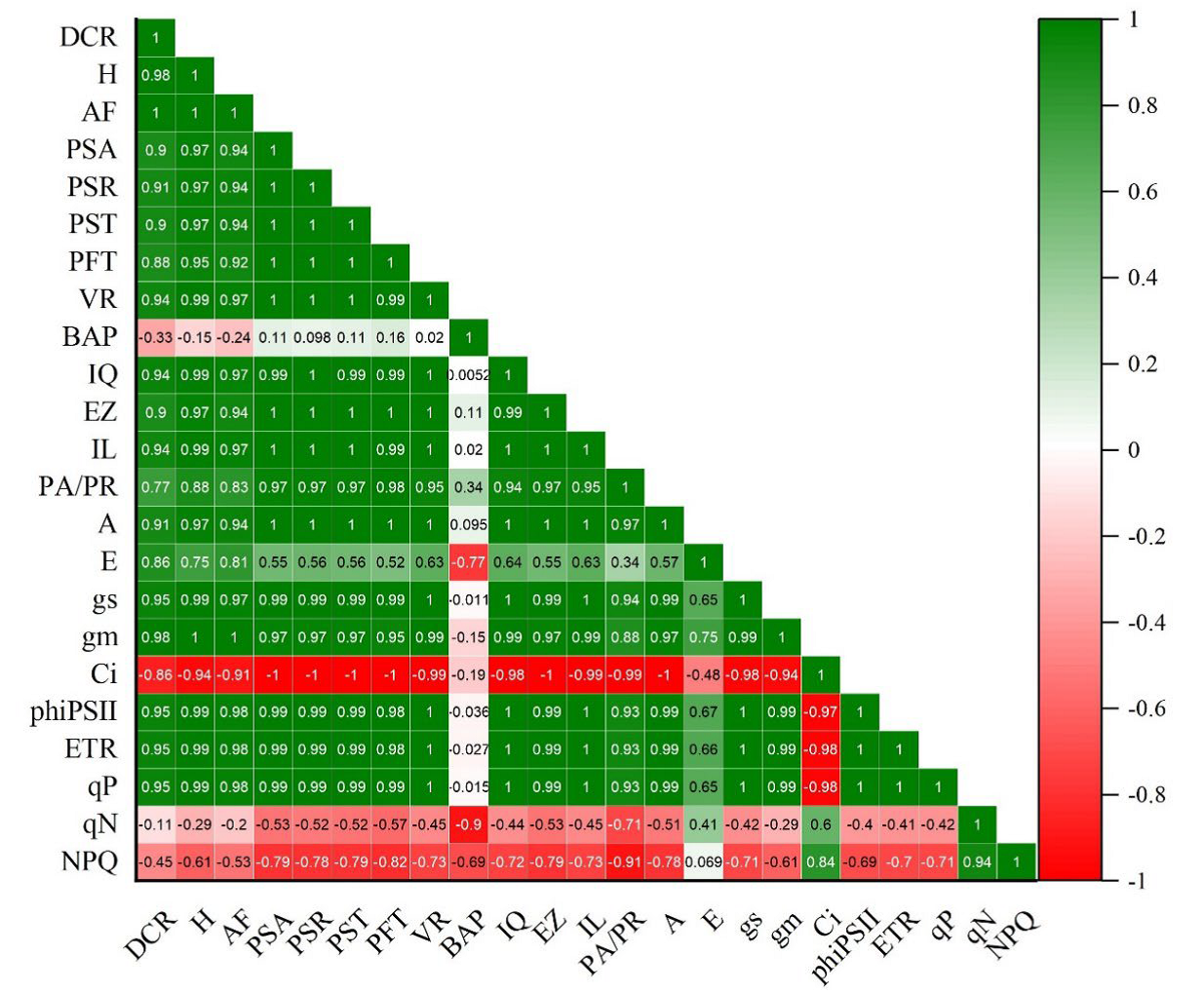
H (altura de las plantas); DCR (diámetro en el cuello de la raíz); AF (área foliar); PSA (peso seco aéreo); PSR (peso seco radical); PST (peso seco total); PFT (peso fresco total); VR (volumen radical); PA/PR (relación parte aérea-radical); EZ (coeficiente de esbeltez); IQ (índice de calidad de Dickson); IL (índice de lignificación); BAP(balance hídrico); A (tasa fotosintética); E (tasa de transpiración); Ci (concentración de CO2 intracelular); gs (conductancia estomática); gm (conductancia mesofílica); phiPSII (rendimiento cuántico de PSII); ETR (velocidad de transporte de electrones); qP (coeficiente de fluorescencia fotoquímica); qN (coeficiente de extinción de fluorescencia no fotoquímica); NPQ (medida de disipación de calor no fotoquímico).
Figura 4 Matriz de correlación de Pearson entre los parámetros e índices morfológicos y fisiológicos de calidad de las plantas de O. pyramidale cultivadas en vivero.
El análisis de componentes principales reflejó la presencia de dos componentes (PC1 y PC2). El PC1 explicó 85.7% de la varianza y el PC2 14.3% de la variabilidad total de los datos. Se encontró un patrón diferencial entre las procedencias de Pastaza y Napo con Orellana en cuanto a las variables morfológicas y fisiológicas. Las variables H, DCR, AF, PSA, PSR, PST, PFT, VR, PA/PR, EZ, IQ, IL, A, gs, gm, phiPSII, ETR y qP se encontraron influenciadas de forma positiva en el PC1, donde se ubicaron las procedencias de Pastaza y Napo, y las variables Ci, qN y NPQ tuvieron una influencia negativa en el PC2, donde se ubicó la procedencia de Orellana, mientras la variable BAP se asoció al PC2 en el extremo negativo (Fig. 5).
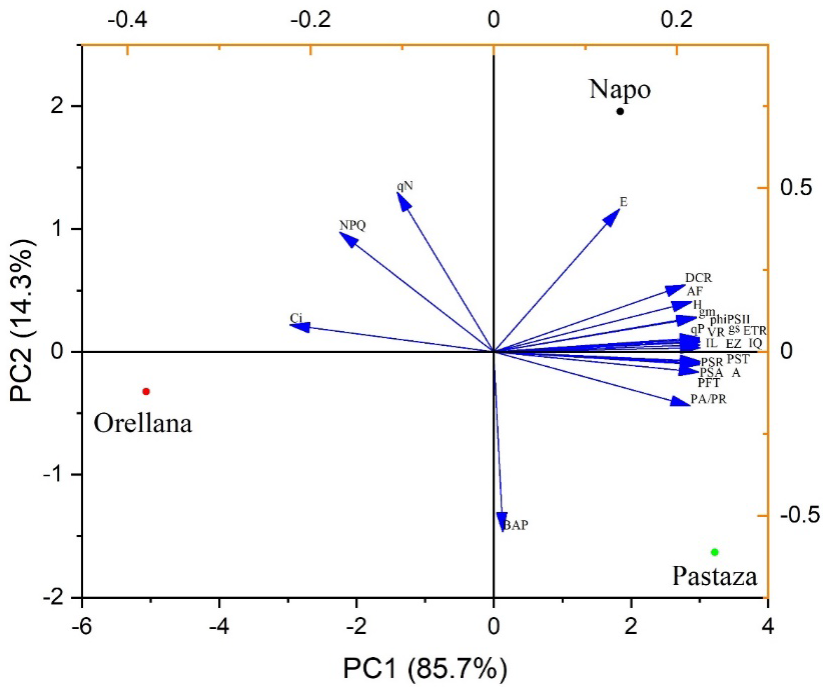
H (altura de las plantas); DCR (diámetro en el cuello de la raíz); AF (área foliar); PSA (peso seco aéreo); PSR (peso seco radical); PST (peso seco total); PFT (peso fresco total); VR (volumen radical); PA/PR (relación parte aérea-radical); EZ (coeficiente de esbeltez); IQ (índice de calidad de Dickson); IL (índice de lignificación); BAP(balance hídrico); A (tasa fotosintética); E (tasa de transpiración); Ci (concentración de CO2 intracelular); gs (conductancia estomática); gm (conductancia mesofílica); phiPSII (rendimiento cuántico de PSII); ETR (velocidad de transporte de electrones); qP (coeficiente de fluorescencia fotoquímica); qN (coeficiente de extinción de fluorescencia no fotoquímica); NPQ (medida de disipación de calor no fotoquímico).
Figura 5 Análisis de componentes principales de los atributos e índices morfológicos y fisiológicos de calidad de las plantas de O. pyramidale cultivadas en vivero. Eje x: PC1 (componente principal 1); Eje y: PC2 (componente principal 2).
La relación entre las procedencias (características ambientales) y las variables de morfología de las semillas, germinación y calidad de las plantas se visualiza en la Figura 6, con 100% de la variabilidad total explicada entre ambos componentes. Se identificó la existencia de dos grupos: por un lado, las procedencias Pastaza y Napo, ubicadas en el PC1 y, por otro Orellana, ubicada en el PC2. El mayor contenido de nutrientes, materia orgánica, pH, morfología de las semillas, germinación y calidad de las plantas se ubicó en las procedencias de Pastaza y Napo.
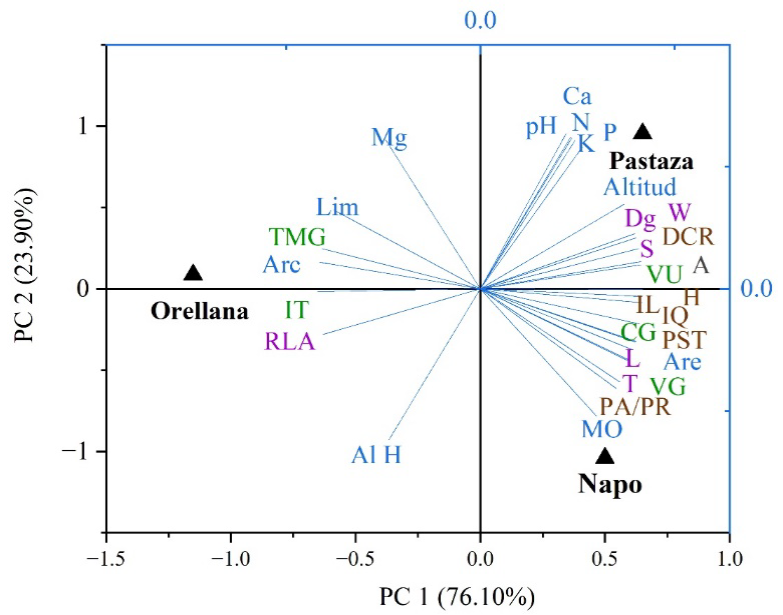
Variables ambientales (color azul): Lim (limo); Arc (arcilla); Are (arena); MO (materia orgánica); AL H( aluminio); pH (potencial de hidrógeno); Mg (magnesio); Ca (calcio); N (nitrógeno); K (potasio); P (fósforo); Variables de morfología de semillas (color morado): L (largo); W (ancho); T (espesor de las semillas); RLA (relación largo-ancho); Dg (diámetro medio geométrico) y S (área superficial); Variables de germinación (color verde): CG (capacidad germinativa); TMG (tiempo medio de germinación); IT (índice de Timpson); VU (valor útil); VG (vigor germinativo); Variables de calidad de planta (color marrón): H (altura de las plantas); DCR (diámetro en el cuello de la raíz); PST (peso seco total); PA/PR (relación parte aérea-radical); IQ (índice de calidad de Dickson); IL (índice de lignificación); A (tasa fotosintética).
Figura 6 Análisis de componentes principales de las características ambientales de las procedencias, morfología de las semillas, germinación y calidad de plantas de O. pyramidale cultivadas en vivero. Eje x: PC1 (componente principal 1); Eje y: PC2 (componente principal 2).
Discusión
La variación encontrada en las propiedades edáficas de los sitios de recolecta (clase textural, pH, M.O y nutrientes) indicaron que no son las mismas condiciones de suelo en las procedencias de estudio (Pastaza, Napo y Orellana). Resultados similares han sido obtenido por varios investígadores donde las propiedades físicas y químicas del suelo (estructura, pH y materia orgánica) cambian relativamente de un sitio a otro (Bravo et al., 2017; Martín y Pérez, 2009; Hernández et al., 2017). Las diferencias en el ambiente edáfico de los orígenes geográficos influyeron en la morfología de las semillas, germinación y calidad de las plantas de la especie O. pyramidale. La heterogeneidad encontrada en la biometría de las semillas (W, T, RLA, Dg y S), sugiere que la morfología de las semillas estuvo influenciada por las características del microambiente de cada procedencia. Las características de las semillas también pueden ser adaptables a las condiciones ambientales (Moles et al., 2004; Iglesias-Andreu y Tivo-Fernández, 2006). Esto coincide con los resultados del estudio donde las semillas de mayor tamaño correspondieron a las procedencias de Pastaza y Napo, las cuales se encontraron ubicadas a una mayor altitud (950 m y 577 m s.n.m., respectivamente), con mayor contenido de nutrientes, materia orgánica, suelos ligeramente ácidos y donde predomina una vegetación característica de bosque siempreverde piemontano de la Amazonía central. Esto pudo permitir un mejor desarrollo de las semillas, en contraste con la procedencia de Orellana, localizada a menor altitud (245 m s.n.m.), con menor contenido de nutrientes y predominio de vegetación de tierras bajas de la Amazonía baja. Las diferencias morfométricas que se presentan en las semillas de una especie se encuentran ligadas a las condiciones ambientales que prevalecen en el ciclo de vida de las mismas, de modo que la estrecha relación planta-ambiente hace énfasis en la importancia de poseer la suficiente flexibilidad en el proceso de desarrollo o de contar con mecanismos de resistencia para prevalecer en condiciones donde estas se encuentren ubicadas (Meyer et al., 1989; Ramírez-Morales y Orozco-Carmona, 2010; Santiago et al., 2017).
Las semillas de mayor tamaño (procedencias Pastaza y Napo) presentaron mayor porcentaje de germinación, valor útil y vigor germinativo, y germinaron en el menor tiempo. Estos resultados coinciden con la idea de que las semillas más grandes germinan más rápido y en mayor porcentaje que las semillas pequeñas (Harrison et al., 2014, Pozo-Gómez et al., 2019). En este sentido, previo a la producción de plantas en vivero y en función de la disponibilidad de germoplasma, se recomienda seleccionar las semillas de mayor tamaño con el propósito de tener mejores parámetros germinativos (Flores y González, 2021).
Varios estudios prueban que semillas de una misma especie, colectadas de distintos ambientes, difieren en viabilidad, germinación y crecimiento (Singh et al., 2006; Singh y Bhatt, 2010). En este estudio se ha comprobado la eficiencia en la respuesta a la germinación a partir de la selección de procedencias, señalando la conveniencia de seleccionar árboles semilleros de procedencias adecuadas para acelerar la germinación (Rodríguez-Vásquez et al., 2018). El comportamiento de los parámetros de germinación de la especie O. pyramidale concuerdan con lo observado por diversos autores (García et al., 2017; Toledo et al., 2019). La capacidad germinativa fue superior a lo mencionado por García et al. (2023) quien obtuvo 62.5%. Los mecanismos que regulan el inicio de la germinación están bajo presiones selectivas; de ahí que la variación de la capacidad de germinación entre y dentro de las especies se interpreta como una adaptación a las condiciones específicas del lugar y hábitat regional (Meyer et al., 1997).
Las variables morfológicas de las plantas cultivadas en vivero presentaron un patrón de comportamiento similar a la germinación de las semillas de O. pyramidale. La variación encontrada en la morfología (H, DCR, PF, PSA, PSR, PST y VR) pudo deberse a la procedencia, al sistema de producción y el manejo en vivero. Esto indicó una alta heterogeneidad de las plantas en vivero con características superiores en la procedencia de Pastaza y Napo.
La altura de la planta es un indicador del grado de desarrollo de la parte aérea, lo cual permite estimar el crecimiento de las plantas en campo (Gomes et al., 2002). Es un indicador poco útil por sí solo, aunque se asume que las plantas más altas pueden enfrentar mejor a la vegetación competidora (Birchler et al., 1998). En este estudio se comprobó que las plantas del origen geográfico Orellana tuvieron menor altura, por lo que es posible que tengan menor ventaja competitiva en las condiciones del sitio de plantación, aunque finalmente la calidad de la planta se determina en campo.
El diámetro en el cuello de la raíz se considera el indicador más confiable de desempeño en campo (Levy y McKay, 2003); también influye en la robustez, lo cual se asocia con el vigor y supervivencia (Tsakaldimi et al., 2013). Una planta con mayor DCR está mejor lignificada, tiene reservas de carbohidratos, presenta más yemas para rebrotes y la raíz está más desarrollada (Rodriguez et al., 2008). La calidad del DCR es uno de los parámetros más importantes para estimar la sobrevivencia después de haber realizado la plantación (Gomes et al., 2002). Conforme a lo que plantean Davide y Faria (2008), las plantas que presentan diámetro basal menor a 3 mm son fácilmente dañadas por hormigas y por lluvias torrenciales. Los resultados de esta investigación indicaron que las plantas de Orellana presentaron valores de DCR inferiores a 3 mm, por lo que pueden ser menos resistentes en las condiciones de plantación.
El peso seco total constituye un buen indicador de la capacidad de resistencia de las plantas en condiciones de campo (Gomes et al., 2003). Los valores de 0.81 y 0.78 g, alcanzados en las plantas de Pastaza y Napo, respectivamente, indicaron mayor resistencia para las condiciones de campo. Es conocido que la magnitud de la distribución de biomasa depende de factores ambientales y del genotipo (Olalde et al., 2000).
La proporción PA/PR refiere que una proporción mayor incrementa gradualmente las probabilidades de desequilibrio hídrico, y pone en peligro la supervivencia de la planta (Rueda et al., 2014). El análisis indicó que la procedencia no tiene un efecto significativo sobre la relación PA/PR, de ahí que un material plantable no podría ser más recomendable que otro frente a condiciones de restricción hídrica. Esto se corresponde con lo registrado en el índice BAP en el cual no se encontraron diferencias en cuanto a la economía hídrica de las plantas. Esto tiene implicaciones prácticas importantes para el establecimiento de la plantación, ya que las plantas de las tres procedencias presentaron homogeneidad en la eficiencia en el uso del agua. El coeficiente de esbeltez estima la resistencia mecánica de las plantas y se recomienda plantas más esbeltas para garantizar mayor capacidad para tolerar condiciones adversas (Rodriguez et al., 2008). Con base en los resultados obtenidos, no pueden señalarse procedencias más idóneas frente a condiciones hostiles, por lo que, de acuerdo con las medias estadísticas todas las plantas son similares.
El índice de calidad de Dickson presenta un carácter integrador, por cuanto expresa la combinación de varios parámetros morfológicos, estableciendo la proporción en que se encuentra la planta en lo que respecta a tamaño y peso seco que esta posee (Gonzalez et al., 1996). Los resultados del presente trabajo indicaron que las plántulas de O. pyramidale de las procedencias de Pastaza y Napo presentaron valores superiores (> 0.25) por lo que se pueden considerar de mejor calidad y con mayor equilibrio entre las fracciones de la parte aérea y radical. Lo deseable es que la planta alcance los máximos valores de IQ, lo cual implica que el desarrollo de la planta sea grande y que, al mismo tiempo, las fracciones aérea y radical estén equilibradas (Dickson et al., 1960). El valor mínimo del IQ para que una planta sea considerada de calidad debe ser de 0.20 (Silva et al., 2013). En este trabajo el índice IQ, en las tres procedencias, se encontró entre 0.16 y 0.26 y solo la procedencia Orellana resultó con un valor inferior a lo registrado en la literatura, por lo que se puede decir que las plantas de la procedencia Orellana no son de buena calidad.
El índice de lignificación es fundamental para evaluar la planta antes de que salga a campo. Las plantas de los orígenes geográficos Pastaza y Napo tuvieron los valores mayores (0.18), lo cual indicó mayor formación de lignina (Reyes et al., 2018).
Se ha observado que altas tasas de fotosíntesis derivan en una ganancia mayor de carbono que propicia la producción de plantas grandes, las cuales tienen mayor oportunidad de sobrevivir en campo (Luis et al., 2009; Villar et al., 2012).
La variación encontrada entre las procedencias en cuanto a la morfología y fisiología de las plantas puede atribuirse a diferencias genéticas causadas por la adaptación a diversas condiciones ambientales (Fontana et al., 2015). Se ha comprobado que presiones de selección diferentes dan lugar a distintas adaptaciones (Climent et al., 2002) que determinan la capacidad de un germoplasma para establecerse en un ambiente. Esto permite fundamentar la necesidad de estudiar y establecer patrones referenciales para los parámetros de calidad morfológica y fisiológica de especies nativas como O. pyramidale con alto potencial para la reforestación en sitios degradados.
Es notorio que los trabajos en vivero están limitados en la determinación de las respuestas fisiológicas. En este estudio, se demostró que, bajo las condiciones experimentales de vivero, existe una variación en los parámetros fisiológicos de las plantas en función del sitio de origen, lo que sugiere que las condiciones edafoclimáticas de los sitios de recolección influyen en los procesos metabólicos, la eficiencia fotosintética y los procesos de translocación de los asimilados desde la etapa temprana. La respuesta funcional de las plantas está relacionada con la mayor asignación de biomasa (Cordeiro et al., 2021), esto podría propiciar para las plantas de la procedencia de Napo y Pastaza un mejor desempeño en condiciones de campo y mayor potencial para uso en reforestaciones y plantaciones forestales comerciales.
La fotosíntesis es uno de los procesos que afecta la asimilación y uso de los recursos tales como carbono, agua y nutrientes. Estos factores influyen directamente en el crecimiento de la planta y son aspectos importantes en la selección del material genético (Cordeiro et al., 2021). En el estudio se obtuvieron resultados similares a los mencionados por Marenco et al. (2001) con una tasa de asimilación de CO2 de 13 μmol m-2s-1, y Arteaga-Crespo et al. (2022) con valores de hasta 15 μmol m-2s-1.
Los menores valores de transpiración observados en la procedencia Orellana sugieren menor consumo de agua (Okuto y Ouma, 2010). La estimación de estos parámetros es clave para predecir la productividad de las plantaciones y la eficiencia del uso del agua en ecosistemas de producción (Yu et al., 2004).
Conclusiones
Se comprobó la influencia del origen geográfico en la morfología de las semillas, parámetros de germinación y características morfofuncionales de plántulas de Ochroma pyramidale cultivadas en vivero, lo cual está regulado por las condiciones ambientales del sitio. Esto determinó la existencia de dos grupos diferentes: por un lado, la procedencia de Pastaza y Napo, con un comportamiento superior y, por otro, Orellana. Estos resultados tienen implicaciones prácticas importantes para definir estrategias de recolección y manejo de las semillas, así como para el manejo de las plantas en vivero para su uso en programas de forestación y/o repoblación forestal en condiciones de la Amazonía ecuatoriana.

